2. Results and Discussion
The exact masses of the compounds (MS
1 data,
Figure S1) were compared with compounds in the MarinLit database (
Table 1). The results showed that exact masses of compounds
1–
3 closely matched with streptodepsipeptide P11B ([M] = 1082.5988), P11A ([M] = 1096.6142), and valinomycin ([M] = 1110.6315), respectively. While, streptodepsipeptide SV21 (
4), having a precursor ion of
m/z 1142.6804 [M + NH
4]
+, had not been reported yet in the MarinLit database. Further analysis, using the GNPS MASST database, on the precursor (MS
1) and product ion (MS
2) spectral data (
Figures S2–S5) showed that compounds
1–
4 were related to valinomycin with cosine scores ranking all above 0.7, while considering more than 40 major product ion peaks. This result indicated that
4 was also a valinomycin analogue.
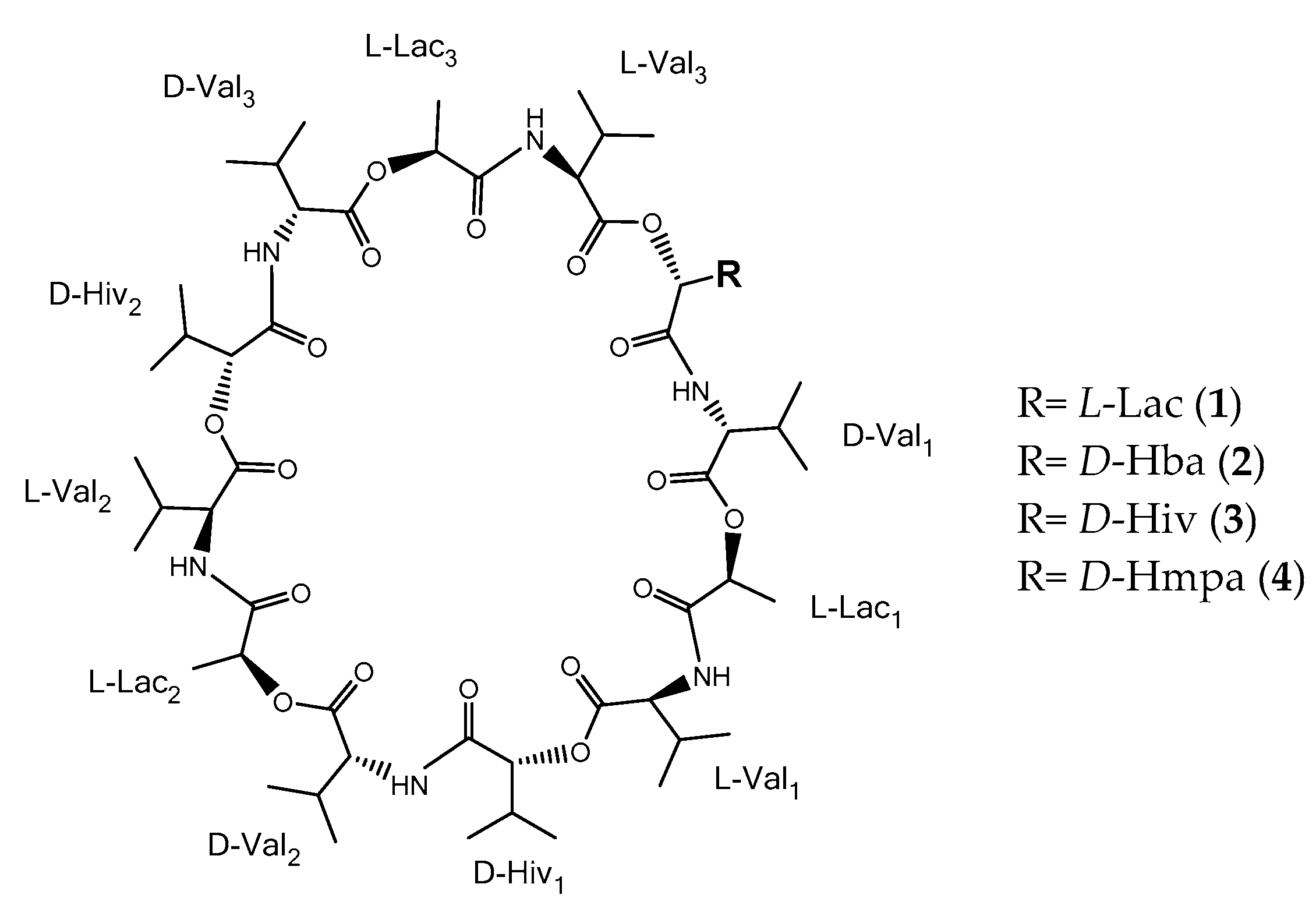
To have an overview of the structure for all four valinomycin analogues, we analyzed and compared the MS
2 spectra of compounds
1,
2, and
4 with valinomycin (
3). Valinomycin consists of the enantiomers
D- and
L-valine (Val),
D-α-hydroxyisovaleric acid (Hiv), and
L-lactic acid (Lac) [
6]. One valinomycin molecule consists out of three repeating units of Val–Hiv–Val–Lac, where one unit has an exact mass of 370.208 Da.
