
Sphingolipids of Asteroidea and Holothuroidea: Structures and Biological Activities





Abstract
:1. Introduction
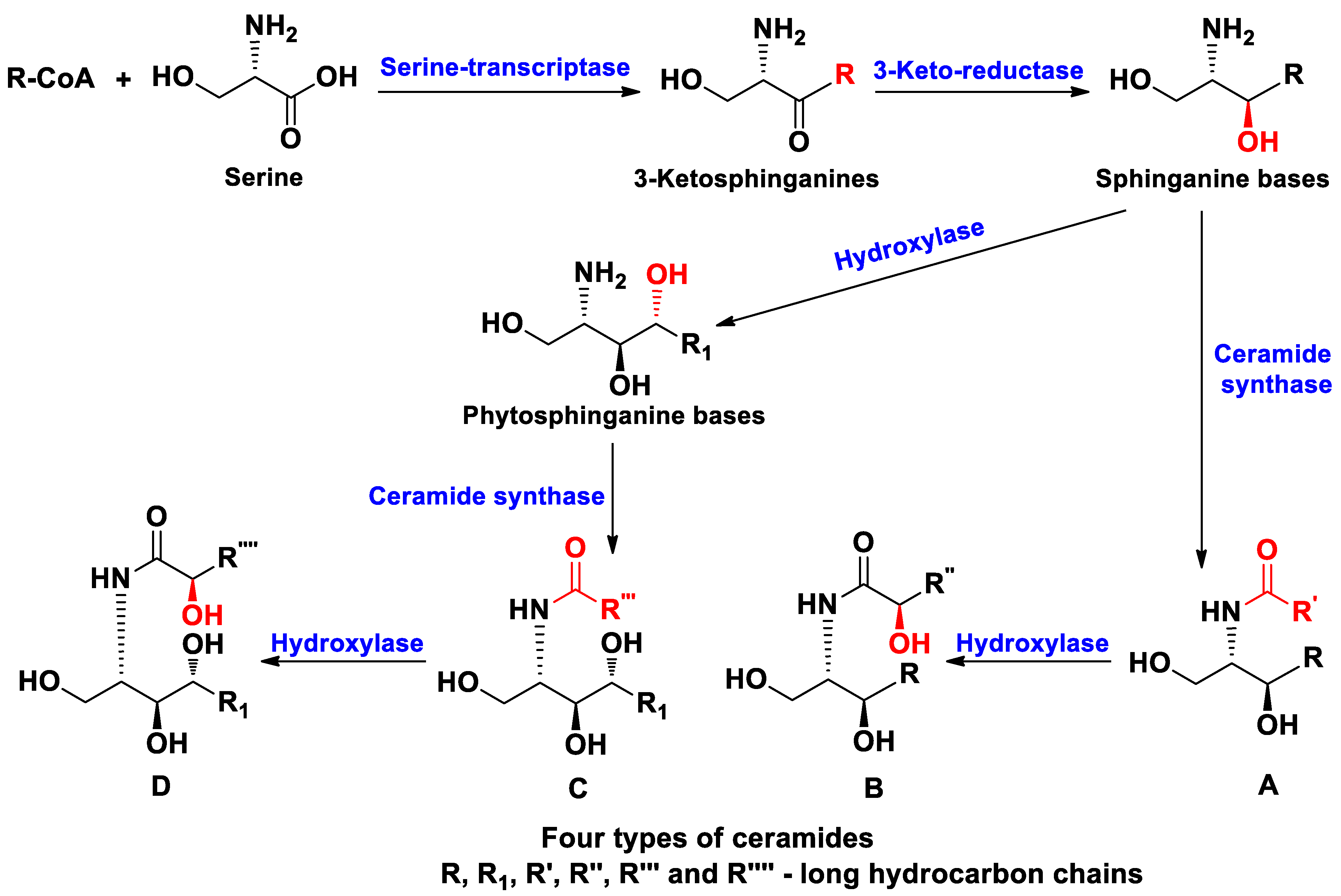
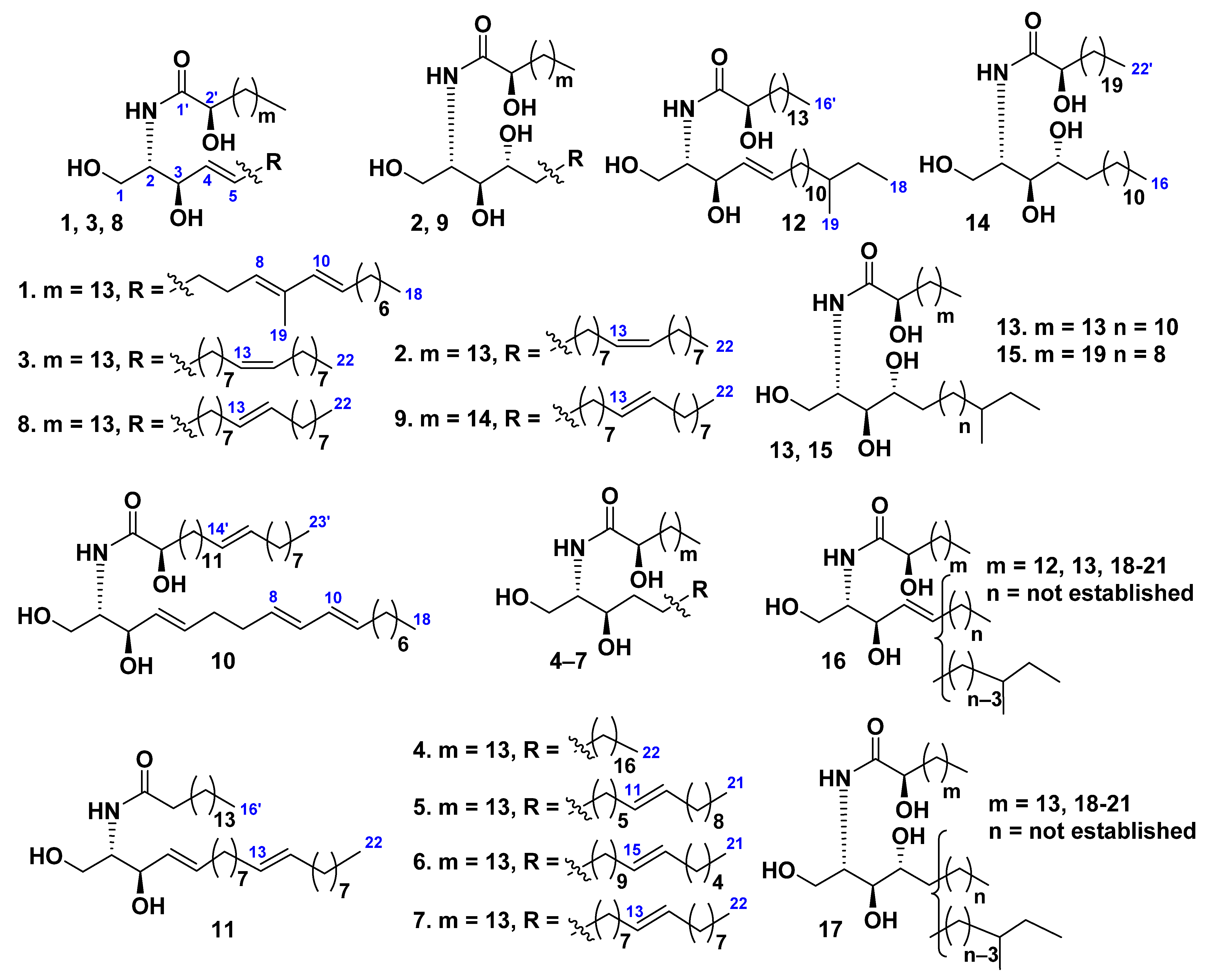
2. Ceramides
Class Asteroidea
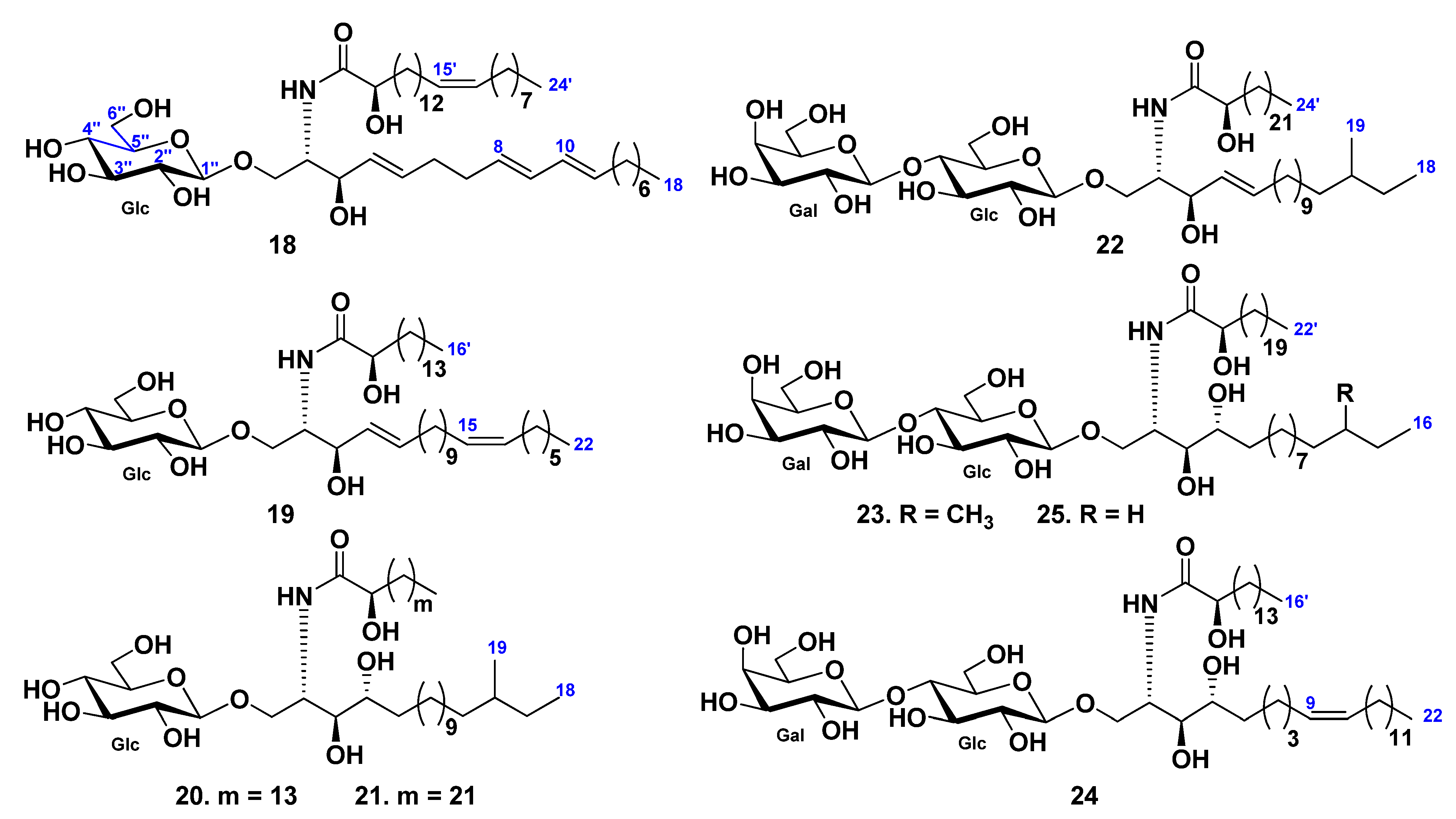
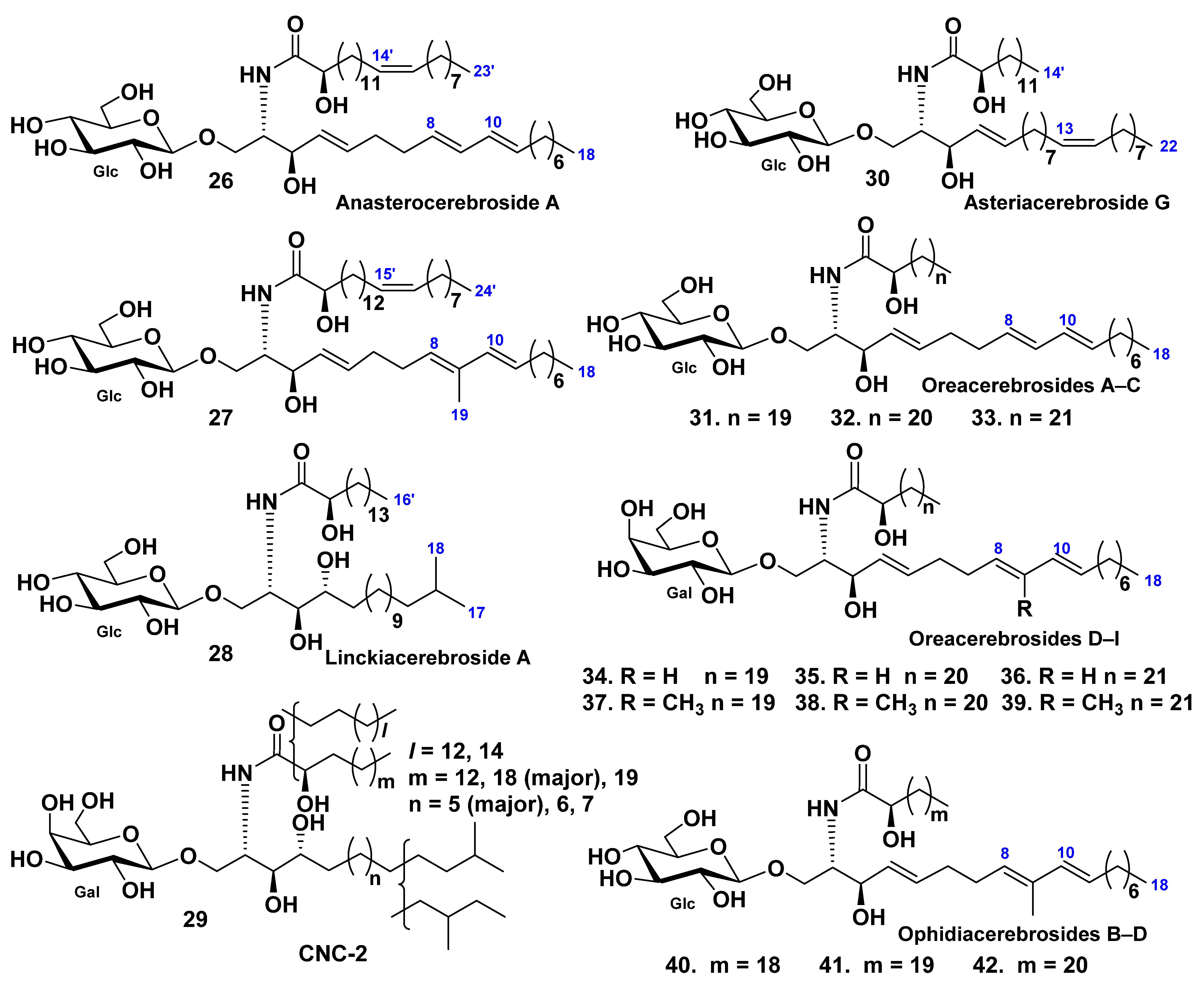
3. Cerebrosides
3.1. Class Asteroidea
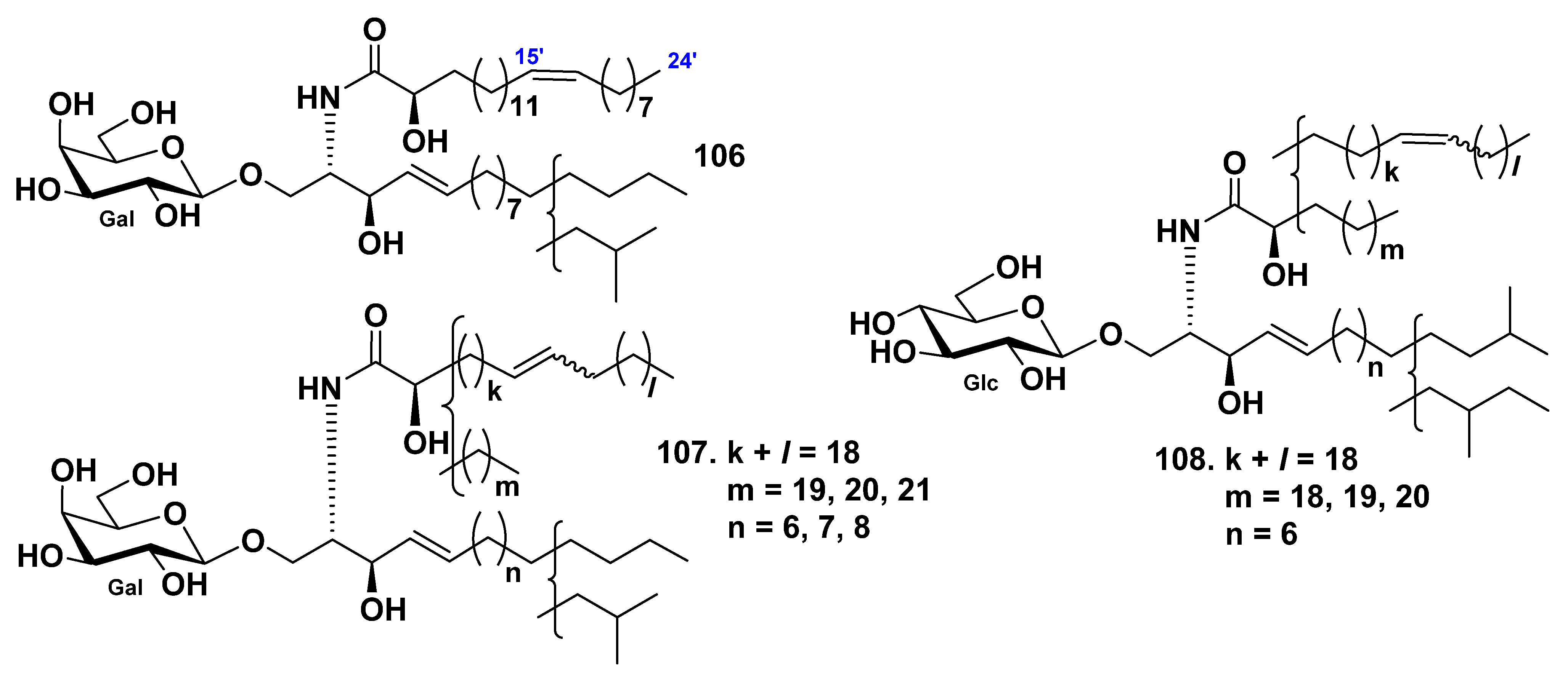
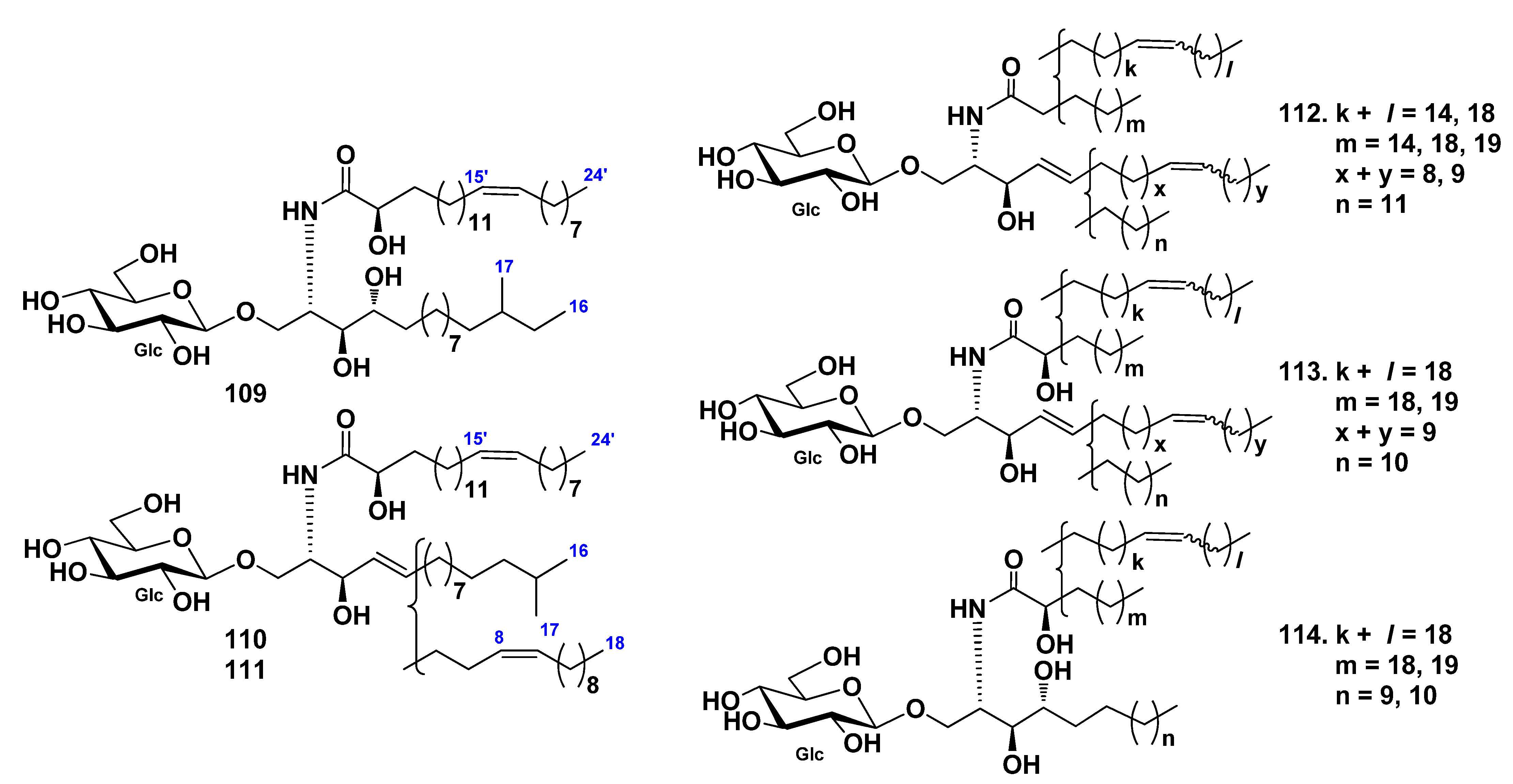
3.2. Class Holothuroidea
4. Gangliosides
4.1. Class Asteroidea
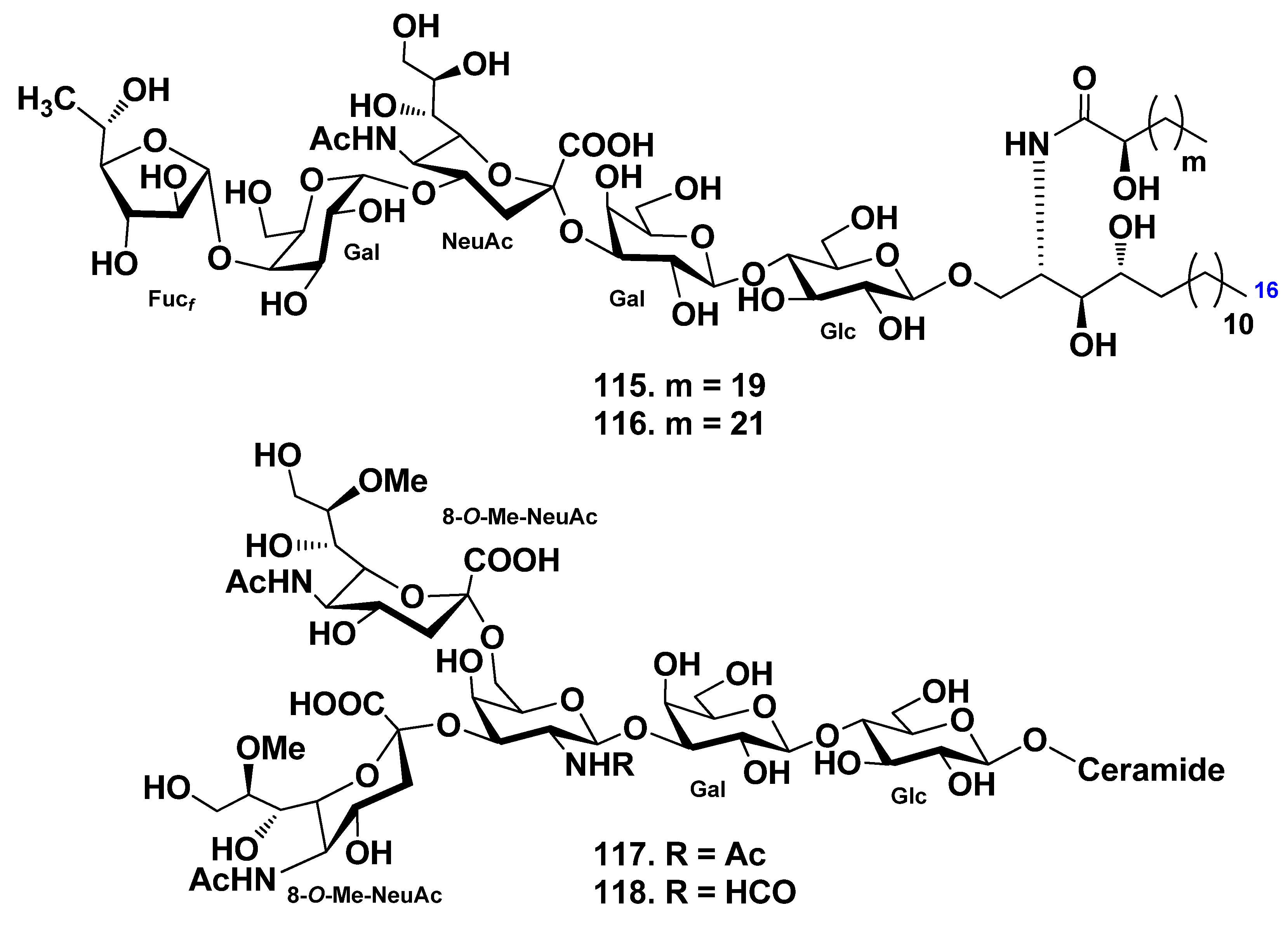
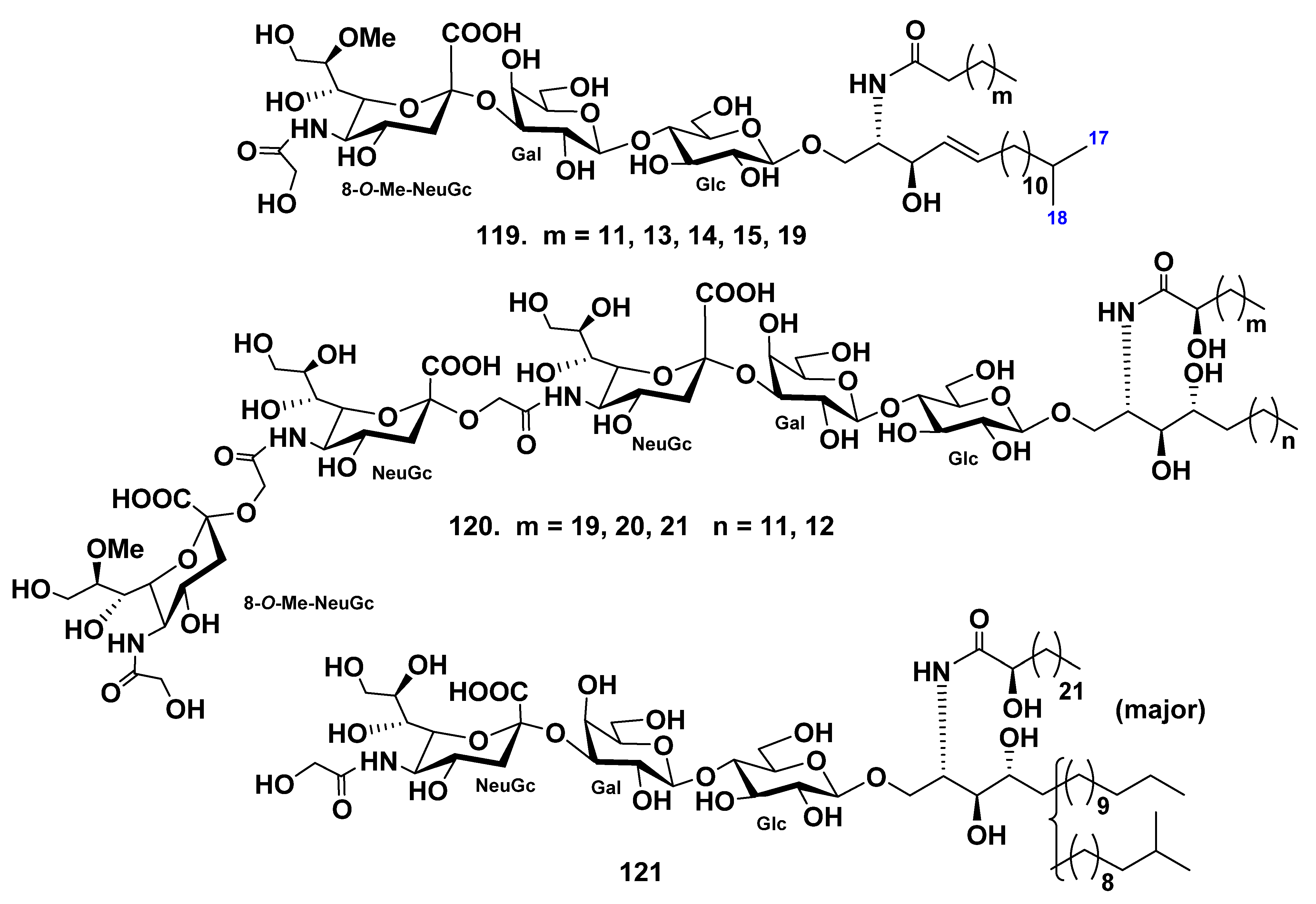
4.2. Class Holothuroidea
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stonik, V.A.; Kalinin, V.I.; Avilov, S.A. Toxins from sea cucumbers (holothuroids): Chemical structures, properties, taxonomic distribution, biosynthesis and evolution. J. Nat. Toxins 1999, 8, 235–248. [Google Scholar] [PubMed]
- Stonik, V.A. Marine polar steroids. Russ. Chem. Rev. 2001, 70, 673–715. [Google Scholar] [CrossRef]
- Stonik, V.A.; Ivanchina, N.V.; Kicha, A.A. New polar steroids from starfish. Nat. Prod. Commun. 2008, 3, 1587–1610. [Google Scholar] [CrossRef] [Green Version]
- Ivanchina, N.V.; Kicha, A.A.; Stonik, V.A. Steroid glycosides from marine organisms. Steroids 2011, 76, 425–454. [Google Scholar] [CrossRef]
- Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Stonik, V.A. Advances in Natural Products Discovery; Gomes, A.R., Rocha-Santos, T., Duarte, A., Eds.; Nova Science Publishers: New York, NY, USA, 2017; Volume 6, pp. 191–224. [Google Scholar]
- Stonik, V.A.; Kicha, A.A.; Malyarenko, T.V.; Ivanchina, N.V. Asterosaponins: Structures, taxonomic distribution, biogenesis and biological activities. Mar. Drugs 2020, 18, 584. [Google Scholar] [CrossRef] [PubMed]
- Fattorusso, E.; Mangoni, A. Marine glycolipids. Prog. Chem. Org. Nat. Prod. 1997, 72, 215–301. [Google Scholar]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Dmitrenok, P.S.; Stonik, V.A. Metabolite profiling of polar steroid constituents in the Far Eastern starfish Aphelasterias japonica using LC–ESI MS/MS. Metabolomics 2014, 10, 1152–1168. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Dmitrenok, P.S.; Stonik, V.A. LC-ESI MS/MS profiling of polar steroid metabolites of the Far Eastern starfish Patiria (=Asterina) pectinifera. Metabolomics 2016, 12, 21. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Grebnev, B.B.; Dmitrenok, P.S.; Stonik, V.A. LC-MS-based metabolome analysis on steroid metabolites from the starfish Patiria (=Asterina) pectinifera in conditions of active feeding and stresses. Metabolomics 2016, 12, 106. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Dmitrenok, P.S. Structural characterization of polar steroid compounds of the Far Eastern starfish Lethasterias fusca by nanoflow liquid chromatography coupled to quadrupole time-of-flight tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 2019, 30, 743–764. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Grebnev, B.B.; Stonik, V.A.; Dmitrenok, P.S. The distribution of asterosaponins, polyhydroxysteroids and related glycosides in different body components of the Far Eastern starfish Lethasterias fusca. Mar. Drugs 2019, 17, 523. [Google Scholar] [CrossRef] [Green Version]
