
Algae-Derived Anti-Inflammatory Compounds against Particulate Matters-Induced Respiratory Diseases: A Systematic Review

 ,
, 
 , , ,
, , ,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
3. Discussion
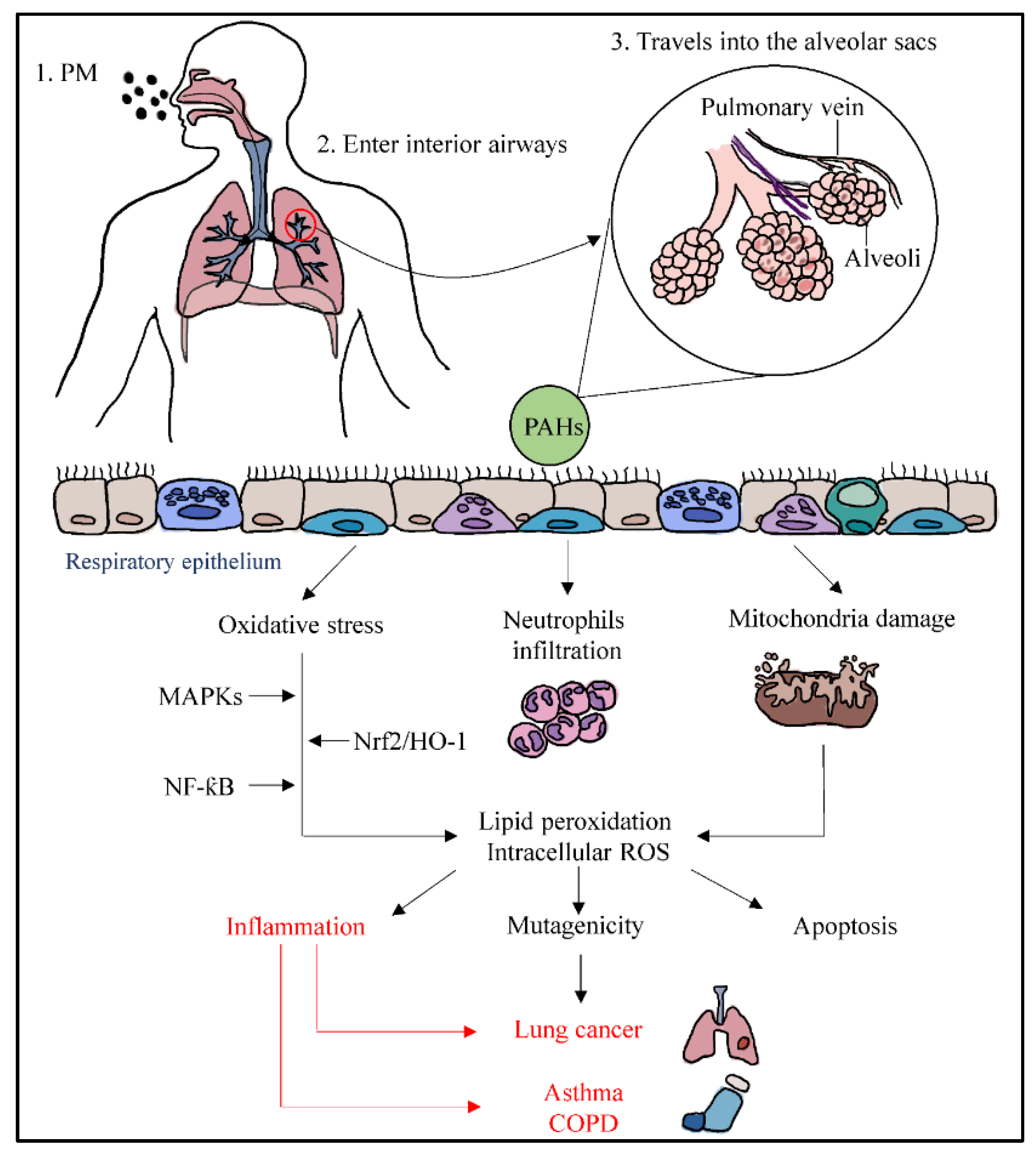
3.1. Pathways of Particulate Matter Travelling into the Respiratory System and Causing Inflammation
3.2. Algae Metabolites with Anti-Inflammatory Activity against Air Pollutants
