
Optimization of Porphyran Extraction from Pyropia yezoensis by Response Surface Methodology and Its Lipid-Lowering Effects

Abstract
:1. Introduction
2. Results
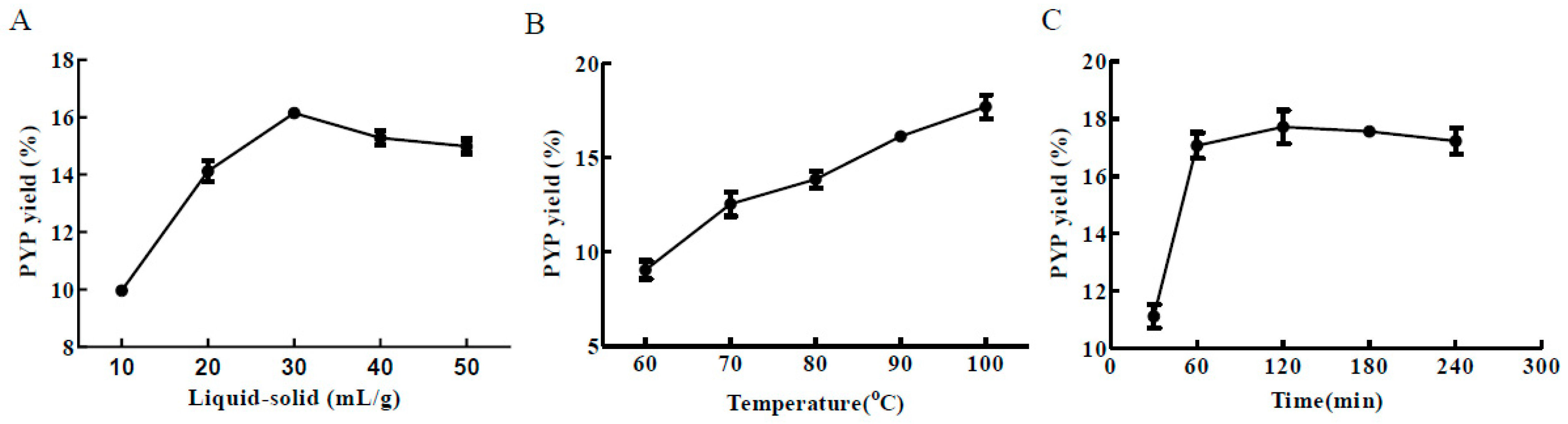
2.1. Single-Factor Experiment Analysis
2.2. Response Surface Analysis
2.3. Purification of PYP and Its Physicochemical Properties
2.4. FT-IR Spectroscopy Characteristics of PPYP
2.5. Effect of PPYP on Lipid Accumulation in Palmitic Acid (PA)-Induced HepG2 Cells
2.5.1. Cytotoxicity of PPYP and PA on HepG2 Cells
2.5.2. PPYP Alleviates PA-Induced TG Accumulation in HepG2 Cells
2.6. Effect of PPYP on Lipid Accumulation in High-Sucrose-Fed D. melanogaster Larvae
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Single-Factor Design for Pyropia Yezoensis Polysaccharide
4.3. Experimental Design
4.4. Purification of PYP
4.5. Characteristics of PPYP Analysis
4.6. Protocol for Obtaining the Palmitic Acid (PA)-Induced HepG2 Cells
4.6.1. Cell Culture and Treatment
4.6.2. Triglyceride Content Analysis
4.6.3. Oil Red O Staining
4.7. Experimental Protocol for Obtaining High-Sucrose-Fed D. melanogaster Larvae
4.7.1. D. melanogaster Larvae and Treatment
4.7.2. Triglyceride Content Analysis
4.8. qPCR Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rahman, M.A.; Halfar, J. First evidence of chitin in calcified coralline algae: New insights into the calcification process of Clathromorphum compactum. Sci. Rep. 2014, 4, 6162. [Google Scholar] [CrossRef]
- Cunha, L.; Grenha, A. Sulfated seaweed polysaccharides as multifunctional materials in drug delivery applications. Mar. Drugs 2016, 14, 42. [Google Scholar] [CrossRef] [PubMed]
- Ścieszka, S.; Klewicka, E. Algae in food: A general review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3538–3547. [Google Scholar] [CrossRef] [PubMed]
- Manlusoc, J.K.T.; Hsieh, C.-L.; Hsieh, C.-Y.; Salac, E.S.N.; Lee, Y.-T.; Tsai, P.-W. Pharmacologic application potentials of sulfated polysaccharide from marine algae. Polymers 2019, 11, 1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-E.; Kim, H.; Seong-Chul, H.; Park, T.; Bin Lee, K.; Yoo, S.-Y.; Hong, S.-C.; Kim, J.T.; Lee, J. Marine polysaccharides: Therapeutic efficacy and biomedical applications. Arch. Pharmacal Res. 2017, 40, 1006–1020. [Google Scholar] [CrossRef] [PubMed]
- Cena, H.; Calder, P.C. Defining a healthy diet: Evidence for the role of contemporary dietary patterns in health and disease. Nutrients 2020, 12, 334. [Google Scholar] [CrossRef] [Green Version]
- Ferramosca, A.; Zara, V. Modulation of hepatic steatosis by dietary fatty acids. World J. Gastroenterol. 2014, 20, 1746–1755. [Google Scholar] [CrossRef]
- Alves-Bezerra, M.; Cohen, D.E. Triglyceride metabolism in the liver. Compr. Physiol. 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Ngamlerst, C.; Udomkasemsab, A.; Kongkachuichai, R.; Kwanbunjan, K.; Chupeerach, C.; Prangthip, P. The potential of an-tioxidant-rich Maoberry (Antidesma bunius) extract on fat metabolism in liver tissues of rats fed a high-fat diet. BMC Complement. Altern. Med. 2019, 19, 294. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease—Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Bedogni, G.; Miglioli, L.; Masutti, F.; Tiribelli, C.; Marchesini, G.; Bellentani, S. Prevalence of and risk factors for nonalcoholic fatty liver disease: The Dionysos nutrition and liver study. Hepatology 2005, 42, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Koehler, E.M.; Schouten, J.; Hansen, B.E.; van Rooij, F.J.A.; Hofman, A.; Stricker, B.H.C.; Janssen, H.L.A. Prevalence and risk factors of non-alcoholic fatty liver disease in the elderly: Results from the Rotterdam study. J. Hepatol. 2012, 57, 1305–1311. [Google Scholar] [CrossRef] [PubMed]
- Perlemuter, G.; Bigorgne, A.; Cassard, A.-M.; Naveau, S. Nonalcoholic fatty liver disease: From pathogenesis to patient care. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Tessari, P.; Coracina, A.; Cosma, A.; Tiengo, A. Hepatic lipid metabolism and non-alcoholic fatty liver disease. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 291–302. [Google Scholar] [CrossRef]
- Lim, J.S.; Mietus-Snyder, M.; Valente, A.; Schwarz, J.-M.; Lustig, R.H. The role of fructose in the pathogenesis of NAFLD and the metabolic syndrome. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 251–264. [Google Scholar] [CrossRef]
- Cohen, J.C.; Horton, J.D.; Hobbs, H.H. Human fatty liver disease: Old questions and new insights. Science 2011, 332, 1519–1523. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.; Xu, Y.; Xie, J.; Sun, C.; Zheng, X.; Chen, W. Systematic evaluation of phenolic compounds and protective capacity of a new mulberry cultivar J33 against palmitic acid-induced lipotoxicity using a simulated digestion method. Food Chem. 2018, 258, 43–50. [Google Scholar] [CrossRef]
- Ogino, N.; Miyagawa, K.; Kusanaga, M.; Hayashi, T.; Minami, S.; Oe, S.; Honma, Y.; Harada, M. Involvement of sarco/endoplasmic reticulum calcium ATPase-mediated calcium flux in the protective effect of oleic acid against lipotoxicity in hepatocytes. Exp. Cell Res. 2019, 385, 111651. [Google Scholar] [CrossRef]
- Malik, V.S.; Popkin, B.M.; Bray, G.A.; Després, J.-P.; Hu, F.B. Sugar-sweetened beverages, obesity, type 2 diabetes mellitus, and cardiovascular disease risk. Circulation 2010, 121, 1356–1364. [Google Scholar] [CrossRef]
- Nikpartow, N.; Danyliw, A.D.; Whiting, S.J.; Lim, H.; Vatanparast, H. Fruit drink consumption is associated with overweight and obesity in Canadian women. Can. J. Public Health 2012, 103, 178–182. [Google Scholar] [CrossRef]
- Kok, N.; Roberfroid, M.; Delzenne, N. Dietary oligofructose modifies the impact of fructose on hepatic triacylglycerol metabolism. Metabolism 1996, 45, 1547–1550. [Google Scholar] [CrossRef]
- Musselman, L.P.; Fink, J.L.; Ramachandran, P.V.; Patterson, B.W.; Okunade, A.L.; Maier, E.; Brent, M.R.; Turk, J.; Baranski, T.J. Role of fat body lipogenesis in protection against the effects of caloric overload in Drosophila. J. Biol. Chem. 2013, 288, 8028–8042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Li, K.; Shao, J.; Luo, Q.; Jin, L.H. Flos Chrysanthemi Indici extract improves a high-sucrose diet-induced metabolic disorder in Drosophila. Exp. Ther. Med. 2018, 16, 2564–2572. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.S.; Qi, W.; Farias-Pereira, R.; Choi, S.; Clark, J.M.; Kim, D.; Park, Y. Permethrin and ivermectin modulate lipid metabolism in steatosis-induced HepG2 hepatocyte. Food Chem. Toxicol. 2019, 125, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Chen, X.; Xu, X.; Liu, J.; Fu, C.; Zhao, D.; Zhao, W.; Ma, R.; Sun, L. Mechanism underlying p-coumaric acid alleviation of lipid accumulation in palmitic acid-treated human hepatoma cells. J. Agric. Food Chem. 2020, 68, 3742–3749. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhao, M.-G.; Jiang, C.-H.; Sheng, X.-P.; Yang, H.-M.; Liu, Y.; Yao, X.-M.; Zhang, J.; Yin, Z.-Q. Triterpenic acids-enriched fraction from Cyclocarya paliurus attenuates insulin resistance and hepatic steatosis via PI3K/Akt/GSK3β pathway. Phytomedicine 2020, 66, 153130. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.; Lee, W.H.; Jeong, J.; Park, M.; Ko, J.Y.; Kwon, O.W.; Lee, J.; Kim, Y.J. Pyropia yezoensis extract suppresses IFN-gamma-and TNF-alpha-induced proinflammatory chemokine production in HaCaT cells via the down-regulation of NF-κB. Nutrients 2020, 12, 1238. [Google Scholar] [CrossRef]
- He, D.; Wu, S.; Yan, L.; Zuo, J.; Cheng, Y.; Wang, H.; Liu, J.; Zhang, X.; Wu, M.; Choi, J.; et al. Antitumor bioactivity of porphyran extracted from Pyropia yezoensis Chonsoo2 on human cancer cell lines. J. Sci. Food Agric. 2019, 99, 6722–6730. [Google Scholar] [CrossRef]
- Geng, L.; Wang, J.; Zhang, Z.; Yue, Y.; Zhang, Q. Structure and bioactivities of porphyrans and oligoporphyrans. Curr. Pharm. Des. 2019, 25, 1163–1171. [Google Scholar] [CrossRef]
- Jin, W.; Zhang, W.; Liu, G.; Yao, J.; Shan, T.; Sun, C.; Zhang, Q. The structure-activity relationship between polysaccharides from Sargassum thunbergii and anti-tumor activity. Int. J. Biol. Macromol. 2017, 105, 686–692. [Google Scholar] [CrossRef]
- Liu, C.; Omer, A.M.; Ouyang, X.K. Adsorptive removal of cationic methylene blue dye using carboxymethyl cellulose/k-carrageenan/activated montmorillonite composite beads: Isotherm and kinetic studies. Int. J. Biol. Macromol. 2006, 106, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Sekkal, M.; Legrand, P. A spectroscopic investigation of the carrageenans and agar in the 1500–100 cm−1 spectral range. Spectrochim. Acta Part A Mol. Spectrosc. 1993, 49, 209–221. [Google Scholar] [CrossRef]
- Zhao, T.; Zhang, Q.; Qi, H.; Zhang, H.; Niu, X.; Xu, Z.; Li, Z. Degradation of porphyran from Porphyra haitanensis and the antioxidant activities of the degraded porphyrans with different molecular weight. Int. J. Biol. Macromol. 2006, 38, 45–50. [Google Scholar] [CrossRef]
- Wang, P.; Zhao, X.; Lv, Y.; Li, M.; Liu, X.; Li, G.; Yu, G. Structural and compositional characteristics of hybrid carrageenans from red algae Chondracanthus chamissoi. Carbohydr. Polym. 2012, 89, 914–919. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhu, Z.; Tang, Y.; Ren, Y.; Song, Q.; Tang, Y.; Zhang, Y. Structural characterization and antitumor activity of a novel Se-polysaccharide from selenium-enriched Cordyceps gunnii. Food Funct. 2018, 9, 2744–2754. [Google Scholar] [CrossRef]