- Popov, R.S.; Ivanchina, N.V.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I.; Dolmatov, I.Y.; Stonik, V.A.; Dmitrenok, P.S. Metabolite profiling of triterpene glycosides of the Far Eastern sea cucumber Eupentacta fraudatrix and their distribution in various body components using LC-ESI QTOF-MS. Mar. Drugs 2017, 15, 302. [Google Scholar] [CrossRef] [Green Version]
- Degroote, S.; Wolthoorn, J.; van Meer, G. The cell biology of glycosphingolipids. Semin. Cell Dev. Biol. 2004, 15, 375–387. [Google Scholar] [CrossRef]
- Hakomori, S.; Igarashi, Y. Functional role of glycosphingolipids in cell recognition and signaling. J. Biochem. 1995, 118, 1091–1103. [Google Scholar] [CrossRef] [PubMed]
- Fahy, E.; Subramaniam, S.; Brown, H.A.; Glass, C.K.; Merrill, A.H., Jr.; Murphy, R.C.; Raetz, C.R.H.; Russell, D.W.; Seyama, Y.; Shaw, W.; et al. A comprehensive classification system for lipids. J. Lipid Res. 2005, 46, 839–861. [Google Scholar] [CrossRef] [Green Version]
- Chebane, K.; Guyot, M. Occurrence of erythro-docosasphinga-4,8-dienine, as an ester, in Anemonia sulcata. Tetrahedron Lett. 1986, 27, 1495–1496. [Google Scholar] [CrossRef]
- Costantino, V.; Fattorusso, E.; Imperatore, C.; Mangoni, A. Glycolipids from sponges. 13.1 Clarhamnoside, the first rhamnosylated α-galactosylceramide from Agelas clathrodes. Improving spectral strategies for glycoconjugate structure determination. J. Org. Chem. 2004, 69, 1174–1179. [Google Scholar] [CrossRef] [PubMed]
- Mansoor, T.A.; Shinde, P.B.; Luo, X.; Hong, J.; Lee, C.O.; Sim, X.; Son, B.W.; Jung, J.H. Renierosides, cerebrosides from a marine sponge Haliclona (Reniera) sp. J. Nat. Prod. 2007, 70, 1481–1486. [Google Scholar] [CrossRef] [PubMed]
- Costantino, V.; Fattorusso, E.; Imperatore, C.; Mangoni, A.; Freigang, S.; Teyton, L. Corrugoside: A new immunostimulatory α-galactoglycosphingolipid from the marine sponge Axinella corrugata. Bioorg. Med. Chem. 2008, 16, 2077–2085. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.Y.; Wen, Z.H.; Chiou, S.F.; Tsai, C.W.; Wang, S.K.; Hsu, C.H.; Dai, C.F.; Chiang, M.Y.; Wang, W.H.; Duh, C.Y. Ceramide and cerebrosides from the octocoral Sarcophyton ehrenbergi. J. Nat. Prod. 2009, 72, 465–468. [Google Scholar] [CrossRef]
- Durán, R.; Zubia, E.; Ortega, M.J.; Naranjo, S.; Salvá, J. Phallusides, new glucosphingolipids from the ascidian Phallusia fumigata. Tetrahedron 1998, 54, 14597–14602. [Google Scholar] [CrossRef]
- Ishii, T.; Okino, T.; Mino, Y. A ceramide and cerebroside from the starfish Asterias amurensis Lütken and their plant-growth promotion activities. J. Nat. Prod. 2006, 69, 1080–1082. [Google Scholar] [CrossRef]
- Duan, J.; Sugawara, T.; Sakai, S.; Aida, K.; Hirata, T. Oral glucosylceramide reduces 2,4-dinitrofluorobenzene induced inflammatory response in mice by reducing TNF-alpha levels and leukocyte infiltration. Lipids 2011, 46, 505–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.; Sugawara, T.; Hirose, M.; Aida, K.; Sakai, S.; Fujii, A.; Hirata, T. Dietary sphingolipids improve skin barrier functions via the upregulation of ceramide synthases in the epidermis. Exp. Dermatol. 2012, 21, 448–452. [Google Scholar] [CrossRef]