3.2.1. Phytosterols
Fucosterol
3.2.2. Polysaccharides
3.2.3. Polyphenols
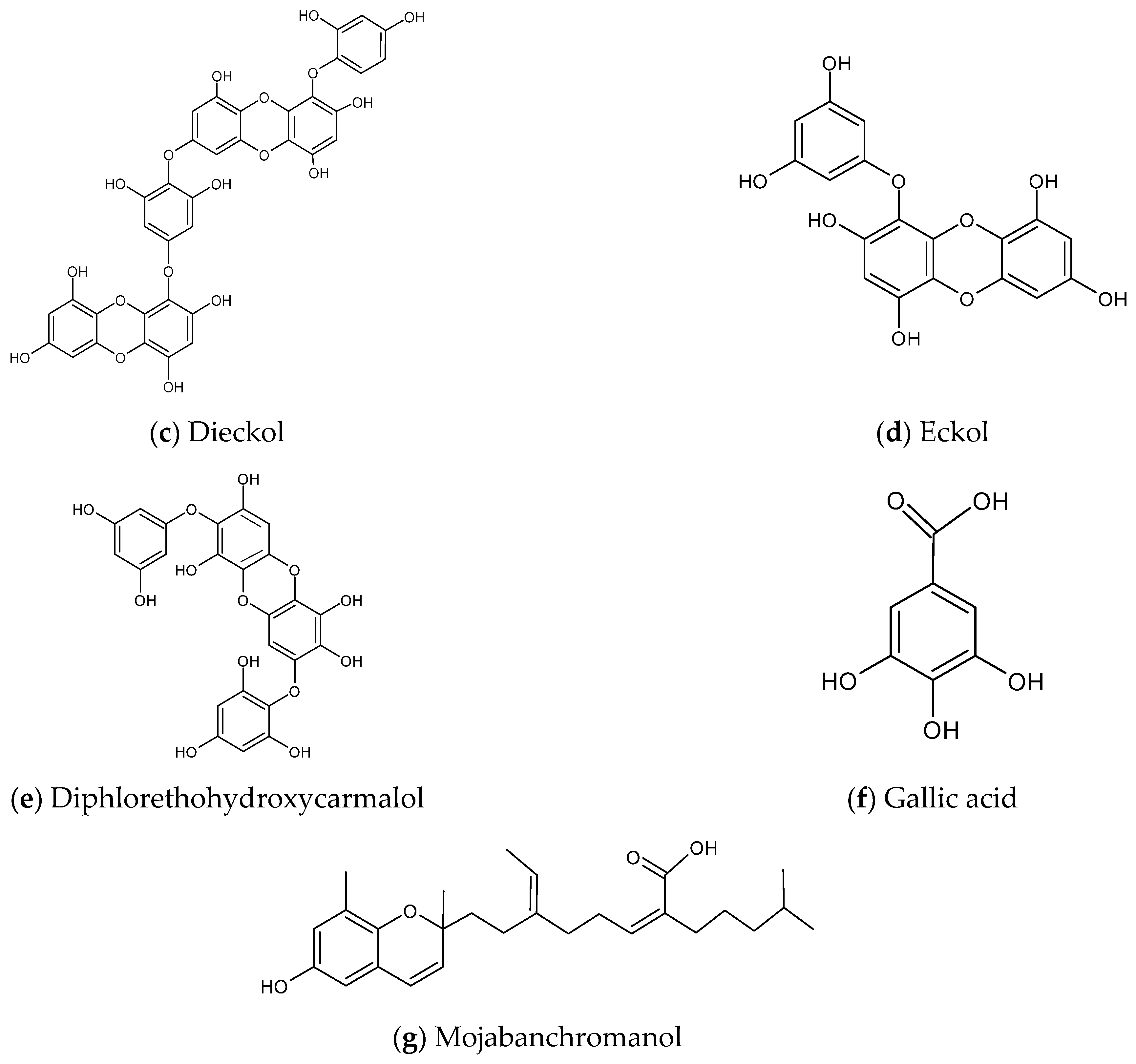
Dieckol and Eckol
Diphlorethohydroxycarmalol (DPHC)
Phenolic Acid
Chromene (Mojabanchromanol)
Phlorotannins and Flavonoid
3.2.4. Ethanol Extract
3.3. Toxicity of Metabolites That Exhibit Anti-Inflammatory Effects
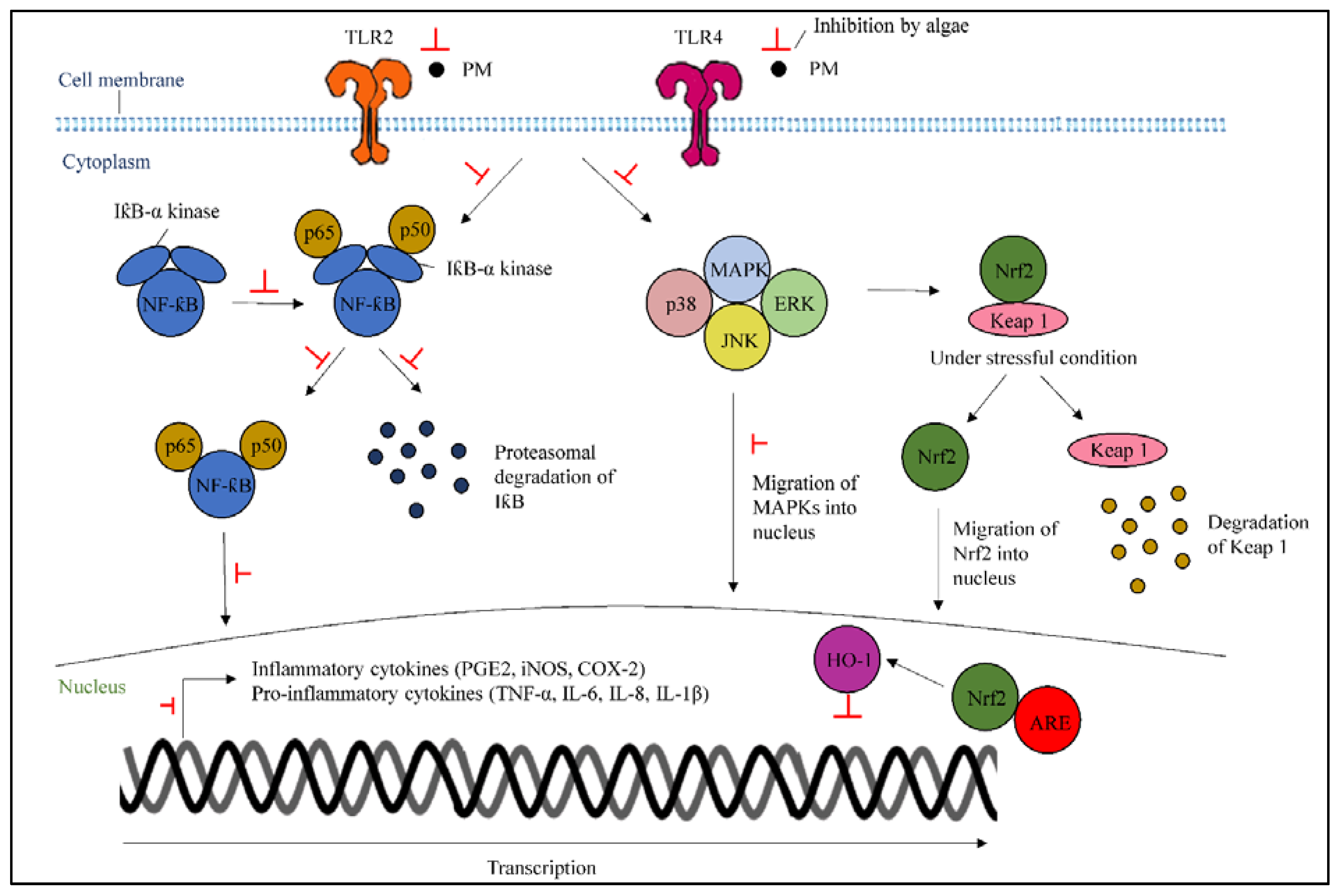