Compound identification started by comparing the precursor ion of each compound (cf.
Table 1) as well as the fragment ions in the mass region between
m/z 600 and
m/z 1150 (
Figure S6). Both, the precursor ion and the fragment ions showed a consecutive mass increase of 14 Da from compound
1 to
4. However, the fragment ion
m/z 713.4 was found to be in all valinomycin analogues. Based on the molecular mass, the fragment ion at
m/z 713.4 is represented by two units of Val–Hiv–Val–Lac after the loss of a unit of C=O for the initial opening of the ring structure. To crosscheck the existence of a unit Val–Hiv–Val–Lac in all compounds, the neutral loss of 370.208 Da was observed in all compound spectra after the loss of a C=O unit, which initially opened the ring structure. The neutral loss of two units of Val–Hiv–Val–Lac, with an exact mass of
m/
z 740.419 Da, was also observed in compounds
1–
4, indicating that all four molecular species contained at least two units of Val–Hiv–Val–Lac.
Despite the similar smaller fragment ions that occurred in the mass spectra at
m/z 50–
m/z 600, further analysis showed some unique fragment ions for each valinomycin analogue. For example, the fragment ion at
m/z 315.192 was only present in streptodepsipeptide P11B (
1). This compound represents a monomer and dimer of depsipeptide (Val–Hiv–Val–Lac) as well as a monomer of Val–Hiv–Val–Lac minus 28 Da. Based on the previous studies, the mass difference of 28 Da might be the result of the substitution of a Hiv with a Lac [
6,
7]. Therefore, the fragment ion at
m/z 315.192 is likely a unit of Val–Lac–Val–Lac. This is further supported by the neutral loss of 171 Da (Val–Lac) from
m/z 315.192 to
m/z 144.103 (shown as black arrow in
Figure S6A). The fragment ion at
m/z 144.103 is a unit of Val–Lac with a loss of C=O.
Another example is represented by the fragment ion at
m/z 329.208, which was only found in streptodepsipeptide P11A (
2). This compound has a mass of about 14 Da lower than a unit of Val–Hiv–Val–Lac. By comparing out the MS
2-based analysis with previous studies, there are at least two possible explanations: first, a substitution of a Hiv with a hydroxybutanoic acid (Hba) [
6]; or second, a substitution of a Val with an isoleucine or a leucine (Ile/Leu) in a unit of Val–Lac–Val–Lac [
7]. Both scenarios would be possible since the ion for Val–Hba–Val–Lac and Ile/Leu–Lac–Val–Lac create a fragment mass of
m/z 329.207. Furthermore, the fragment ion at
m/z 186.113 in
2 might be the protonated ion of Val–Hba or Ile/Leu–Lac. However, after the fragmentation pathway of the compound was simulated for both substitutions, a peak for Val–Lac–Val–Lac around
m/z 315.191 could not be found in the spectra, if a Val was substituted with Ile/Leu. Therefore, the most suitable substitution in
2 is a Hiv with a Hba, as also mentioned in [
6].