- Dillehay, D.L.; Webb, S.K.; Schmelz, E.M.; Merrill, A.H., Jr. Dietary sphingomyelin inhibits 1,2-dimethylhydrazine-induced colon cancer in CF1 mice. J. Nutr. 1994, 124, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. Angiogenesis in health and disease. Nat. Med. 2003, 9, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Muralidhar, P.; Radhika, P.; Krishna, N.; Venkata Rao, D. Bheemasankara Rao, Ch. Sphingolipids from marine organisms: A review. Nat. Prod. Sci. 2003, 9, 117–142. [Google Scholar]
- Higuchi, R.; Inagaki, M.; Yamada, K.; Miyamoto, T. Biologically active gangliosides from echinoderms. J. Nat. Med. 2007, 61, 367–370. [Google Scholar] [CrossRef]
- Yamada, K. Chemo-pharmaceutical studies on the glycosphingolipid constituents from Echinoderm, sea cucumbers, as the medicinal materials. Yakugaku Zasshi 2002, 122, 1133–1143. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, M. Structure and biological activity of glycosphingolipids from starfish and feather star. Yakugaku Zasshi 2008, 128, 1187–1194. [Google Scholar] [CrossRef] [Green Version]
- Sperling, P.; Heinz, E. Plant sphingolipids: Structural diversity, biosynthesis, first genes and functions. Biochem. Biophys. Acta 2003, 1632, 1–15. [Google Scholar] [CrossRef]
- Rho, J.-R.; Kim, Y.H. Isolation and structure determination of three new ceramides from the starfish Distolasterias nipon. Bull. Korean Chem. Soc. 2005, 26, 1457–1460. [Google Scholar] [CrossRef]
- Yoo, J.S.; Park, T.; Bang, G.; Lee, C.; Rho, J.R.; Kim, Y.H. High-energy collision-induced dissociation of [M+Na]+ ions desorbed by fast atom bombardment of ceramides isolated from the starfish Distolasterias nipon. J. Mass Spectrom. 2013, 48, 164–171. [Google Scholar] [CrossRef]
- Inagaki, M.; Ikeda, Y.; Kawatake, S.; Nakamura, K.; Tanaka, M.; Misawa, E.; Yamada, M.; Higuchi, R. Isolation and structure of four new ceramides from the starfish Luidia maculata. Chem. Pharm. Bull. 2006, 54, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Bibel, D.J.; Aly, R.; Shinefield, H.R. Antimicrobial activity of sphingosines. J. Investig. Dermatol. 1992, 98, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.L.; Drake, D.R.; Dawson, D.V.; Blanchette, D.R.; Brogden, K.A.; Wertz, P.W. Antibacterial activity of sphingoid bases and fatty acids against gram-positive and gram-negative bacteria. Antimicrob. Agents Chemother. 2012, 56, 1157–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murshid, S.S.A.; Badr, J.M.; Youssef, D.T.A. Penicillosides A and B: New cerebrosides from the marine-derived fungus Penicillum species. Rev. Bras. Farmacognosia. 2016, 26, 29–33. [Google Scholar] [CrossRef] [Green Version]
- De Vivar, M.E.D.; Seldes, A.M.; Maier, M.S. Two novel glucosylceramides from gonads and body walls of the Patagonian starfish Allostichaster inaequalis. Lipids 2002, 37, 597–603. [Google Scholar] [CrossRef]
- Maier, M.S.; Kuriss, A.; Seldes, A.M. Isolation and structure of glucosylceramides from the starfish Cosmasterias lurida. Lipids 1998, 33, 825–827. [Google Scholar] [CrossRef]
- Kawatake, S.; Nakamura, K.; Inagaki, M.; Higuchi, R. Isolation and structure determination of six glucocerebrosides from the starfish Luidia maculata. Chem. Pharm. Bull. 2002, 50, 1091–1096. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, M.; Nakamura, K.; Kawatake, S.; Higuchi, R. Isolation and structural determination of four new ceramide lactosides from the Starfish Luidia maculata. Eur. J. Org. Chem. 2003, 325–331. [Google Scholar] [CrossRef]
