1. Introduction
Whiteleg shrimp,
Litopenaeus vannamei, is one of the world’s largest farmed tropical shrimp species native to the Pacific Ocean [
1]. The shrimp is characterized by rapid growth, high tolerance to a wide range of water salinity (5–40 ppt), high intensification, low protein requirements, and other features suitable for super intensive aquaculture [
2]. Nevertheless, several environmental factors affect the shrimp growth and survival rates, such as water temperature [
3], salinity fluctuation [
4], sharp and rapid changes in pH [
5], low dissolved oxygen [
5], and toxins such as ammonia, nitrite, hydrogen sulfide, and heavy metals [
6].
The worldwide progress of shrimp farming is facing rising environmental and pathological difficulties [
7], besides low capability for cold water tolerance in most shrimp producing countries. Using natural immunostimulants as a healthier/safer therapy than administrating antibiotics and/or vaccines is recommended to control aquaculture pathogens [
8]. Increasing shrimp immunity is an important direction in the current supporting policies in order to prevent shrimp diseases [
7].
Nucleotides are chemical compounds that played chief roles in nearly all biological cellular metabolic processes like energy production (e.g., ATP), metabolism, cell signaling (e.g., cAMP). They provided the natural building block of DNA, RNA, and essential cofactors in the biosynthesis of proteins and enzymes (e.g., NAD+) [
9,
10]. It is supposed that crustaceans have inadequate ability to synthesize nucleotides de novo. Therefore, it would be highly beneficial to support shrimp feeds with dietary supplementation of nucleotides [
11], especially during the stressful periods. Though nucleotides’ usage is relatively novel in shrimp aquaculture, there is growing evidence exhibiting their benefits [
12].
Marine plants (seaweeds or macroalgae) are considered a rich source of bioactive compounds [
13,
14]. There is a rising awareness of the exploitation of seaweeds (dried, isolated compounds and/or extracts) as an aquafeed additive in shrimp farming. Seaweeds (red, brown, and green) are characterized by many properties such as nutritional, growth-promoting, cytostatic, antioxidant, immune-modulatory, neuroprotective, hepatoprotective, antibacterial, antifungal, and antiviral activities [
15,
16,
17,
18,
19].
The seaweed genus,
Sargassum sp. is the most important, common, and widespread brown seaweed that is found in tropical and subtropical regions. They have been used in agriculture, the food/feed industry, folk remedy, and cosmetology [
17,
20]. Various Sargassum species have folk applications in human food as a rich source of proteins, carotenoids, vitamins, and minerals [
17].
Sargassum sp. has ecological and economic importance in producing nutritional feed additives and different medicinal products. Many species are found in Egypt; some were identified, and many more need to be identified [
21].
Sargassum polycystum C. Agardh is brown seaweed abundant in the Red Sea, Hurghada, Egypt.
S. polycystum (sp.) is traditionally used against several human diseases [
22].
The biochemical composition of
S. polycystum revealed the nutritive value of crude protein (14.2%), crude lipid (7.6%), fiber (21.3%), carbohydrate (25.0%), polyunsaturated fatty acids, soluble nutritional fibers, and ash (29.0%) higher than that documented in terrestrial plants and animal products [
23]. Sargassum is the richest source of fucoidans and alginic acids [
24]. Several bioactive compounds were extracted from different Sargassum species as polysaccharides, alkaloids, tannins, saponins, flavonoids, glycose, sulfated polyphenols, phenolic, sterols, B-carotene, terpenoids, sargaquinoic acids, sargachromenol, and pheophytin [
17,
25,
26]. Chotigeat et al. [
27] stated that the extracts from several brown algae species containing a polysaccharide element had an effective capability to improve the disease resistance and immune responses of different aquatic species. In addition, alginic acid [
28] and fucoidan [
29] extracted from brown algae showed a useful role as an immune induction in preventing various aquatic diseases [
30]. The immunity parameters, such as total hemocyte count and phagocytic activity, are well-documented [
31,
32].