A fragment ion at
m/z 357.239 occurred in the spectra for streptodepsipeptide SV21 (
4). It has a molecular mass of 14 Da higher than a unit Val–Hiv–Val–Lac. In a previous study, the additional 14 Da were suggested to result from a substitution of a Val with either an Ile or Leu unit [
7]. However, when we simulated the fragmentation of Val–Hiv–Ile/Leu–Lac, we were unable to detect the peak at
m/z 158.118 in the MS
2 spectra. Fragmentation of Val–Hiv–Ile/Leu–Lac should have resulted in Val–Hiv and Ile/Leu–Lac with the calculated
m/z 172.133 and
m/z 158.118, respectively. Therefore, we propose the structure of
4 to have a substitution of Hiv with hydroxymethylpentanoic acid (Hmpa), since we detected the fragment ions
m/z 186.150 (Val–Hmpa) and
m/z 144.103 (Val–Lac), thus indicating a single unit of Val–Hmpa–Val–Lac. In summary, the difference between compounds (
1–
4) is only a substitution of a Hiv with either a Lac, a Hba or Hmpa within a single unit of Val–Hiv–Val–Lac (cf.
Figure 1).
The configuration of the compounds
1–
3 in
Figure 1 was assumed to be identical with [
6], while configuration of
4 was derived from the biosynthetic pathway of valinomycin. The structure of compounds
1–
4 were quite similar, therefore we agreed with [
7] that valinomycin and their analogues are derived from the same biosynthetic pathway. Biosynthesis of valinomycin is accomplished by nonribosomal peptides (NRPS) that are composed of two proteins, namely, VLM1 and VLM2. Those proteins are divided into four modules, each one responsible for incorporation of one unit of
D-Hiv,
D-Val,
L-Lac, and
L-Val. The depsipeptide chain (
D-Hiv–
D -Val–
L-Lac–
L-Val) is linked to the C-terminal iterative thioesterase (TE) domain at the last module in VLM2. The terminal TE domain controls the termination, release and cyclization of the growing chains in the biosynthetic process [
16].
Biosynthesis of
D-Hiv in valinomycin is occurring in module 1 in VLM1 [