- Chludil, H.D.; Seldes, A.M.; Maier, M.S. Anasterocerebroside A, a new glucosylceramide from the Patagonian starfish Anasterias minuta. Z. Naturforsch. C 2003, 58, 433–440. [Google Scholar] [CrossRef]
- Maruta, T.; Saito, T.; Inagaki, M.; Shibata, O.; Higuchi, R. Biologically active glycosides from Asteroidea, 41. Isolation and structure determination of glucocerebrosides from the starfish Linckia laevigata. Chem. Pharm. Bull. 2005, 53, 1255–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, M.; Nakata, T.; Higuchi, R. Isolation and structure of a galactocerebroside molecular species from the starfish Culcita novaeguineae. Chem. Pharm. Bull. 2006, 54, 260–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costantino, V.; de Rosa, C.; Fattorusso, E.; Imperatore, C.; Mangoni, A.; Irace, C.; Maffettone, C.; Capasso, D.; Malorni, L.; Palumbo, R.; et al. Oreacerebrosides: Bioactive cerebrosides with a triunsaturated sphingoid base from the sea star Oreaster reticulatus. Eur. J. Org. Chem. 2007, 5277–5283. [Google Scholar] [CrossRef]
- Farokhi, F.; Wielgosz-Collin, G.; Clement, M.; Kornprobst, J.-M.; Barnathan, G. Cytotoxicity on human cancer cells of ophidiacerebrosides isolated from the African starfish Narcissia canariensis. Mar. Drugs 2010, 8, 2988–2998. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Rinehart, K.L.; Jares-Erijman, E.A. Ophidiacerebrosides: Cytotoxic glycosphingolipids containing a novel sphingosine from a sea star. J. Org. Chem. 1994, 59, 144–147. [Google Scholar] [CrossRef]
- Pan, K.; Inagaki, M.; Ohno, N.; Tanaka, C.; Higuchi, R.; Miyamoto, T. Identification of sixteen new galactocerebrosides from the starfish Protoreaster nodosus. Chem. Pharm. Bull. 2010, 58, 470–474. [Google Scholar] [CrossRef] [Green Version]
- Park, T.; Park, Y.S.; Rho, J.-R.; Kim, Y.H. Structural determination of cerebrosides isolated from Asterias amurensis starfish eggs using high-energy collision-induced dissociation of sodium-adducted molecules. Rapid Commun. Mass Spectrom. 2011, 25, 572–578. [Google Scholar] [CrossRef]
- Mikami, D.; Sakai, S.; Sasaki, S.; Igarashi, Y. Effects of Asterias amurensis-derived sphingoid bases on the de novo ceramide synthesis in cultured normal human epidermal keratinocytes. J. Oleo Sci. 2016, 65, 671–680. [Google Scholar] [CrossRef] [Green Version]
- Hue, N.; Montagnac, A.; Paїs, M.; Serani, L.; Laprѐvote, O. Structural elucidation of eighteen cerebrosides from Holothuria coronopertusa in a complex mixture by high-energy collision-induced dissociation of [M + Li]+ ions. Eur. J. Mass Spectrom. 2001, 7, 409–417. [Google Scholar] [CrossRef]
- Yamada, K.; Sasaki, K.; Harada, Y.; Isobe, R.; Higuchi, R. Constituents of Holothuroidea. 12. Isolation and structure of glucocerebrosides from the sea cucumber Holothuria pervicax. Chem. Pharm. Bull. 2002, 50, 1467–1470. [Google Scholar] [CrossRef] [Green Version]