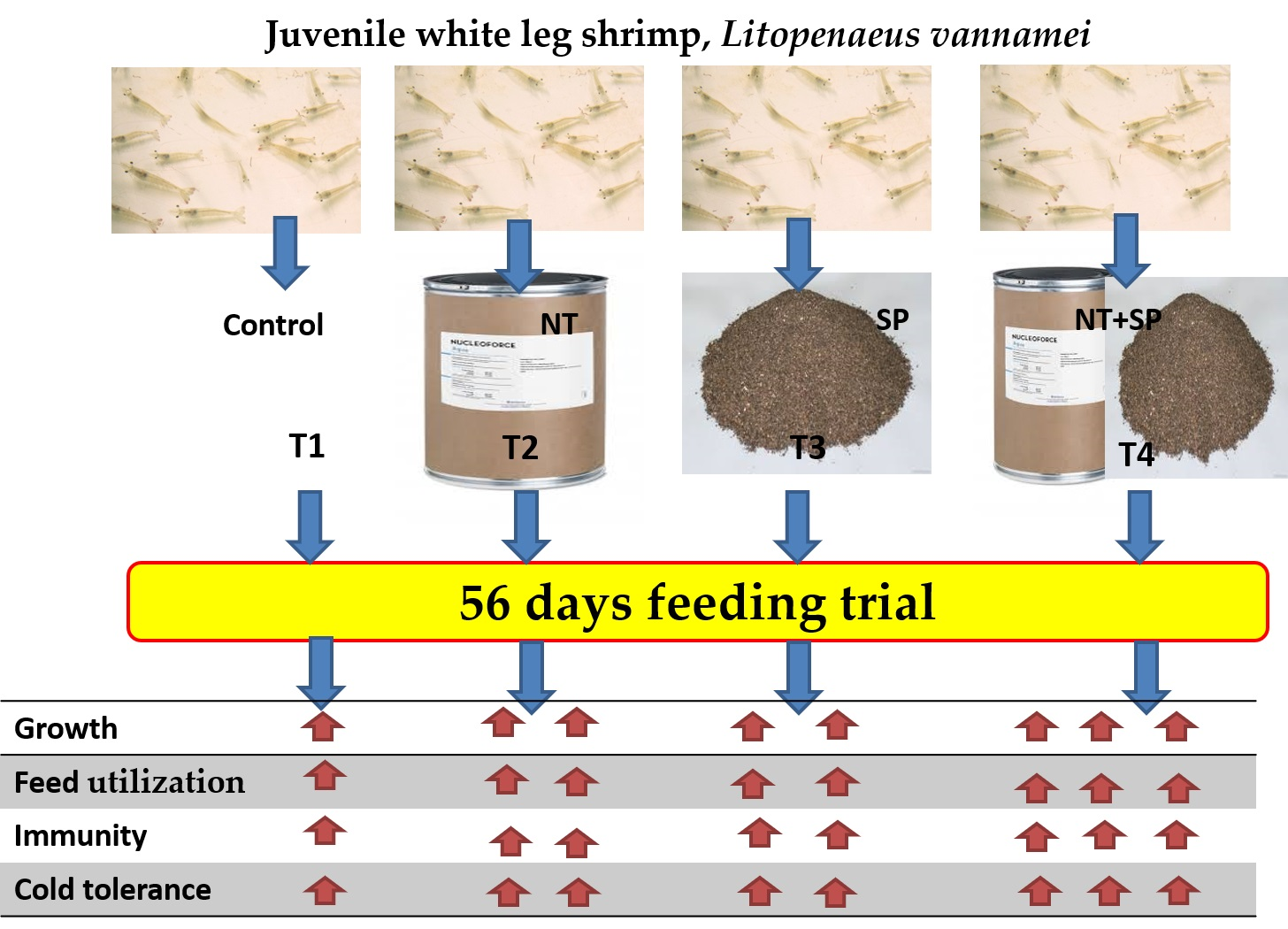
The winter months in Egypt and many other Mediterranean countries (December, January, and February), with an average day/night difference of 10–16 °C, are the main drawback for continuous aquaculture production of shrimps. The cold winter caused massive economic death to the L. vannamei aquaculture sector. However, insignificant data is available about shrimp responses during cold temperature stress. In the present study, the effects of nutritional additives on the cold tolerance of whiteleg shrimp were studied. Accordingly, this study investigated the single and synergistic effects of dietary supplementation of brown seaweed, S. polycystum, and nucleotides (NucleoforceFish™) on water quality, growth, feed utilization, proximate composition, immune responses, phagocytic activity, histology of hepatopancreas, and intestinal tract, besides antioxidant, antimicrobial, and cold-tolerant gene expression in juvenile whiteleg shrimp, during the winter season. The results could deliver valuable orientation towards the importance of dietary management through the use of feed additives as a potential method to adapt to or face the winter season.
3. Discussion
Modern feed additives and their impact on improving water quality, survival rates, feed utilization, health status, and disease resistance of cultured aquatic organisms are critical issues that scientific research focused on during recent decades. The direct/indirect effects of dietary nucleotides and seaweed on water quality have not been focused on much before. The present study revealed a significant decrease in both TAN and NH3, parallel with the increase in dissolved oxygen concentration. This might be attributed to the better utilization of offered feeds in the diets T2, T3, and T4, compared with T1. The minimum averages of water temperature recorded in this study were higher than the lethal limits stated by [
33]. They found that the cold tolerance level of
L. vannamei varied between 7.5 and 11 °C and it seems to be more sensitive to low water temperatures than other penaeid species. Additionally, the previous authors stated that for successful commercial aquaculture, water temperature must never fall below12 °C during the overwintering season to avoid mortalities.