16]. The study also explained that module 1 contains four functional domains: adenylation (A; designated as VLM1A1), hypothetical transaminase (TA), hypothetical dehydrogenase (DH2) and peptidyl carrier protein (PCP). Extracted NRPS codes from VLM1A1 did not yield any predictable substrate, leading to the assumption that VLM1A1 might have adapted to select and activate hydroxyl acids independently [
16]. However, the adenylation domain is a core of each module that recognize the cognate substrate [
17]. Therefore, feeding experiments using different substrates, i.e.,
D-Hiv,
D-Hba, or
D-Hmpa, are needed to verify which substrates are needed to produce valinomycin and its analogues. In turn, substitution of
D-Hiv with
L-Lac in streptodepsipeptide P11B (
1) could be explained as a variation in the linearity within modules 3 and 4 instead of modules 1 and 2 in one round of the tetradepsipetide assembly [
7].
Streptodepsipeptide P11B (
1) proved to be the known depsipeptide based on the comparison of the measured MS, NMR, and optical rotation data with the references [
6,
7]. To crosscheck the structure with the MS
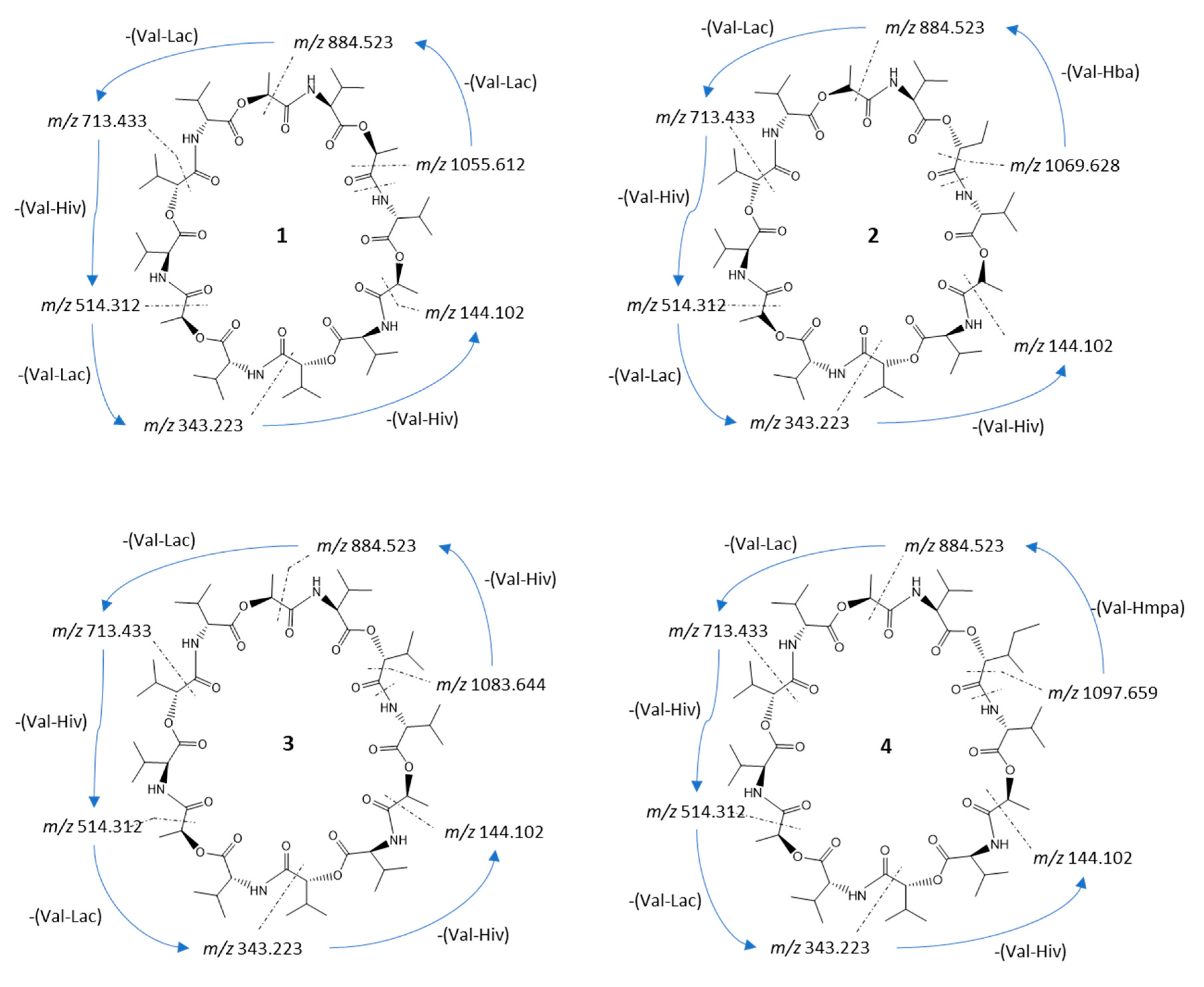
2 data, we simulated the fragmentation pathways for each compound. The fragmentation pathway of compound
1 started with the ring opening and the loss of a C=O unit of 27.995 Da. After the ring opened, the fragment ion
m/z 1055.611 (calc.
m/z 1055.612) (
Figure 2) continued to lose either a unit of Val–Lac or Val–Hiv with 171.089 and 199.121 Da, respectively (cf.
Figure S7). The sequential loss of a unit of Val–Lac or Val–Hiv explained the occurrence of fragment ions
m/z 884.520 (calc.
m/z 884.523) or
m/z 856.491 (calc.
m/z 856.491; fragment ions from MS
2 spectra
cf. Figure S7).
In total, the fragmentation pathway of streptodepsipeptide P11B (
1) took five major steps of losing either a unit of Val–Lac or Val–Hiv. However, since
1 has one substitution of Hiv with a Lac residue, one of the fragmentation steps is the repetition of losing a unit of Val–Lac, which is indicated by the consecutive tan arrows in
Figure S7.