- Kisa, F.; Yamada, K.; Kaneko, M.; Inagaki, M.; Higuchi, R. Constituents of Holothuroidea, 14. Isolation and structure of new glucocerebroside molecular species from the sea cucumber Stichopus japonicus. Chem. Pharm. Bull. 2005, 53, 382–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.; Ishida, M.; Aida, K.; Tsuduki, T.; Zhang, J.; Manabe, Y.; Hirata, T.; Sugawara, T. Dietary cerebroside from sea cucumber (Stichopus japonicus): Absorption and effects on skin barrier and caecal short-chain fatty acids. J. Agric. Food Chem. 2016, 64, 7014–7021. [Google Scholar] [CrossRef]
- Yamada, K.; Wada, N.; Onaka, H.; Matsubara, R.; Isobe, R.; Inagaki, M.; Higuchi, R. Constituents of Holothuroidea. 15. Isolation of ante-iso type regioisomer on long chain base moiety of glucocerebroside from the sea cucumber Holothuria leucospilota. Chem. Pharm. Bull. 2005, 53, 788–791. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, T.; Zaima, N.; Yamamoto, A.; Sakai, S.; Noguchi, R.; Hirata, T. Isolation of sphingoid bases of sea cucumber cerebrosides and their cytotoxicity against human colon cancer cells. Biosci. Biotechnol. Biochem. 2006, 70, 2906–2912. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Inagaki, M.; Yamada, K.; Zhang, X.W.; Zhang, B.; Miyamoto, T.; Higuchi, R. Isolation and structure of a galactocerebroside from the sea cucumber Bohadschia argus. Chem. Pharm. Bull. 2009, 57, 315–317. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Wang, Y.M.; Feng, T.Y.; Zhang, B.; Sugawara, T.; Xue, C.H. Isolation and anti-fatty liver activity of a novel cerebroside from the sea cucumber Acaudina molpadioides. Biosci. Biotechnol. Biochem. 2011, 75, 1466–1471. [Google Scholar] [CrossRef]
- Xu, J.; Duan, J.; Xue, C.; Feng, T.; Dong, P.; Sugawara, T.; Hirata, T. Analysis and comparison of glucocerebroside species from three edible sea cucumbers using liquid chromatography–ion trap–time-of-flight mass spectrometry. J. Agric. Food Chem. 2011, 59, 12246–12253. [Google Scholar] [CrossRef]
- Du, L.; Xu, J.; Xue, Y.; Takahashi, K.; Xue, C.H.; Wang, J.F.; Wang, Y.M. Cerebrosides from sea cucumber ameliorates cancer-associated cachexia in mice by attenuating adipose atrophy. J. Funct. Foods 2015, 17, 352–363. [Google Scholar] [CrossRef]
- Liu, X.; Xu, J.; Xue, Y.; Gao, Z.; Li, Z.; Leng, K.; Wang, J.; Xue, C.; Wang, Y. Sea cucumber cerebrosides and long-chain bases from Acaudina molpadioides protect against high fat diet-induced metabolic disorders in mice. Food Funct. 2015, 6, 3428–3536. [Google Scholar] [CrossRef]
- La, M.-P.; Shao, J.-J.; Jiao, J.; Yi, Y.-H. Three cerebrosides from the sea cucumber Cucumaria frondosa. Chin. J. Nat. Med. 2012, 10, 105–109. [Google Scholar] [CrossRef]
- Xu, J.; Guo, S.; Du, L.; Wang, Y.M.; Sugawara, T.; Hirata, T.; Xue, C.H. Isolation of cytotoxic glucocerebrosides and long-chain bases from sea cucumber Cucumaria frondosa using high speed counter-current chromatography. J. Oleo Sci. 2013, 62, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Wang, F.; Wang, J.; Xu, J.; Wang, Y.; Xue, C. The WNT/β-catenin pathway is involved in the anti-adipogenic activity of cerebrosides from the sea cucumber Cucumaria frondosa. Food Funct. 2015, 6, 2396–2404. [Google Scholar] [CrossRef]
- Smirnova, G.P. Gangliosides from the starfish Evasterias echinosoma: Identification of a disialoganglioside containing 8-O-methyl-N-acetylneuraminic acid and N-formylgalactosamine. Russ. Chem. Bull. 2000, 49, 159–164. [Google Scholar] [CrossRef]