The indirect effects of marine algae on water quality in shrimp ponds are illustrated by [
34], who tested the effects of coculture of two species of macroalgae separately (
Gracilaria vermiculophylla and
Dictyota dichotoma) in
L. vannamei ponds, and recorded a significant decrease in the concentration of both ammonia and nitrites.
The growth rate of a living organism is a measure of the extent of utilization of the feed provided. In the present study, it was found that the experimental diets containing
S. polycystum and nucleotides induced a clear improvement in growth performance, especially the T4 treatment receiving both additives. This result is in agreement with [
35] concerning
Sargassum addition in
M. rosenbergii, and [
9,
12,
36,
37], regarding the addition of the nucleotide. Arizo et al. [
35] concluded that dietary supplementation of fucoidan’s extracted from
S. polycystum at 500 mg kg
−1 exhibited the highest increment on growth performance parameters. The
P. monodon, PL15–35 fed
Artemia instar II enriched with various concentrations of
Sargassum sp. extracts at 250, 500, and 750 mg/L showed significantly higher growth performance indices than the control group; the best concentration was 750 mg/L [
38]. Moreover,
Penaeus indicus juveniles fed with
Sargassum wightii-enriched
Artemia nauplii exhibited higher weight gains and SGR than the control after 30 days of feeding trial [
39].
For nucleotides, Xiong et al. [
40] found that shrimp,
L. vannamei fed 50 g/kg nucleotides-enriched diet displayed significant higher growth rates compared to nonsupplemental diet. Similarly, Lucien-Brun and Vidal [
10] stated that shrimp fed the nucleotides-supplemented feed had 23.2–25.0% higher final weight than the control group in normal condition and 57.7% after infection with white spot syndrome virus. Andrino et al. [
37] recorded higher values in shrimp gain and growth indices of
L. vanemmei fed with a nucleotide-supplemented diet than those fed the control diet. However, Schleder et al. [
18] did not find a significant effect of
S. filipendula dry biomass’ dietary addition on the growth performance of Pacific white shrimp at concentrations 0.5–4%. The effect of seaweed on the performance of shrimp was significantly different. It depended on the species used, the processing technique of the extract, the quality of feed, the content of fishmeal in the diet, and the inclusion level [
18,
41,
42].
The lower FCR, the more effectively fish utilizes the aquafeed nutrients [
43]. In the present study, the decrease in the FCR values obtained in the
Sargassum-enriched diet (T3) and nucleotides-enriched diet (T2) is consistent with the obtained results by [
35,
37], respectively. Arizo et al. [
35] stated that fucoidan’s dietary addition extracted from
S. polycystum at 500 mg kg
−1 recorded the lowest values of FCR. Schleder et al. [
18] noted that seaweed’s dietary supplementation in the shrimp diet improved nutrient absorption, feed utilization, and digestive tract development. Additionally, Chithambaran and David [
44] found that plant extracts stimulated shrimp digestive enzymes. Furthermore, nucleotide-fed crustaceans exhibited better FCRs and more efficient utilization of proteins and lipids [
37,
40]. Xiong et al. [
40] indicated that shrimp fed 50 g/kg nucleotides-enriched diet showed significantly higher PER, PPV, and lower FCR than a nonsupplemental diet. Similarly, Lucien-Brun and Vidal [
10] recorded that shrimp fed the nucleotides-supplemented feed had 27.3% better FCR than the control one.
For the present study’s survival rate, nutritionally enriched groups increased survival rate compared to the control one. This is consistent with the other researchers regarding
Sargassum additives [
35] and nucleotides [
9]. This might be attributed to the fact that nucleotides/
Sargassum supplements improved innate and adaptive immune systems, reducing pathogenic infections [
9], and tolerated more efficiently cold-water temperatures as recorded in the present study. Shrimp-fed nucleotides-enriched diet exhibited higher survival rates during different normal growing stages [
45,
46] and violent environmental changes conditions like changes in salinities [
47]. Similarly, Lucien-Brun and Vidal [
10] recorded that shrimp fed the nucleotides-supplemented feed had a 14.3–38.5% higher survival than the control and 63% higher after infection with white spot syndrome virus.