3.4. Mechanism of Actions of Algae to Inhibit Inflammation Caused by Particulate Matters
3.4.1. Inhibition of Mitogen-Activated Protein Kinase and Nuclear Factor-Kappa B
3.4.2. Activation of Nuclear Factor Erythroid 2-Related Factor 2/Heme Oxygenase-1
4. Limitations and Future Directions
4.1. Algae with Anti-Inflammatory Effect
4.2. Experimental Model
5. Materials and Methods
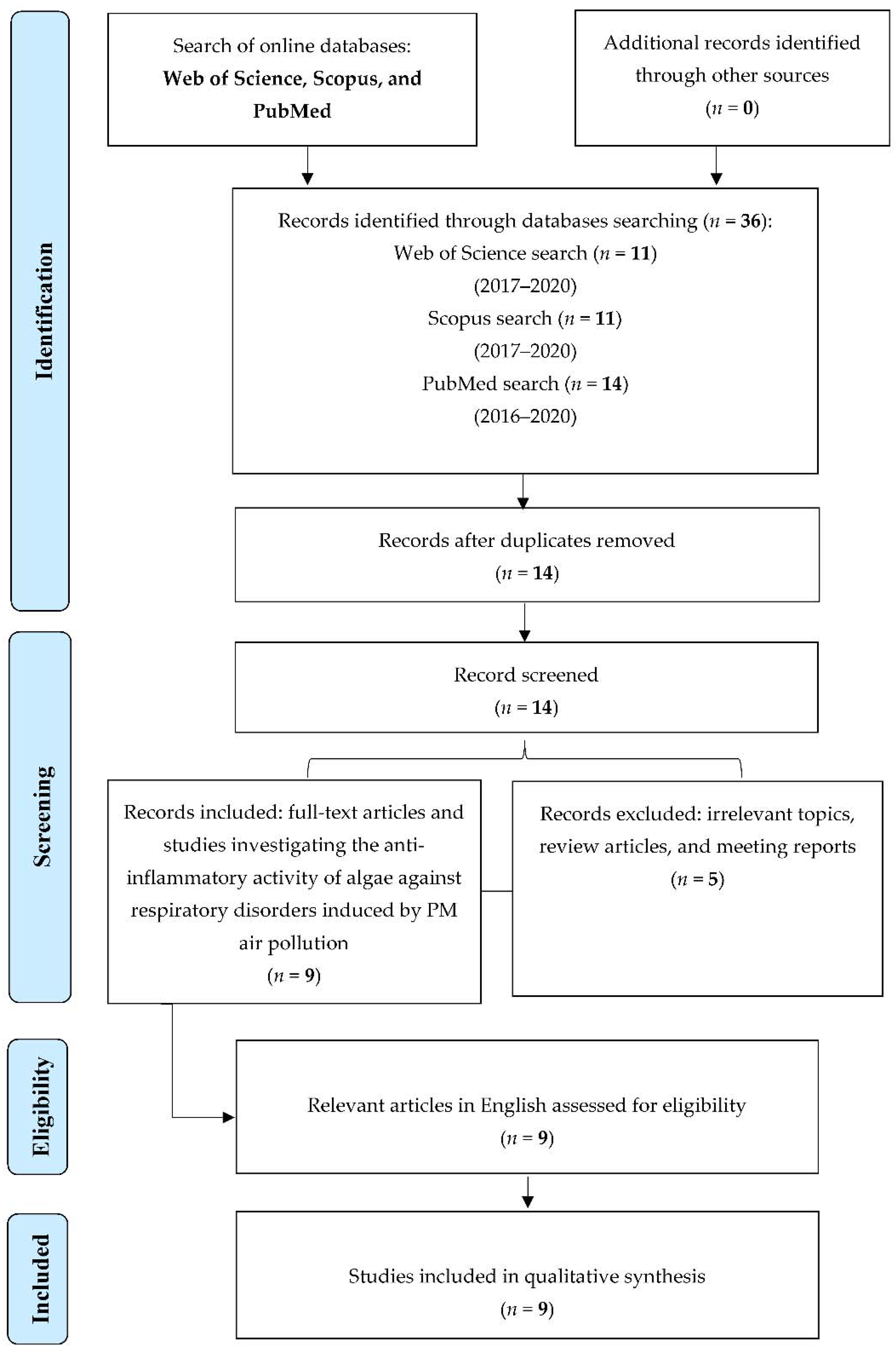
5.1. Source and Search Strategy
5.2. Inclusion and Exclusion Criteria
5.3. Data Extraction
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Li, T.; Hu, R.; Chen, Z.; Li, Q.; Huang, S.; Zhu, Z.; Zhou, L.F. Fine particulate matter (PM2.5): The culprit for chronic lung diseases in China. Chronic Dis. Transl. Med. 2018, 4, 176–186. [Google Scholar]
- Ambient Air Pollution: Health Impacts; World Health Organization (WHO): Geneva, Switzerland, 2018.