- Smirnova, G.P. Structure of gangliosides from gonads of the starfish Evasterias retifera. Russ. Chem. Bull. 2003, 52, 2270–2275. [Google Scholar] [CrossRef]
- Kawatake, S.; Inagaki, M.; Isobe, R.; Miyamoto, T.; Higuchi, R. Isolation and structure of monomethylated GM3-type ganglioside molecular species from the starfish Luidia maculata. Chem. Pharm. Bull. 2002, 50, 1386–1389. [Google Scholar] [CrossRef] [Green Version]
- Kawatake, S.; Inagaki, M.; Isobe, R.; Miyamoto, T.; Higuchi, R. Isolation and structure of a GD3-type ganglioside molecular species possessing neuritogenic activity from the starfish Luidia maculata. Chem. Pharm. Bull. 2004, 52, 1002–1004. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, T.; Yamamoto, A.; Wakabayashi, M.; Nagaregawa, Y.; Inagaki, M.; Higuchi, R.; Iha, M.; Teruya, K. Biologically active glycosides from Asteroidea. 40. Two new gangliosides, acanthagangliosides I and J from the starfish Acanthaster planci. Eur. J. Org. Chem. 2000, 2295–2301. [Google Scholar] [CrossRef]
- Kawano, Y.; Higuchi, R.; Komori, T. Biologically active glycosides from Asteroidea. XIX. Glycosphingolipids from the starfish Acanthaster planci. 4. Isolation and structure of five new gangliosides. Liebigs Ann. Chem. 1990, 43–50. [Google Scholar] [CrossRef]
- Miyamoto, T.; Inagaki, M.; Isobe, R.; Tanaka, Y.; Higuchi, R.; Iha, M.; Teruya, K. Biologically active glycosides from Asteroidea. 36. Re-examination of the structure of acanthaganglioside C, and the identification of three minor acanthagangliosides F, G and H. Liebigs Ann. Chem. 1997, 931–936. [Google Scholar] [CrossRef]
- Hanashima, S.; Sato, K.I.; Naito, Y.; Takematsu, H.; Kozutsumi, Y.; Ito, Y.; Yamaguchi, Y. Synthesis and binding analysis of unique AG2 pentasaccharide to human Siglec-2 using NMR techniques. Bioorg. Med. Chem. 2010, 18, 3720–3725. [Google Scholar] [CrossRef]
- Higuchi, R.; Inoue, S.; Inagaki, K.; Sakai, M.; Miyamoto, T.; Komori, T.; Inagaki, M.; Isobe, R. Biologically active glycosides from Asteroidea. 42. Isolation and structure of a new biologically active ganglioside molecular species from the starfish Asterina pectinifera. Chem. Pharm. Bull. 2006, 54, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Smirnova, G.P. Hematoside with 8-O-methyi-N-glycolylneuraminic acid from the starfish Linckia laevigata. Russ. Chem. Bull. 2000, 49, 165–168. [Google Scholar] [CrossRef]
- Inagaki, M.; Miyamoto, T.; Isobe, R.; Higuchi, R. Biologically active glycosides from Asteroidea. 43. Isolation and structure of a new neuritogenic-active ganglioside molecular species from the starfish Linckia laevigata. Chem. Pharm. Bull. 2005, 53, 1551–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, M.; Saito, T.; Miyamoto, T.; Higuchi, R. Isolation and structure of hematoside-type ganglioside from the starfish Linckia laevigata. Chem. Pharm. Bull. 2009, 57, 204–206. [Google Scholar] [CrossRef] [Green Version]