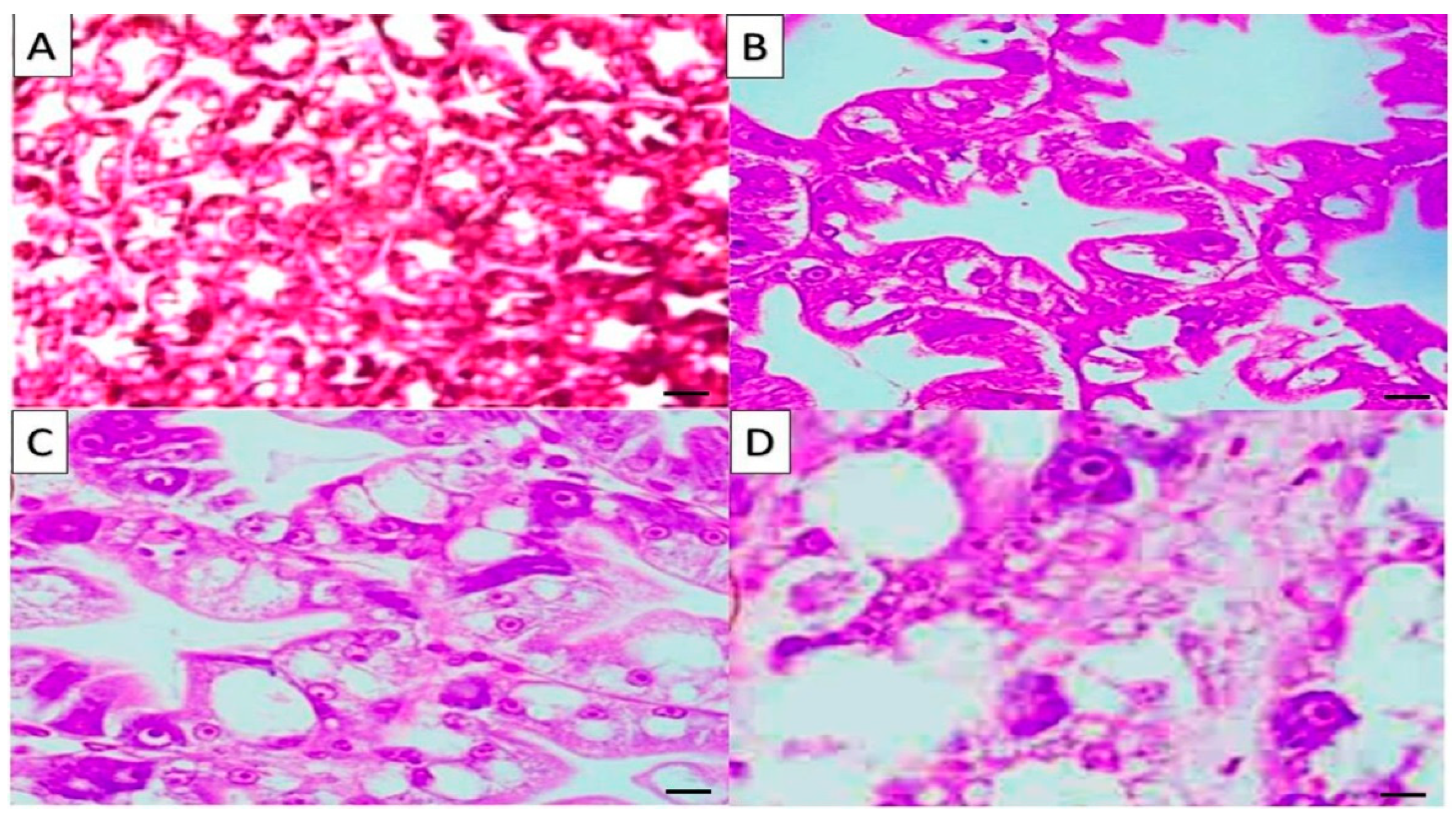
In the present study, whole-body proximate composition indicated a significant decrease in lipid and protein content in shrimp fed
Sargassum and nucleotides diets. In
Sargassum fed group, this might be attributed to the high level of amylase activity and the lower activity of the proteolytic enzymes, as stated by [
18] who found that the dietary addition of seaweeds,
S. filipendula did not increase proteolytic enzymes but increased amylase activity significantly. The
S. polycystum extracts have hepatoprotective and antioxidant properties, enhancing insulin sensitivity in diabetic rats [
48]. The low doses of
S. polycystum extract at 150 mg/kg body weight were useful in improving histological damages in diabetic tissues and organs. In comparison, the concentration of 300 mg/kg body weight was helpful to the pancreas but may be poisonous to the kidney and liver of diabetic rats [
48]. Therefore, with higher insulin content, a direct relation between the lower content of lipids and the dietary inclusion of
Sargassum might be explained. For nucleotides fed diet, the authors of [
40] found no significant differences in the whole body and proximate muscle composition of
L. vannamei fed nucleotides-rich yeast diets at doses of 10 and 30 g/kg. However, the higher dose of 50 g/kg increased the content of protein significantly. Abedian-Kenari and Oujifard [
49] found that dietary nucleotides did not significantly change the chemical composition (moisture, protein, SFA, MUFA, PUFA, and ash) in
L. vannamei. Similar results were observed in white shrimp [
50].