- Latif, M.T.; Dominick, D.; Ahamad, F.; Khan, M.F.; Juneng, L.; Hamzah, F.M.; Nadzir, M.S.M. Long term assessment of air quality from a background station on the Malaysian Peninsula. Sci. Total Environ. 2014, 482, 336–348. [Google Scholar] [CrossRef]
- Wu, W.; Jin, Y.; Carlsten, C. Inflammatory health effects of indoor and outdoor particulate matter. J. Allergy Clin. Immunol. 2018, 141, 833–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Chen, Z.; Zhou, L.-F.; Huang, S.-X. Air pollutants and early origins of respiratory diseases. Chronic Dis. Transl. Med. 2018, 4, 75–94. [Google Scholar] [CrossRef]
- Jiang, X.-Q.; Mei, X.-D.; Feng, D. Air pollution and chronic airway diseases: What should people know and do? J. Thorac. Dis. 2016, 8, E31. [Google Scholar] [PubMed]
- Jayawardena, T.U.; Sanjeewa, K.A.; Fernando, I.S.; Ryu, B.M.; Kang, M.C.; Jee, Y.; Lee, W.W.; Jeon, Y.J. Sargassum horneri (Turner) C. Agardh ethanol extract inhibits the fine dust inflammation response via activating Nrf2/HO-1 signaling in RAW 264.7 cells. Bmc Complementary Altern. Med. 2018, 18, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Karakatsani, A.; Analitis, A.; Perifanou, D.; Ayres, J.G.; Harrison, R.M.; Kotronarou, A.; Kavouras, I.G.; Pekkanen, J.; Hämeri, K.; Kos, G.P.; et al. Particulate matter air pollution and respiratory symptoms in individuals having either asthma or chronic obstructive pulmonary disease: A European multicentre panel study. Environ. Health 2012, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Factories 2018, 17, 1–21. [Google Scholar] [CrossRef]
- Cikoš, A.-M.; Jokić, S.; Šubarić, D.; Jerković, I. Overview on the application of modern methods for the extraction of bioactive compounds from marine macroalgae. Mar. Drugs 2018, 16, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sousa, A.A.; Benevides, N.M.; de Freitas Pires, A.; Fiúza, F.P.; Queiroz, M.G.; Morais, T.M.; Pereira, M.G.; Assreuy, A.M. A report of a galactan from marine alga Gelidium crinale with in vivo anti-inflammatory and antinociceptive effects. Fundam. Clin. Pharmacol. 2013, 27, 173–180. [Google Scholar] [CrossRef]
- Priyadarshini, L.; Aggarwal, A. Astaxanthin inhibits cytokines production and inflammatory gene expression by suppressing IκB kinase-dependent nuclear factor κB activation in pre and postpartum Murrah buffaloes during different seasons. Vet. World 2018, 11, 782. [Google Scholar] [CrossRef] [PubMed]
- de Brito, T.V.; Prudêncio, R.D.; Sales, A.B.; Vieira Júnior, F.D.; Candeira, S.J.; Franco, Á.X.; Aragão, K.S.; Ribeiro, R.D.; Ponte de Souza, M.H.; Chaves, L.D.; et al. Anti-inflammatory effect of a sulphated polysaccharide fraction extracted from the red algae H ypnea musciformis via the suppression of neutrophil migration by the nitric oxide signalling pathway. J. Pharm. Pharmacol. 2013, 65, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.A.; de Vanderlei, E.S.; Silva, L.M.; de Araújo, I.W.; de Queiroz, I.N.; de Paula, G.A.; Abreu, T.M.; Ribeiro, N.A.; Bezerra, M.M.; Chaves, H.V.; et al. Antinociceptive and anti-inflammatory activities of a sulfated polysaccharide isolated from the green seaweed Caulerpa cupressoides. Pharmacol. Rep. 2012, 64, 282–292. [Google Scholar] [CrossRef]