- Pan, K.; Tanaka, C.; Inagaki, M.; Higuchi, R.; Miyamoto, T. Isolation and structure elucidation of GM4-type gangliosides from the Okinawan starfish Protoreaster nodosus. Mar. Drugs 2012, 10, 2467–2480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, K.; Harada, Y.; Miyamoto, T.; Isobe, R.; Higuchi, R. Constituents of Holothuroidea. 9. Isolation and structure of a new ganglioside molecular species from the sea cucumber Holothuria pervicax. Chem. Pharm. Bull. 2000, 48, 157–159. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.; Harada, Y.; Nagaregawa, Y.; Miyamoto, T.; Isobe, R.; Higuchi, R. Constituents of Holothuroidea. 7. Isolation and structure of biologically active gangliosides from the sea cucumber Holothuria pervicax. Eur. J. Org. Chem. 1998, 2519–2525. [Google Scholar] [CrossRef]
- Yamada, K.; Matsubara, R.; Kaneko, M.; Miyamoto, T.; Higuchi, R. Constituents of Holothuroidea. 10. Isolation and structure of a biologically active ganglioside molecular species from the sea cucumber Holothuria leucospilota. Chem. Pharm. Bull. 2001, 49, 447–452. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, M.; Kisa, F.; Yamada, K.; Miyamoto, T.; Higuchi, R. Structure of a new neuritogenic-active ganglioside from the sea cucumber Stichopus japonicus. Eur. J. Org. Chem. 2003, 1004–1008. [Google Scholar] [CrossRef]
- Kaneko, M.; Kisa, F.; Yamada, K.; Miyamoto, T.; Higuchi, R. Constituents of Holothuroidea. 8-Structure of neuritogenic active ganglioside from the sea cucumber Stichopus japonicus. Eur. J. Org. Chem. 1999, 3171–3174. [Google Scholar] [CrossRef]
- Yamada, K.; Hamada, A.; Kisa, F.; Miyamoto, T.; Higuchi, R. Constituents of Holothuroidea. 13. Structure of neuritogenic active ganglioside molecular species from the sea cucumber Stichopus chloronotus. Chem. Pharm. Bull. 2003, 51, 46–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisa, F.; Yamada, K.; Miyamoto, T.; Inagaki, M.; Higuchi, R. Constituents of Holothuroidea. 17. Isolation and structure of biologically active monosialo-gangliosides from the sea cucumber Cucumaria echinata. Chem. Pharm. Bull. 2006, 54, 982–987. [Google Scholar] [CrossRef] [Green Version]
- Kisa, F.; Yamada, K.; Miyamoto, T.; Inagaki, M.; Higuchi, R. Constituents of Holothuroidea. 18. Isolation and structure of biologically active disialo- and trisialo-gangliosides from the sea cucumber Cucumaria echinata. Chem. Pharm. Bull. 2006, 54, 1293–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traving, C.; Schauer, R. Structure, function and metabolism of sialic acids. Cell. Mol. Life Sci. 1998, 54, 1330–1349. [Google Scholar] [CrossRef]
- Kolter, T. Ganglioside biochemistry. ISRN Biochem. 2012, 2012, 506160. [Google Scholar] [CrossRef] [Green Version]
- Sipione, S.; Monyror, J.; Galleguillos, D.; Steinberg, N.; Kadam, V. Gangliosides in the brain: Physiology, pathophysiology and therapeutic applications. Front. Neurosci. 2020, 14, 572965. [Google Scholar] [CrossRef]
- Schengrund, C.-L. Gangliosides and neuroblastomas. Int. J. Mol. Sci. 2020, 21, 5313. [Google Scholar] [CrossRef] [PubMed]
- Schömel, N.; Geisslingera, G.; Wegnera, M.-S. Influence of glycosphingolipids on cancer cell energy metabolism. Prog. Lipid Res. 2020, 79, 101050. [Google Scholar] [CrossRef]
- Yamada, K.; Hara, E.; Miyamoto, T.; Higuchi, R.; Isobe, R.; Honda, S. Constituents of Holothuroidea, 6-Isolation and structure of biologically active glycosphingolipids from the sea cucumber Cucumaria echinata. Eur. J. Org. Chem. 1998, 371–378. [Google Scholar] [CrossRef]