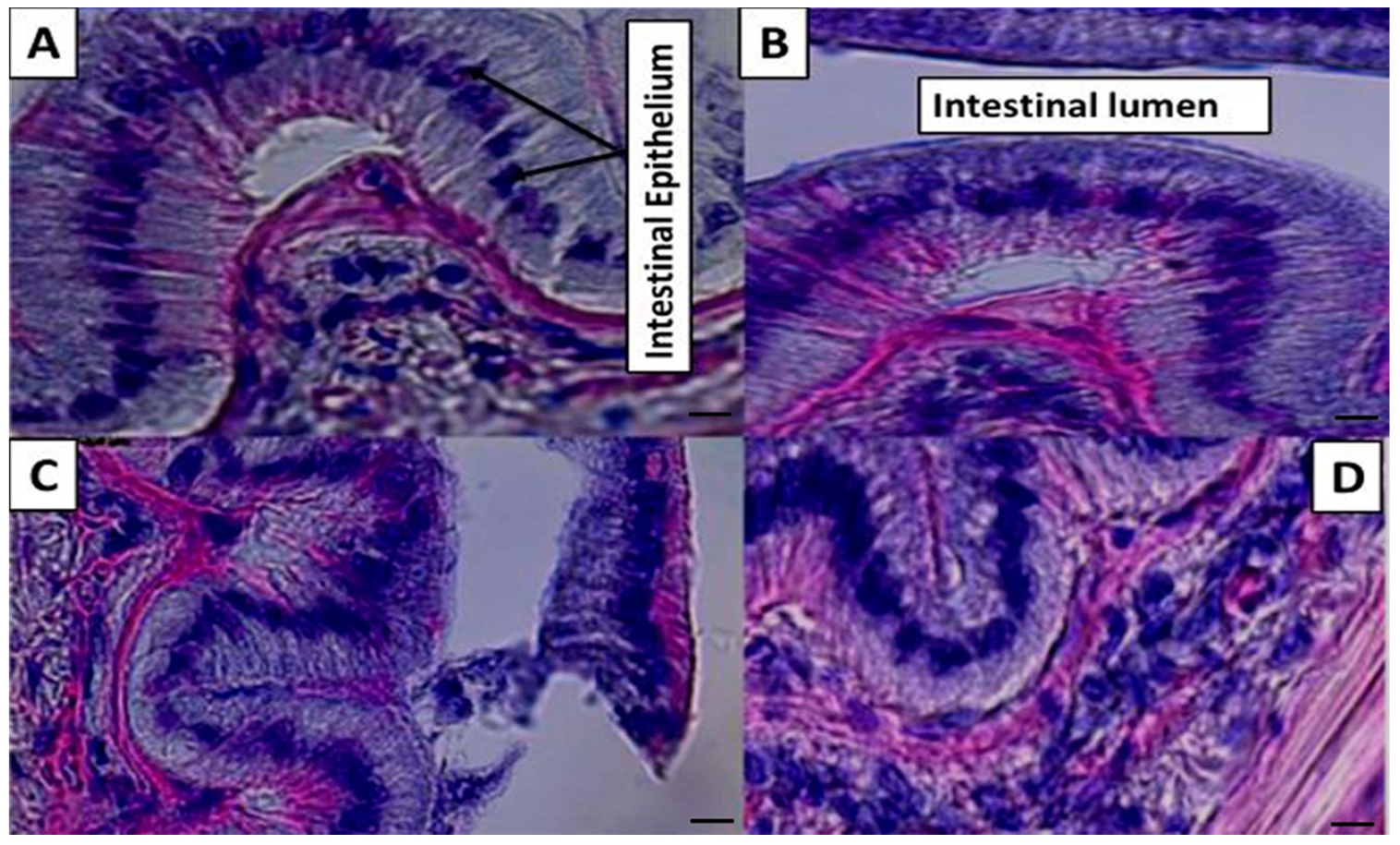
The nutritional quality of shrimp diets positively or negatively affects the morphology, amelioration, and functional health status of the gastrointestinal (GI) tract [
40,
50]. In the present study, the dietary inclusion of both
Sargassum and nucleotides improved the intestinal epithelium histology and lumen with more cells and width. The current study results are alike with earlier studies for shrimp [
12,
40,
50] regarding the positive effects of nucleotides. Dietary nucleotides have a constructive impact on intestinal growth [
12], intestinal villi height, and jejunum wall thickness [
50]. This development led to an increase in the shrimp gut’s total mucosal surface [
51]. Moreover, for
Sargassum, the authors of [
18] concluded that the diet supplemented with 0.5%
S. filipendula (dry biomass) significantly enlarged the surface area of the gut absorption epithelium of
L. vannamei. According to [
52], the dietary addition of
Gracilaria lemaneiformis at 2% dry biomass improved integrity of GI tract. It enlarged the length of microvilli cells in the gut absorption epithelium of
L. vannamei. Likewise, the dietary addition of 2%
U. pinnatifida (dry biomass) enlarged the length of intestinal villi in
P. monodon juveniles [
53].