- Fernando, I.S.; Jayawardena, T.U.; Kim, H.S.; Lee, W.W.; Vaas, A.P.; De Silva, H.I.; Abayaweera, G.S.; Nanayakkara, C.M.; Abeytunga, D.T.; Lee, D.S.; et al. Beijing urban particulate matter-induced injury and inflammation in human lung epithelial cells and the protective effects of fucosterol from Sargassum binderi (Sonder ex J. Agardh). Environ. Res. 2019, 172, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Fernando, I.S.; Jayawardena, T.U.; Sanjeewa, K.A.; Wang, L.; Jeon, Y.-J.; Lee, W.W. Anti-inflammatory potential of alginic acid from Sargassum horneri against urban aerosol-induced inflammatory responses in keratinocytes and macrophages. Ecotoxicol. Environ. Saf. 2018, 160, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.A.; Fernando, I.P.; Kim, H.S.; Jayawardena, T.U.; Ryu, B.; Yang, H.W.; Ahn, G.; Lee, W.; Jeon, Y.J. Dieckol: An algal polyphenol attenuates urban fine dust-induced inflammation in RAW 264.7 cells via the activation of anti-inflammatory and antioxidant signaling pathways. J. Appl. Phycol. 2020, 32, 2387–2396. [Google Scholar] [CrossRef]
- Fernando, I.; Kim, H.-S.; Sanjeewa, K.; Oh, J.-Y.; Jeon, Y.-J.; Lee, W.W. Inhibition of inflammatory responses elicited by urban fine dust particles in keratinocytes and macrophages by diphlorethohydroxycarmalol isolated from a brown alga Ishige okamurae. Algae 2017, 32, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Herath, K.H.; Kim, H.J.; Lee, J.H.; Je, J.G.; Yu, H.S.; Jeon, Y.J.; Kim, H.J.; Jee, Y. Sargassum horneri (Turner) C. Agardh containing polyphenols attenuates particulate matter-induced inflammatory response by blocking TLR-mediated MYD88-dependent MAPK signaling pathway in MLE-12 cells. J. Ethnopharmacol. 2021, 265, 113340. [Google Scholar] [CrossRef]
- Herath, K.H.; Kim, H.J.; Mihindukulasooriya, S.P.; Kim, A.; Kim, H.J.; Jeon, Y.J.; Jee, Y. Sargassum horneri extract containing mojabanchromanol attenuates the particulate matter exacerbated allergic asthma through reduction of Th2 and Th17 response in mice. Environ. Pollut. 2020, 265, 114094. [Google Scholar] [CrossRef]
- Sanjeewa, K.A.; Jayawardena, T.U.; Lee, H.G.; Herath, K.H.; Jee, Y.; Jeon, Y.J. The protective effect of Sargassum horneri against particulate matter-induced inflammation in lung tissues of an in vivo mouse asthma model. Food Funct. 2019, 10, 7995–8004. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.A.; Jayawardena, T.U.; Kim, S.-Y.; Lee, H.G.; Je, J.-G.; Jee, Y.; Jeon, Y.J. Sargassum horneri (Turner) inhibit urban particulate matter-induced inflammation in MH-S lung macrophages via blocking TLRs mediated NF-κB and MAPK activation. J. Ethnopharmacol. 2020, 249, 112363. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, N.; Alashkar Alhamwe, B.; Caraballo, L.; Ding, M.; Ferrante, A.; Garn, H.; Garssen, J.; Hii, C.S.; Irvine, J.; Llinás-Caballero, K.; et al. Perinatal and Early-Life Nutrition, Epigenetics, and Allergy. Nutrients 2021, 13, 724. [Google Scholar] [CrossRef]
- Acevedo, N.; Frumento, P.; Harb, H.; Alashkar Alhamwe, B.; Johansson, C.; Eick, L.; Alm, J.; Renz, H.; Scheynius, A.; Potaczek, D.P. Histone Acetylation of Immune Regulatory Genes in Human Placenta in Association with Maternal Intake of Olive Oil and Fish Consumption. Int. J. Mol. Sci. 2019, 20, 1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cait, A.; Hughes, M.R.; Antignano, F.; Cait, J.; Dimitriu, P.A.; Maas, K.R.; Reynolds, L.A.; Hacker, L.; Mohr, J.; Finlay, B.B.; et al. Microbiome-driven allergic lung inflammation is ameliorated by short-chain fatty acids. Mucosal Immunol. 2018, 11, 785–795. [Google Scholar] [CrossRef]