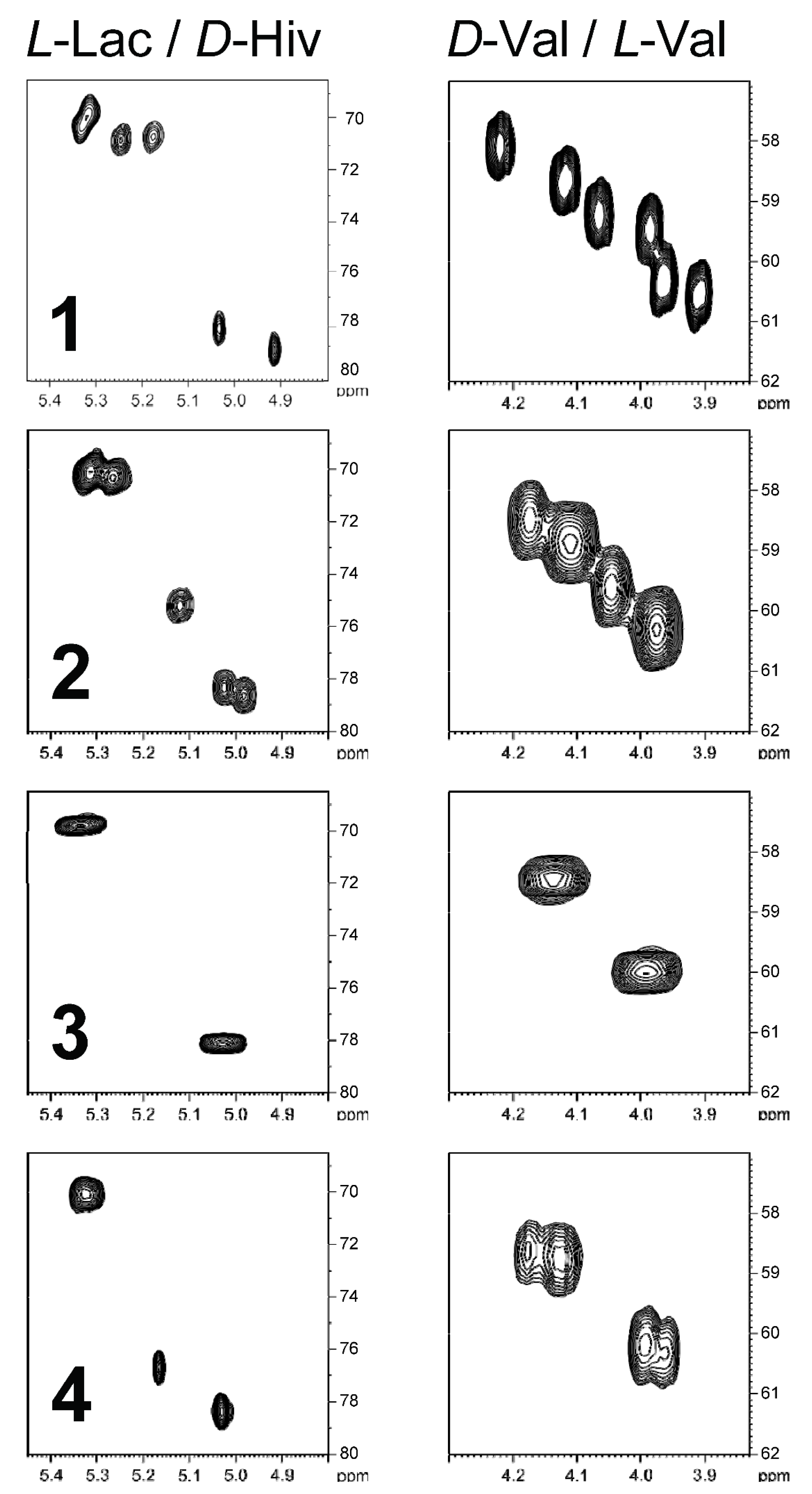
Besides a thorough mass spectral analysis, the four valinomycin analogues were also compared by their Hα/Cα region of the HSQC spectra (
Figure 3). Every residue showed a distinct fingerprint region in the HSQC spectrum and thus allows counting the different residues. For example, in the spectra of streptodepsipeptide P11B (
1), all 12 Hα/Cα correlations can be observed. The
D-Val/
L-Val plot indicates no changes since there are three signals for
L-Val and three signals for
D-Val. However, the plot on the left side indicates a loss of a Hiv residue and an increase of a Lac residue, which is in accordance with the molecular formula of
1 (valinomycin minus 2 × CH
2). In case of streptodepsipeptide SV21 (
4), there is quite some overlap in the four regions of the HSQC spectrum. Two of the three
L-Val and two of the three
D-Val residues overlapped. The same is true for the two Hiv residues. The three Lac residues all appear in one signal. The only signal without any overlap is the new Hmpa residue (5.17 ppm/76.7 ppm).