Shrimp has a deficiency in the adaptive/specific immune system and depends on the innate/nonspecific immunity for protection against diseases and environmental changes [
45]. Feed additives can stimulate both the humoral and cellular contents of the innate immune system [
54]. Activities of lysozyme, total hemocytes, AP, SOD, PO, and TNO are vitally important nonspecific immune parameters reflecting the innate immune capacity [
40]. The present study revealed that the dietary administration of both nucleotides and
Sargassum significantly increased all the nonspecific immune responses with different levels of improvement. The superior results were in favor of T4. The results of this study are in agreement with many previous studies, both in terms of the positive effect of nucleotides [
9,
40,
50,
55] or
Sargassum [
25,
31,
56]. For nucleotides, shrimps fed dietary nucleotides-supplemented diets showed significantly higher lysozyme activities, T-SOD, and T-NOS when dietary nucleotides added at levels from 60 to 120 mg/kg, and activities decreased when the level of addition increased to 1200 mg/kg [
50]. Similarly, the authors of [
40] manifested that shrimp fed nucleotides-rich yeast diet at 50 g/kg indicated higher PO and lysozyme activities than the control group of
L. vannamei. The administration of dietary nucleotides increased respiratory burst activity, PO activity, and THC [
37].
For
Sargassum, flavonoids are one of the compounds that play a significant role in enhancing the immune system. In this regard, in [
25] the authors concluded that shrimp immersed in 150 ppm
Sargassum extract showed proliferation of hemocyte and other hemolymph parameters. Schleder et al. [
57] stated that the dietary addition of 4% of
U. pinnatifida improved PO activity in
L. vannamei. Ghaednia et al. [
7] found that the content of total hematocytes count (THC), phagocytic activity (PA), differential hemocyte count (DHC), total plasma protein, bactericidal activity, and bacterial clearance efficiency increased significantly in
F. indicus immersed in hot seawater containing
Sargassum glaucescens extracts at 300 and 500 mg/L compared with the control. Phagocytic index, phagocytosis %, ABPC increased in shrimp fed with fucoidan and white spot syndrome virus (WSSV) infected 2.84-, 1.66-, and 1.55-fold compared with the control [
8].
S. polycystum exhibited the best antimicrobial activity and the best natural immunostimulant compared to other
Sargassum species (
S. oligocystum,
S. crassifolium,
S. cristaefoliumas) against tested aquaculture microbial diseases [
19].
Environmental stress negatively affects the shrimp immune system. Stressors increase shrimp vulnerability to diseases, probably due to long-lasting raised cortisol content, leading to immunosuppression [
21]. Le Moullac and Haffner [
58] observed that
P. stylirostris showed decreased THC after exposure to ammonia content at 3 mg/L. The effects of low water temperature on the immune system of
L. vannamei have not been studied yet. However, increasing water temperature from 27 to 33 °C significantly increased the total hemocyte count in
L. vannamei hemolymph infected orally with WSSV, resulting in 100% survival [
59]. However, the higher temperature might increase
Vibrio bacteria populations in shrimp farms [
60].