- Harb, H.; Irvine, J.; Amarasekera, M.; Hii, C.S.; Kesper, D.A.; Ma, Y.; D’Vaz, N.; Renz, H.; Potaczek, D.P.; Prescott, S.L.; et al. The role of PKCζ in cord blood T-cell maturation towards Th1 cytokine profile and its epigenetic regulation by fish oil. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Wang, M.; Chu, H.; Chu, M.; Na, T.; Wen, Y.; Wu, D.; Han, B.; Bai, Z.; Chen, W.; et al. Global gene expression profiling of human bronchial epithelial cells exposed to airborne fine particulate matter collected from Wuhan, China. Toxicol. Lett. 2014, 228, 25–33. [Google Scholar] [CrossRef]
- Guo, Z.; Hong, Z.; Dong, W.; Deng, C.; Zhao, R.; Xu, J.; Zhuang, G.; Zhang, R. PM2.5-induced oxidative stress and mitochondrial damage in the nasal mucosa of rats. Int. J. Environ. Res. Public Health 2017, 14, 134. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Kim, H.J.; Jee, Y.; Jeon, Y.-J.; Kim, H.J. Antioxidant potential of Sargassum horneri extract against urban particulate matter-induced oxidation. Food Sci. Biotechnol. 2020, 29, 855–865. [Google Scholar] [CrossRef]
- Hou, L.; Zhu, Z.Z.; Zhang, X.; Nordio, F.; Bonzini, M.; Schwartz, J.; Hoxha, M.; Dioni, L.; Marinelli, B.; Pegoraro, V.; et al. Airborne particulate matter and mitochondrial damage: A cross-sectional study. Environ. Health 2010, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakim, M.M.; Patel, I.C. A review on phytoconstituents of marine brown algae. Future J. Pharm. Sci. 2020, 6, 1–11. [Google Scholar]
- Azadmard-Damirchi, S.; Dutta, P.C. Phytosterol classes in olive oils and their analysis by common chromatographic methods. In Olives and Olive oil in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2010; pp. 249–257. [Google Scholar]
- Sánchez-Machado, D.; López-Hernández, J.; Paseiro-Losada, P.; López-Cervantes, J. An HPLC method for the quantification of sterols in edible seaweeds. Biomed. Chromatogr. 2004, 18, 183–190. [Google Scholar] [CrossRef]
- Pozzi, R.; De Berardis, B.; Paoletti, L.; Guastadisegni, C. Inflammatory mediators induced by coarse (PM2.5-10) and fine (PM2.5) urban air particles in RAW 264.7 cells. Toxicology 2003, 183, 243–254. [Google Scholar] [CrossRef]
- Pereira, L. Biological and therapeutic properties of the seaweed polysaccharides. Int. Biol. Rev. 2018, 2, 1–50. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.; Shimizu, M. Natural sleep aids and polyphenols as treatments for insomnia. In Bioactive Nutraceuticals and Dietary Supplements in Neurological and Brain Disease; Elsevier: Amsterdam, The Netherlands, 2015; pp. 141–151. [Google Scholar]
- Zhao, D.; Kwon, S.-H.; Chun, Y.S.; Gu, M.-Y.; Yang, H.O. Anti-neuroinflammatory effects of fucoxanthin via inhibition of Akt/NF-κB and MAPKs/AP-1 pathways and activation of PKA/CREB pathway in lipopolysaccharide-activated BV-2 microglial cells. Neurochem. Res. 2017, 42, 667–677. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Ichinose, T.; Ren, Y.; Song, Y.; Yoshida, Y.; Arashidani, K.; Yoshida, S.; Nishikawa, M.; Takano, H.; Sun, G. PM2.5-rich dust collected from the air in Fukuoka, Kyushu, Japan, can exacerbate murine lung eosinophilia. Inhal. Toxicol. 2015, 27, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liao, J.; Chu, W.; Wang, S.; Yang, T.; Tao, Y.; Wang, G. Involvement of TLR2 and TLR4 and Th1/Th2 shift in inflammatory responses induced by fine ambient particulate matter in mice. Inhal. Toxicol. 2012, 24, 918–927. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Lee, M.S.; Shin, T.S.; Hua, H.; Jang, B.C.; Choi, J.S.; Byun, D.S.; Utsuki, T.; Ingram, D.; Kim, H.R. Phlorofucofuroeckol A inhibits the LPS-stimulated iNOS and COX-2 expressions in macrophages via inhibition of NF-κB, Akt, and p38 MAPK. Toxicol. Vitr. 2011, 25, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Ryu, B.; Choi, I.W.; Qian, Z.J.; Heo, S.J.; Kang, D.H.; Oh, C.; Jeon, Y.J.; Jang, C.H.; Park, W.S.; Kang, K.H.; et al. Anti-inflammatory effect of polyphenol-rich extract from the red alga Callophyllis japonica in lipopolysaccharide-induced RAW 264.7 macrophages. Algae 2014, 29, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Soontornchaiboon, W.; Joo, S.S.; Kim, S.M. Anti-inflammatory effects of violaxanthin isolated from microalga Chlorella ellipsoidea in RAW 264.7 macrophages. Biol. Pharm. Bull. 2012, 35, 1137–1144. [Google Scholar] [CrossRef] [Green Version]
- Suh, S.S.; Hong, J.M.; Kim, E.J.; Jung, S.W.; Kim, S.M.; Kim, J.E.; Kim, I.C.; Kim, S. Anti-inflammation and Anti-Cancer Activity of Ethanol Extract of Antarctic Freshwater Microalga, Micractinium sp. Int. J. Med. Sci. 2018, 15, 929. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algae-Derived Metabolites | Algae Species | Experiment Model | Mechanism of Action | Reference |
|---|---|---|---|---|
Phytosterol
| Sargassum binderi | In vitro (A549 immortalized alveolar basal epithelial) |
| [15] |
Polysaccharides
| Sargassum horneri | In vitro (HaCaT human keratinocyte and RAW264.7 mouse macrophage) |
| [16] |
Polyphenol
| Ecklonia cava | In vitro (RAW264.7 mouse macrophage) |
| [17] |
| Ishige okamurae | In vitro (HaCaT human keratinocyte and RAW264.7 mouse macrophage) and In vivo (zebrafish embryo) |
| [18] |
| Sargassum horneri | In vitro (MLE-12 type II alveolar epithelial cell) |
| [19] |
| Sargassum horneri | In vivo (BALB/c mice) |
| [20] |
| Sargassum horneri | In vivo (BALB/c mice) |
| [21] |
| Sargassum horneri | In vitro (murine MH-S cell) |
| [22] |
| Ethanol extract | Sargassum horneri | In vitro (RAW264.7 mouse macrophage) |
| [7] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, P.X.; Thiyagarasaiyar, K.; Tan, C.-Y.; Jeon, Y.-J.; Nadzir, M.S.M.; Wu, Y.-J.; Low, L.-E.; Atanasov, A.G.; Ming, L.C.; Liew, K.B.; et al. Algae-Derived Anti-Inflammatory Compounds against Particulate Matters-Induced Respiratory Diseases: A Systematic Review. Mar. Drugs 2021, 19, 317. https://doi.org/10.3390/md19060317
Tan PX, Thiyagarasaiyar K, Tan C-Y, Jeon Y-J, Nadzir MSM, Wu Y-J, Low L-E, Atanasov AG, Ming LC, Liew KB, et al. Algae-Derived Anti-Inflammatory Compounds against Particulate Matters-Induced Respiratory Diseases: A Systematic Review. Marine Drugs. 2021; 19(6):317. https://doi.org/10.3390/md19060317
Chicago/Turabian StyleTan, Pek Xyen, Krishnapriya Thiyagarasaiyar, Cheng-Yau Tan, You-Jin Jeon, Mohd Shahrul Mohd Nadzir, Yong-Jiang Wu, Liang-Ee Low, Atanas G. Atanasov, Long Chiau Ming, Kai Bin Liew, and et al. 2021. "Algae-Derived Anti-Inflammatory Compounds against Particulate Matters-Induced Respiratory Diseases: A Systematic Review" Marine Drugs 19, no. 6: 317. https://doi.org/10.3390/md19060317
APA StyleTan, P. X., Thiyagarasaiyar, K., Tan, C. -Y., Jeon, Y. -J., Nadzir, M. S. M., Wu, Y. -J., Low, L. -E., Atanasov, A. G., Ming, L. C., Liew, K. B., Goh, B. -H., & Yow, Y. -Y. (2021). Algae-Derived Anti-Inflammatory Compounds against Particulate Matters-Induced Respiratory Diseases: A Systematic Review. Marine Drugs, 19(6), 317. https://doi.org/10.3390/md19060317