- Barker, S.A.; Bourne, E.J.; Stacey, M.; Whiffen, D.H. Infra-red spectra of carbohydrates: Part I: Some derivatives of D-glucopyranose. J. Chem. Soc. 1954, 1, 171–176. [Google Scholar] [CrossRef]
- Yang, X.; Huang, M.; Qin, C.; Lv, B.; Mao, Q.; Liu, Z. Structural characterization and evaluation of the antioxidant activities of polysaccharides extracted from Qingzhuan brick tea. Int. J. Biol. Macromol. 2017, 101, 768–775. [Google Scholar] [CrossRef]
- Ran, X.L.; Zhang, M.; Wang, Y.; Adhikari, B. Novel technologies applied for recovery and value addition of high value compounds from plant byproducts: A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 450–461. [Google Scholar] [CrossRef]
- García-Vaquero, M.; Rajauria, G.; O’Doherty, J.V.; Sweeney, T. Polysaccharides from macroalgae: Recent advances, innovative technologies and challenges in extraction and purification. Food Res. Int. 2017, 99, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.-H.; Chiu, C.-Y.; Lu, T.-J.; Liu, S.-H.; Chiang, M.-T. The anti-obesity effect of polysaccharide-rich red algae (Gelidium amansii) hot-water extracts in high-fat diet-induced obese hamsters. Mar. Drugs 2019, 17, 532. [Google Scholar] [CrossRef] [Green Version]
- El Rashed, Z.; Grasselli, E.; Khalifeh, H.; Canesi, L.; Demori, I. Brown-algae polysaccharides as active constituents against nonalcoholic fatty liver disease. Planta Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Chater, P.I.; Wilcox, M.D.; Houghton, D.; Pearson, J.P. The role of seaweed bioactives in the control of digestion: Implications for obesity treatments. Food Funct. 2015, 6, 3420–3427. [Google Scholar] [CrossRef] [PubMed]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donato, M.; Tolosa, L.; Gómez-Lechón, M.J. Culture and functional characterization of human hepatoma HepG2 cells. In Protocols in In Vitro Hepatocyte Research; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2014; Volume 1250, pp. 77–93. [Google Scholar]
- Tu, Z.; Moss-Pierce, T.; Ford, P.; Jiang, T.A. Rosemary (Rosmarinus officinalis L.) extract regulates glucose and lipid metabolism by activating AMPK and PPAR pathways in HepG2 cells. J. Agric. Food Chem. 2013, 61, 2803–2810. [Google Scholar] [CrossRef] [PubMed]
- Zhong, D.; Xie, Z.; Huang, B.; Zhu, S.; Wang, G.; Zhou, H.; Lin, S.; Lin, Z.; Yang, B. Ganoderma lucidum polysaccharide peptide alleviates hepatoteatosis via modulating bile acid metabolism dependent on FXR-SHP/FGF. Cell. Physiol. Biochem. 2018, 49, 1204–1220. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lang, H.; Chen, K.; Zhang, Y.; Gao, Y.; Ran, L.; Yi, L.; Mi, M.; Zhang, Q. Resveratrol protects against nonalcoholic fatty liver disease by improving lipid metabolism and redox homeostasis via the PPARα pathway. Appl. Physiol. Nutr. Metab. 2020, 45, 227–239. [Google Scholar] [CrossRef]
- Zhang, Q.; Huang, Y.; Li, X.; Liu, H.; He, B.; Wang, B.; Ma, Y.; Zhou, X.; Liu, Y.; Wu, S. Tangduqing granules attenuate insulin resistance and abnormal lipid metabolism through the coordinated regulation of PPARγ and DGAT2 in type 2 diabetic rats. J. Diabetes Res. 2019, 2019, 7403978. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.; Pan, Y.; Zhang, Z.; He, Y.; Teng, Y.; Liang, H.; Wu, X.; Yang, H.; Zhou, P. Amelioration of the lipogenesis, oxidative stress and apoptosis of hepatocytes by a novel proteoglycan from Ganoderma lucidum. Biol. Pharm. Bull. 2020, 43, 1542–1550. [Google Scholar] [CrossRef]