Shrimp diseases caused by viruses, particularly WSSV, and cold-water tolerance are the highest shrimp aquaculture challenges. A good understanding of shrimp immune response may help control these challenges [
61]. The modulation of the immune response in shrimp farming through various marine seaweeds and/or prebiotics has become the main direction of scientific studies [
40,
62,
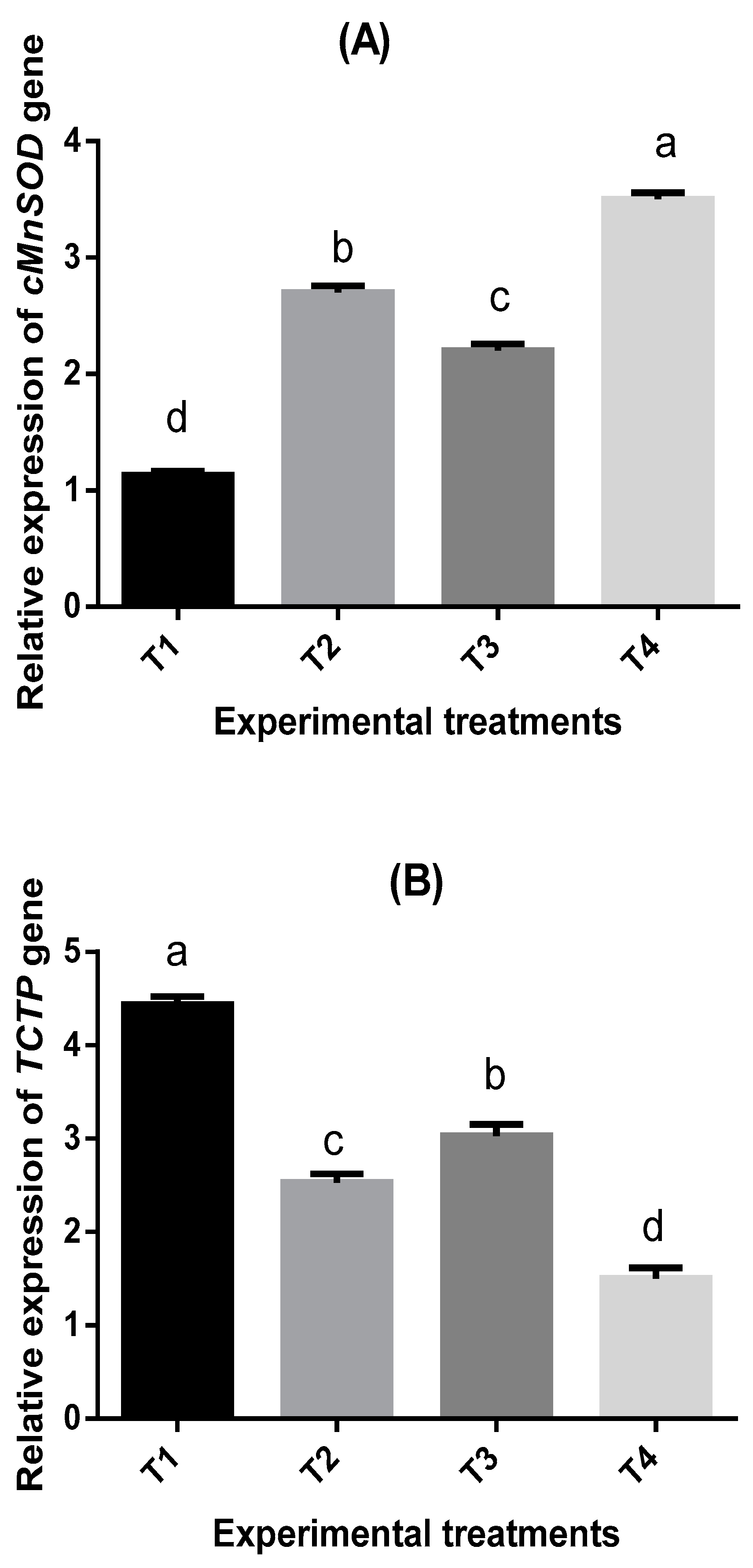
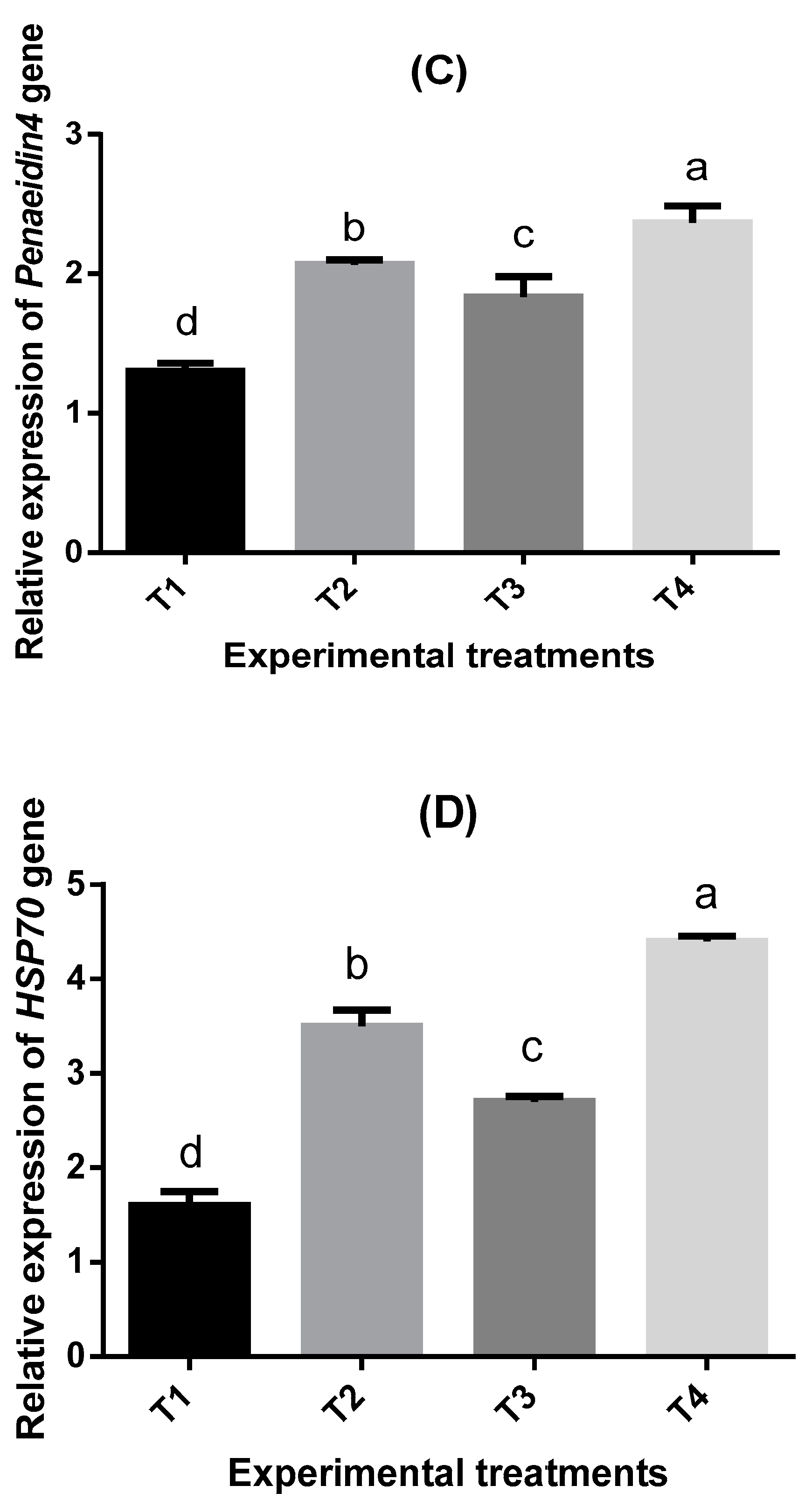
63]. Studies on the relationship between nucleotides/seaweeds-enriched diets and gene expression in shrimps are minimal, whereas the effects of feed additives on gene expression under low water temperature have not been studied yet. Therefore, one of the main objectives of this study was to address this challenge. The present study findings showed that nucleotides/seaweeds-supplemented diets upregulated the relative expression of cMnSOD, Penaeidin4, and HSP70 genes, while TCTP was downregulated. The results agreed with the previous study for
Litopenaeus vannamei fed
Aloe vera [
63], who reported similar findings in the immune-related genes (SOD, HSP70, penaeidin4, and TCTP). The addition of nucleotides in shrimp feeds displayed the maximum values of alkaline phosphatase and lysozyme relative genes expression when
L. vannamei were fed 30 g/kg nucleotides-rich yeast supplementation compared with all treatments [
40].
Heat shock protein (HSP70) is an accessory protein that has a vital role in the immune response as cheaper ones and inducers of proinflammatory cytokines secretion [
64]. In
P. vannamei shrimp, HSP70 has been documented [
65]. It existed at low concentrations in many cells under stressful conditions, irrespective of their cell cycle stage [
63]. The infection with bacteria or viruses caused an elevation in HSP70 level in