The precursor ion of streptodepsipeptide P11A (
2) was 1114.649 [M + NH
4]
+. It has 14 Da more compared to streptodepsipeptide P11B (
1). Most of the fragment ions of
2 also have a 14 Da difference to fragment ions of
1 (cf.
Figure S6). Therefore, the fragment ion at
m/z 1069.627 (calc. 1069.628) resulted from a neutral loss of a C=O (
Figure 2). The loss of a C=O may have happened anywhere in the structure; therefore, fragmentation of
2 then continued with a loss of a unit of Val–Lac, Val–Hiv, or Val–Hba at any positions near the opened ring. It explains the occurrence of the fragment ion
m/z 898.536 (calc. 898.538),
m/z 884.524 (calc. 884.523),
m/z 870.509 (calc. 870.507), etc. (cf.
Figure S8). Fragmentation pathways for
2 in
Figure S8 seems to be more complex than
1. However, the pathways still consist of five major steps of losing intermittently a unit of Val–Lac then Val–Hiv, or Val–Lac then Val–Hba, with only the loss of one Val–Hba for each possible pathway (blue arrows in
Figure S8).
The reported fragmentation pattern for streptodepsipeptide P11A (
2) in [
7] did not fit well with our measured fragment ions of compound
2. However, the NMR data for streptodepsipeptide P11A (
2) (
Figure 3) matched closely with the published data in [
6] and also resembled our fragmentation pattern (cf.
Figure S8).
The exact mass and also precursor ion (±0.01 Da) of the isolated valinomycin (
3) matched the reported mass data in [
6,
7,
9]. The fragmentation of
3 is initiated by the loss of a C=O at any position in the ring to produce the fragment ion
m/z 1083.643 (calc. 1083.644). The fragmentation process is then followed by the loss of a unit of Val–Lac or Val–Hiv. Following the same five major fragmentation steps as for streptodepsipeptide P11B (
1) and P11A (
2), fragment ions
m/z 912.549 (calc. 912.554),
m/z 884.524 (calc. 884.523),
m/z 713.433 (calc. 713.433), etc., were detected (cf.
Figure S9).
Streptodepsipeptide SV21 (
4) had a precursor ion of
m/z 1142.680 [M + NH
4]
+ and did not match any reported data in MarinLit. However, MS
2 analysis of
4 using the MASST GNPS database showed that
4 was strongly related to valinomycin based on a cosine score of 0.77 and 42 shared peaks (
Figure S5). A cosine score has a value between 0 and 1, with 1 indicating 100% similarity. A sample is considered an analogue of a reported compound if the cosine score is >0.7 [
18]. Several measured peaks of
4 were identical to the fragment ions of valinomycin (
3) (visualized by green lines, see
Figures S2–S5). Fragment ion
m/z 1097.660 (calc.
m/z 1097.659) resulted from the loss of a C=O group via ring opening. The fragmentation process then followed the previous reported pattern via loss of Val–Lac, Val–Hiv, or Val–Hmpa, resulting in peaks of
m/z 926.566 (calc.
m/z 926.570),
m/z 898.540 (calc.
m/z 898.538), and
m/z 884.523 (calc.
m/z 884.523), respectively (cf.
Figure S10).
For the
13C and
1H NMR assignments of valinomycin (
3), please see
Figure 3. The structure of valinomycin consisted of four units:
L-Val (appr. 60 ppm),
D-Val (appr. 59 ppm),
L-Lac (appr. 71 ppm), and
D-Hiv (appr. 79 ppm). Each unit showed characteristic NMR signals, which were useful for the characterization of the analogues (
1,
2, and
4). The
1H NMR spectra showed the purity of compounds
1–
4 and allowed the comparison with the reported compounds from [
6] (cf.
Figures S11–S15 and Tables S1–S4).
The
13C NMR signals for streptodepsipeptide SV21 (
4) displayed 55 carbon signals for 12 carbonyls (δ
C 169.9–172.4), six oxymethines (δ
C 19.1–19.3), six nitrogenated methines (δ
C 118.0–118.8), nine methines (δ
C 19.0–26.1), a methylene (δ
C 14.0), and 21 methyls (
Table 2 and
Figure S16). The
1H NMR spectrum of
4 showed six signals for NH at δ
H 7.88, 7.83, 7.80, 7.75, 7.69, and 7.67 ppm (
Table 2 and
Figure S17). Those data indicated that
4 consisted of 6 esters and 6 amino acids residues.