- Kim, M.K.; Kim, S.H.; Yu, H.S.; Park, H.G.; Kang, U.G.; Ahn, Y.M.; Kim, Y.S. The effect of clozapine on the AMPK-ACC-CPT1 pathway in the rat frontal cortex. Int. J. Neuropsychopharmacol. 2011, 15, 907–917. [Google Scholar] [CrossRef] [Green Version]
- Tailleux, A.; Wouters, K.; Staels, B. Roles of PPARs in NAFLD: Potential therapeutic targets. Biochim. Biophys. Acta 2012, 1821, 809–818. [Google Scholar] [CrossRef]
- Kim, M.H.; Seong, J.B.; Huh, J.-W.; Bae, Y.C.; Lee, H.-S.; Lee, D.-S. Peroxiredoxin 5 ameliorates obesity-induced non-alcoholic fatty liver disease through the regulation of oxidative stress and AMP-activated protein kinase signaling. Redox Biol. 2020, 28, 101315. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.; Abdelmalek, M.F.; Sullivan, S.; Nadeau, K.J.; Green, M.; Roncal, C.; Nakagawa, T.; Kuwabara, M.; Sato, Y.; Kang, D.-H.; et al. Fructose and sugar: A major mediator of non-alcoholic fatty liver disease. J. Hepatol. 2018, 68, 1063–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rovenko, B.M.; Kubrak, O.I.; Gospodaryov, D.V.; Perkhulyn, N.V.; Yurkevych, I.S.; Sanz, A.; Lushchak, O.; Lushchak, V.I. High sucrose consumption promotes obesity whereas its low consumption induces oxidative stress in Drosophila melanogaster. J. Insect Physiol. 2015, 79, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Koyama, T.; Texada, M.J.; Halberg, K.A.; Rewitz, K. Metabolism and growth adaptation to environmental conditions in Drosophila. Cell. Mol. Life Sci. 2020, 77, 4523–4551. [Google Scholar] [CrossRef] [PubMed]
- Ugur, B.; Chen, K.; Bellen, H.J. Drosophila tools and assays for the study of human diseases. Dis. Model Mech. 2016, 9, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Ugrankar, R.; Liu, Y.; Provaznik, J.; Schmitt, S.; Lehmann, M. Lipin is a central regulator of adipose tissue development and function in Drosophila melanogaster. Mol. Cell. Biol. 2011, 31, 1646–1656. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Lee, S.K.; Paik, D.; Min, K.J. Overexpression of fatty-acid-β-oxidation-related genes extends the lifespan of Drosophila melanogaster. Oxid. Med. Cell. Longev. 2012, 2012, 854502. [Google Scholar] [CrossRef] [Green Version]
- Qian, Z.-G. Cellulase-assisted extraction of polysaccharides from Cucurbita moschata and their antibacterial activity. Carbohydr. Polym. 2014, 101, 432–434. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, X.; Liu, J.; Song, J.; Yu, P.; Chen, P.; Liao, Z.; Wu, M.; Tong, H. Physicochemical characterization of Sargassum fusiforme fucoidan fractions and their antagonistic effect against P-selectin-mediated cell adhesion. Int. J. Biol. Macromol. 2019, 133, 656–662. [Google Scholar] [CrossRef]
- Dodgson, K.S.; Price, R.G. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | A: Liquid–Solid Ratio (mL/g) | B: Time (min) | C: Temperature (°C) | PYP Yield (%) | |
|---|---|---|---|---|---|
| Actual Value | Predicted Value | ||||