P. vannamei [
66].
Some plant compounds such as sodium alginate [
67],
Panax ginseng extract [
68],
Rubus coreanus extracts [
69],
A. vera [
63] enhanced
SOD gene expression in different species of shrimp. Additionally, environmental stressors such as abrupt changes in water temperature increased
MnSOD gene expression. González-Ruiz et al. [
70] concluded that high water temperature and hypoxia exhibited a synergistic effect in the upregulation on gene expression of
mMnSOD in both gills and hepatopancreas. On the other side, the upregulated production of
MnSOD gene expression in shrimp was related to pathogens infection and defense processes, as detected in
P. monodon challenged with
V. harveyi [
71].
Microbial infections seem to be the main inducer of
Penaeidin4 gene expression [
72], especially against Gram-positive, not Gram-negative bacteria [
73]. Nevertheless, Wang et al. [
74] discovered that
penaeidin4 was upregulated in
P. vannamei challenged with Gram-negative bacteria like
V. harveyi. The same trend was detected in
F. indicus infected with
V. parahaemolyticus [
75]. On the other hand, some nutritional immunostimulants had an influential role in
Penaeidin4 gene expression. An increase in
Penaeidin4 gene expression due to the dietary addition of
A. vera [
63,
76] was detected.
Downregulation of
TCTP might be attributed to low water temperature.
TCTP played essential multifunctional roles in cell growth [
77], eukaryotes [
71], and cell death pathway in shrimp hemocytes [
78]. Bangrak et al. [
78] stated that
TCTP gene expression was decreased in
P. monodon infected with a high WSSV burden. However, a study carried out by [
61] concluded that
TCTP was upregulated after WSSV challenge test.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}