The MS results clearly indicate that compound
4 has an additional methylene group compared to valinomycin (
3). In principle, the additional CH
2 group could be added to each of the four residues. The inspection of the four Hα/Cα regions in the HSQC spectrum (cf.
Figure 3) clearly shows a loss of a Hiv residue, which means that the extra CH
2 group of
4 was added to this residue. There are two possibilities how to add a methylene group: First, a transition from Hiv to Hmpa would be possible. Second, from an “amino acid” point of view, it could be the transition from Val to Leu or Ile. However, the Hmpa unit was established by analysis of the COSY and HMBC spectra. The complete spin system of Hmpa could be assigned by the TOCSY spectrum without the determination of the explicit positions. Starting from Hα, the Hβ can be assigned by the COSY correlation, and the corresponding Cβ (36.7 ppm) is accessible by the HSQC (
Figure 3 and
Figure S18). Furthermore, three HMBC correlations can be observed starting from Hα (36.7 ppm, 26.1 ppm, and 14.0 ppm). This already indicates the existence of a Hmpa residue. For the constitutional isomer “Leu” only two HMBC correlations would have been expected and no correlation to a methyl group (here 14.0 ppm). The complete assignment of the Hmpa residue is given in
Table 2.
Valinomycin (
3) and its analogues (
1,
2, and
4) showed a narrow spectrum of antimicrobial activities (
Table 3). Compounds
1–
4 showed that all antifungal activity against
Mucor hiemalis (Mh) and
Ruegeria glutinis (Rg), with streptodepsipeptide P11A (
2) and valinomycin (
3) being the most active ones, revealed similar or lower MIC values compared to the commercial antifungal compound nystatin (cf.
Table 3). Valinomycin was eight times stronger than nystatin against
Mh and as strong as nystatin against
Rg. Previous studies had also shown antifungal activity of valinomycin against the plant pathogens
Phytophthora capsici and
Botrytis cinerea [
13,
14]. Only valinomycin (
3) and streptodepsipeptide SV21 (
4) also exhibited activities against the Gram-positive bacterium
Staphylococcus aureus (Sa) and
Bacillus subtilis (
Bs) (only (
4) had activity). However, the activity of valinomycin on
Bs was strongly affected by the pH regime in the conducted assay. At the different pH values from 5.5 to 9.5, valinomycin showed an increase in antibacterial activity against
Bs at higher pH values [
19]. To the best of our knowledge, this is the first report on the antimicrobial activity of compounds
1,
2, and
4.
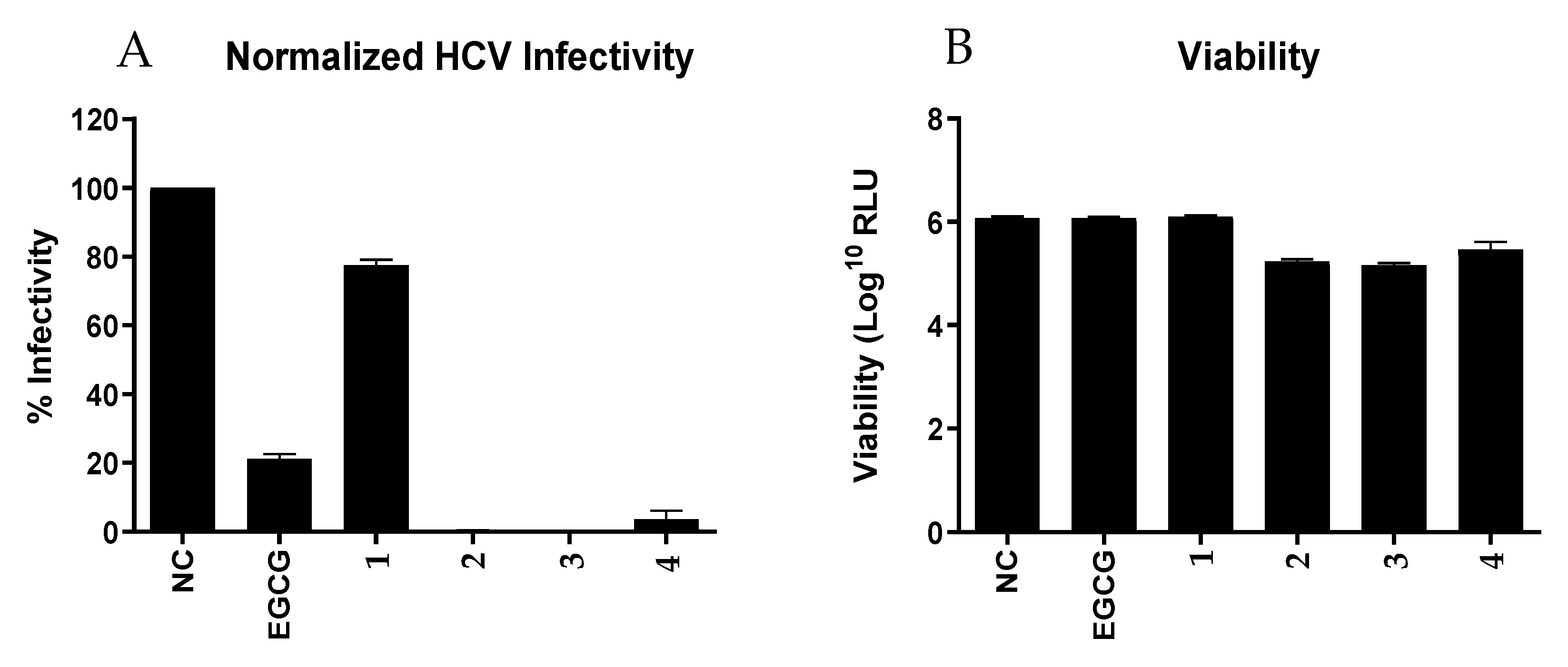
Furthermore, we tested all 4 compounds (
1–
4) against the Hepatitis C Virus (HCV,
Figure 4). Although structurally closely related, only the compounds
2–