| 1 | 20 (0) | 30 (−1) | 100 (1) | 18.26 | 18.40 |
| 2 | 10 (−1) | 120 (1) | 90 (0) | 9.96 | 10.07 |
| 3 | 20 (0) | 75 (0) | 90 (0) | 14.10 | 14.34 |
| 4 | 20 (0) | 75 (0) | 90 (0) | 14.27 | 14.34 |
| 5 | 20 (0) | 120 (1) | 80 (−1) | 14.12 | 13.98 |
| 6 | 20 (0) | 30 (−1) | 80 (−1) | 11.57 | 11.48 |
| 7 | 20 (0) | 75 (0) | 90 (0) | 14.48 | 14.34 |
| 8 | 10 (−1) | 30 (−1) | 90 (0) | 10.67 | 10.73 |
| 9 | 20 (0) | 75 (0) | 90 (0) | 14.78 | 14.34 |
| 10 | 30 (1) | 75 (0) | 100 (1) | 20.28 | 20.25 |
| 11 | 10 (−1) | 75 (0) | 80 (−1) | 10.46 | 10.49 |
| 12 | 10 (−1) | 75 (0) | 100 (1) | 17.63 | 17.43 |
| 13 | 30 (1) | 30 (−1) | 90 (0) | 10.74 | 10.63 |
| 14 | 20 (0) | 120 (1) | 100 (1) | 20.60 | 20.69 |
| 15 | 20 (0) | 75 (0) | 90 (0) | 14.05 | 14.34 |
| 16 | 30 (1) | 120 (1) | 90 (0) | 16.15 | 16.09 |
| 17 | 30 (1) | 75 (0) | 80 (−1) | 13.37 | 13.57 |
| Source | Sum of Squares | Df | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 172.93 | 9 | 19.21 | 254.25 | *** |
| A-Liquid–solid | 17.46 | 1 | 17.46 | 231.10 | *** |
| B-Time | 11.50 | 1 | 11.50 | 152.12 | *** |
| C-Temperature | 92.82 | 1 | 92.82 | 1228.26 | *** |
| AB | 9.36 | 1 | 9.36 | 123.91 | *** |
| AC | 0.017 | 1 | 0.017 | 0.22 | 0.6507 |
| BC | 0.011 | 1 | 0.011 | 0.15 | 0.7138 |
| A2 | 10.50 | 1 | 10.50 | 138.96 | *** |
| B2 | 3.24 | 1 | 3.24 | 42.83 | *** |
| C2 | 30.20 | 1 | 30.20 | 399.65 | *** |
| Residual | 0.53 | 7 | 0.076 | ||
| Lack of fit | 0.17 | 3 | 0.056 | 0.63 | 0.6344 |
| Pure error | 0.36 | 4 | 0.090 | ||
| Correlation total | 173.46 | 16 | |||
| R2 | 0.9970 | R2adj | 0.9930 | ||
| C.V.% | 1.90 | Pred R-Squared | 0.9811 |
| Name | Sugar (%) | Sulfate (mmol/mL) | 3,6-anhydro-α-ʟ-galactose (%) | Molecular Weight (kDa) | Monosaccharide Molar Ratio Gal Rha | |
|---|---|---|---|---|---|---|
| PPYP | 93.2 ± 1.5 | 1.2 ± 0.03 | 20.8 ± 1.1 | 151.6 | 5.3 | 1.0 |
| Symbols | Independent Variable | Level | ||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| A | Liquid–solid ratio (mL/g) | 10:1 | 20:1 | 30:1 |
| B | Time (min) | 30 | 75 | 120 |
| C | Temperature (°C) | 80 | 90 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, D.; Yan, L.; Hu, Y.; Wu, Q.; Wu, M.; Choi, J.-i.; Tong, H. Optimization of Porphyran Extraction from Pyropia yezoensis by Response Surface Methodology and Its Lipid-Lowering Effects. Mar. Drugs 2021, 19, 53. https://doi.org/10.3390/md19020053
He D, Yan L, Hu Y, Wu Q, Wu M, Choi J-i, Tong H. Optimization of Porphyran Extraction from Pyropia yezoensis by Response Surface Methodology and Its Lipid-Lowering Effects. Marine Drugs. 2021; 19(2):53. https://doi.org/10.3390/md19020053
Chicago/Turabian StyleHe, Dan, Liping Yan, Yingxia Hu, Qifang Wu, Mingjiang Wu, Jong-il Choi, and Haibin Tong. 2021. "Optimization of Porphyran Extraction from Pyropia yezoensis by Response Surface Methodology and Its Lipid-Lowering Effects" Marine Drugs 19, no. 2: 53. https://doi.org/10.3390/md19020053
APA StyleHe, D., Yan, L., Hu, Y., Wu, Q., Wu, M., Choi, J. -i., & Tong, H. (2021). Optimization of Porphyran Extraction from Pyropia yezoensis by Response Surface Methodology and Its Lipid-Lowering Effects. Marine Drugs, 19(2), 53. https://doi.org/10.3390/md19020053