4 showed pronounced infectivity against HCV compared to the positive control epigallocatechin gallate (EGCG). Streptodepsipeptide P11B (
1) significantly less affected the HCV. This finding indicated that valinomycin (
3) and its analogues (
2 and
4) have a strong potential to function as potent ani-HCV agents. However, viability of the Huh7.5 cells for valinomycin (
3) and its analogues (
2 and
4) were lower than for the positive control, meaning these compounds also slightly affected the host cell as well.
The isolation and structure assignment of valinomycin and the three derivatives, including the new streptodepsipeptide SV21 (
4), allowed us for the first time to conduct a structure activity relationship (SAR) analysis to determine the essential functional groups for the observed antimicrobial and newly reported antiviral activities. The symmetry of the ring system seems to be key for the activity of valinomycin and its analogues. Compounds
1–
4 have the same number of carbonyl groups and share the same two units of Val–Hiv–Val–Lac. The difference is only in one unit of the depsipeptide, which affects the symmetry of the molecule. Valinomycin affects the cells by dissipating the electrochemical gradient, which is essential for cell life through influx of the potassium ions into the cell. Valinomycin can change its conformation, allowing it to dissolve in aqueous but also lipophilic environments. The carbonyl groups in valinomycin form the hydrophilic site, while the methyl and isopropanyl groups form the hydrophobic site. A previous study on the antibiotic mechanism of valinomycin indicated that potassium was released from the hydrophilic site after forming a complex with valinomycin. At the hydrophobic membrane interface, the potassium ion is selectively released through the substitution with water molecules at the Lac rather than Hiv site [
20].
Valinomycin (3) was the most active of the four tested compounds. Deletion of one or two CH2 groups in the valinomycin structure resulted in weaker antifungal activity, while addition of a CH2 group gave weaker antifungal but broader antibacterial activity. Thus, it seems likely that the symmetry of valinomycin is important for the observed higher bioactivity.
The antiviral activity of valinomycin (
3) and its analogues (
2 and
4) against HCV amplifies the potential of these compounds to be developed as or used a scaffold for the development of anti-viral agents. In a previous study, valinomycin also showed promising activity against the SARS-CoV virus [
21]. Therefore, it will be interesting to conduct experiments with valinomycin and its analogues against the new strain of Coronavirus (SARS-CoV-2).


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}