- Kisa, F.; Yamada, K.; Miyamoto, T.; Inagaki, M.; Higuchi, R. Determination of the absolute configuration of sialic acids in gangliosides from the sea cucumber Cucumaria echinata. Chem. Pharm. Bull. 2007, 55, 1051–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kicha, A.A.; Kalinovsky, A.I.; Malyarenko, T.V.; Ivanchina, N.V.; Dmitrenok, P.S.; Menchinskaya, E.S.; Yurchenko, E.A.; Pislyagin, E.A.; Aminin, D.L.; Huong, T.T.; et al. Cyclic steroid glycosides from the starfish Echinaster luzonicus: Structures and immunomodulatory activities. J. Nat. Prod. 2015, 78, 1397–1405. [Google Scholar] [CrossRef]
- Malyarenko, T.V.; Kharchenko, S.D.; Kicha, A.A.; Ivanchina, N.V.; Dmitrenok, P.S.; Chingizova, E.A.; Pislyagin, E.A.; Evtushenko, E.V.; Antokhina, T.I.; Minh, C.V.; et al. Anthenosides L‒U, steroidal glycosides with unusual structural features from the starfish Anthenea aspera. J. Nat. Prod. 2016, 79, 3047–3056. [Google Scholar] [CrossRef]
- Mondol, M.A.M.; Shin, H.J.; Rahman, M.A.; Islam, M.T. Sea cucumber glycosides: Chemical structures, producing species and important biological properties. Mar. Drugs 2017, 15, 317. [Google Scholar] [CrossRef] [Green Version]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
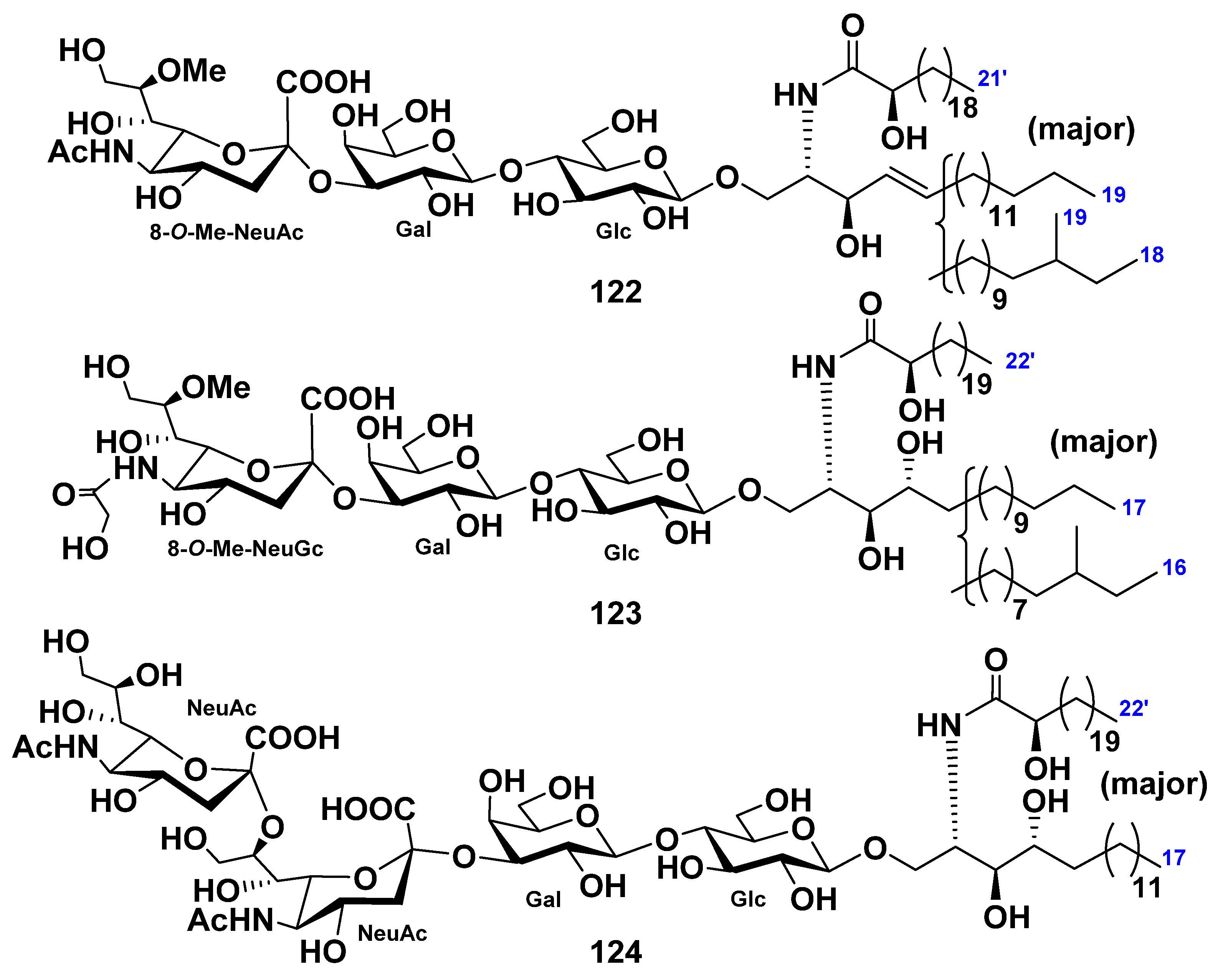{kind=link}
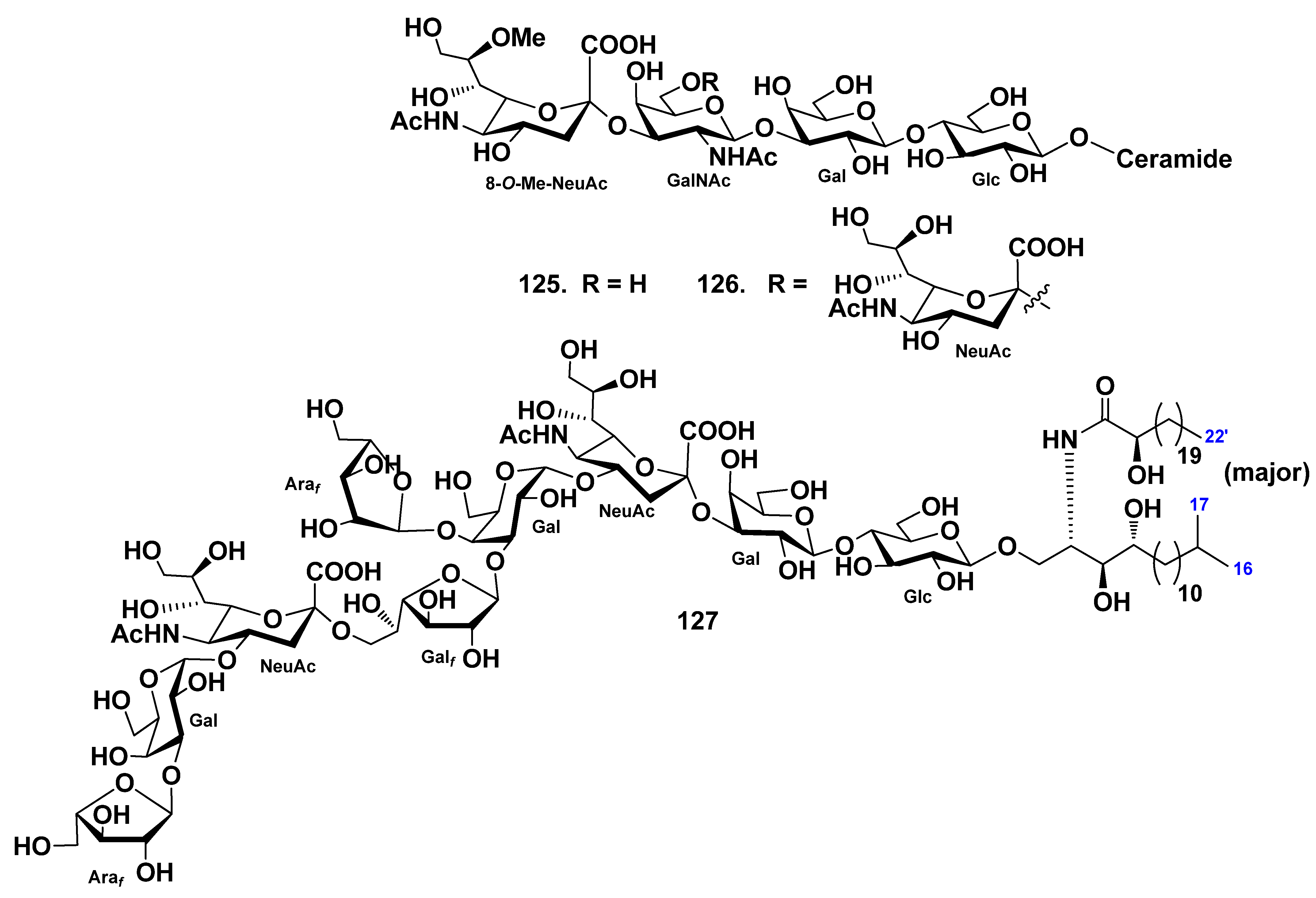{kind=link}
{kind=link}
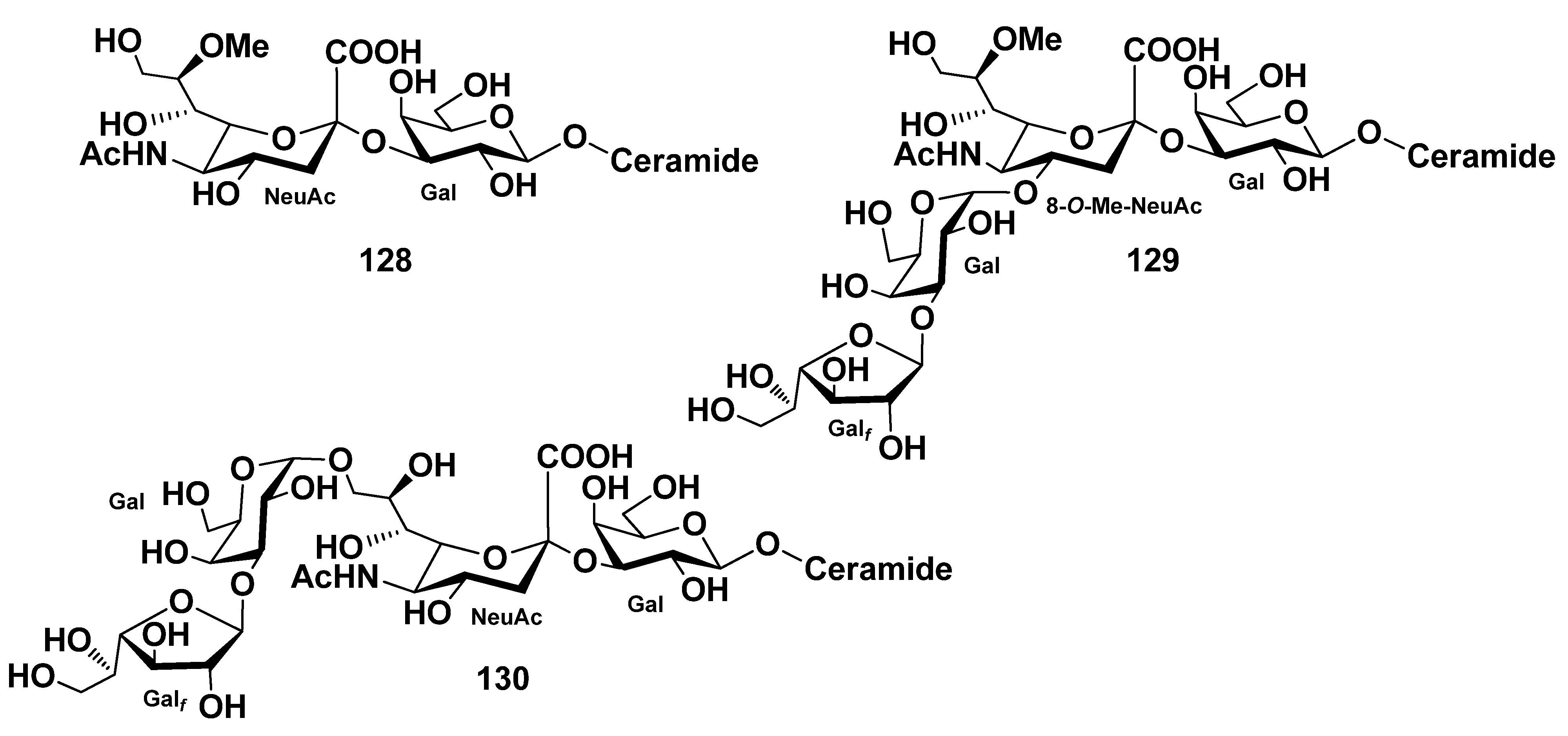
{kind=link}
{kind=link}
{kind=link}
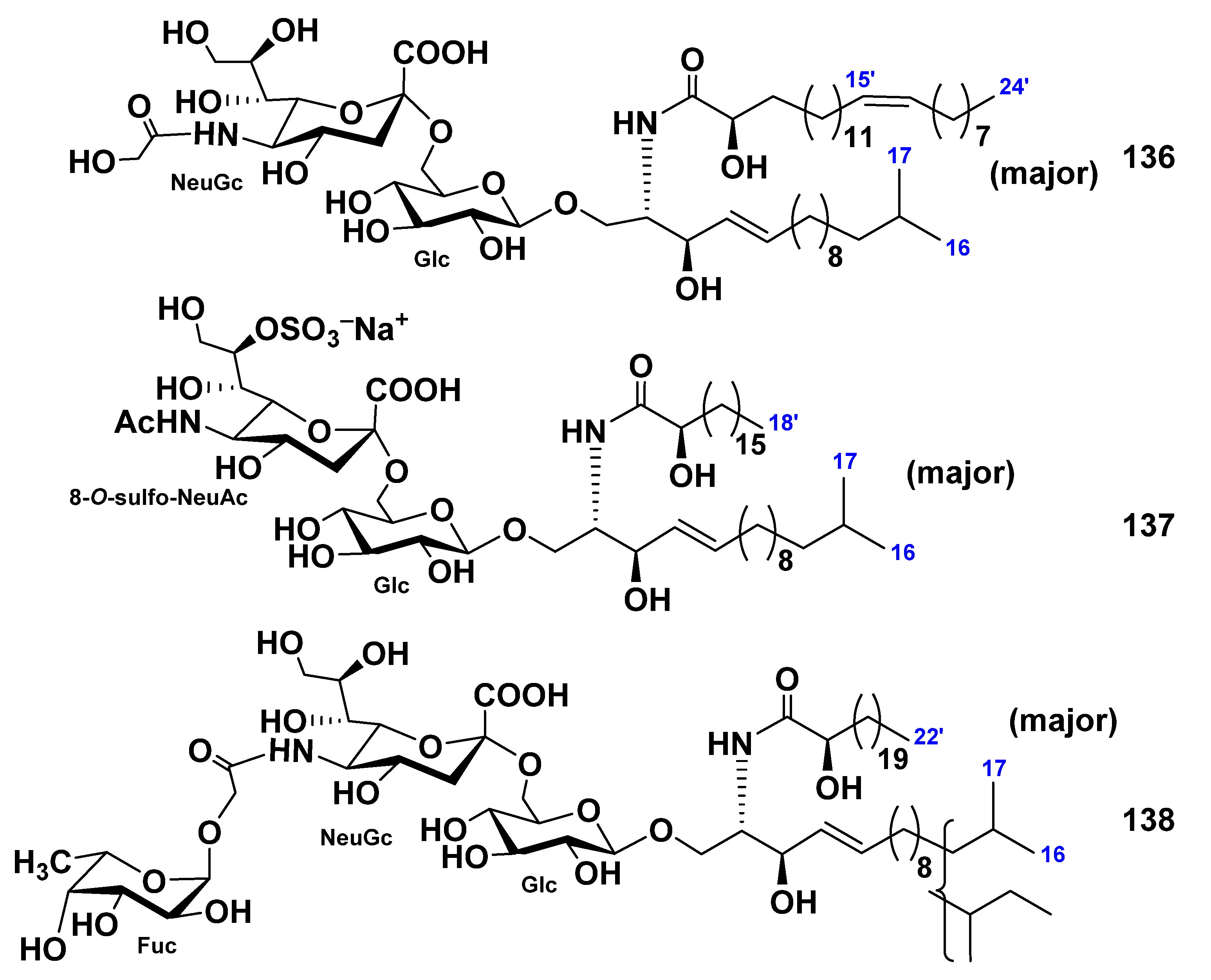{kind=link}
{kind=link}
| Order | Family | Scientific Name | Compounds | Type of Biological Activity | Ref. | ||
|---|---|---|---|---|---|---|---|
| Ceramides | Cerebrosides | Gangliosides | |||||
| Class Asteroidea | |||||||
| Forcipulatida | Asteriidae | Allostichaster inaequalis | 18, 19 | [39] | |||
| Anasterias minuta | 26, 27 | [43] | |||||
| Asterias amurensis | 2 | 30, 59–64 | Stimulating root growth Brassica campestris (2, 30); | [23,50] | |||
| Distolasterias nipon | 1–11 | 65–68 | [33,34] | ||||
| Evasterias echinosoma | 117, 118 | [66] | |||||
| Evasterias retifera | 125, 126 | [67] | |||||
| Cosmasterias lurida | 27 | [40] | |||||
| Paxillosida | Luidiidae | Luidia maculata | 12–17 | 20–25 | 122–124 | Neuritogenic activity toward the rat pheochromocytoma PC12 cells in the presence of NGF (124) | [35,41,42,68,69] |
| Valvatida | Acanthaster-idae | Acanthaster planci | 115, 116 | Binding epitope of AG2 pentasaccharide to human Siglec-2 | [70,71,72,73] | ||
| Asterinidae | Patiria (=Asterina) pectinifera | 127 | Neuritogenic activity toward the rat pheochromocytoma PC12 cells in the presence of NGF | [74] | |||
| Ophidiasteridae | Linckia laevigata | 28 | 119–121 | Neuritogenic activity toward the rat pheochromocytoma PC12 cells in the presence of NGF (120) | [44,75,76,77] | ||
| Oreasteridae | Culcita novaeguineae | 29 | [45] | ||||
| Protoreaster nodosus | 43–58 | 128–130 | [49,78] | ||||
| Oreaster reticulatus | 31–39 | (1) Mildly cytotoxic activity on the rat glioma C6 cells (31–39); (2) exertion of proangiogenic activity and increase of VEGF-induced human endothelial cell proliferation (39). | [46] | ||||
| Narcissia canariensis | 40–42 | Cytotoxic activity against KB cells (40) | [47] | ||||
| Class Holothuroidea | |||||||
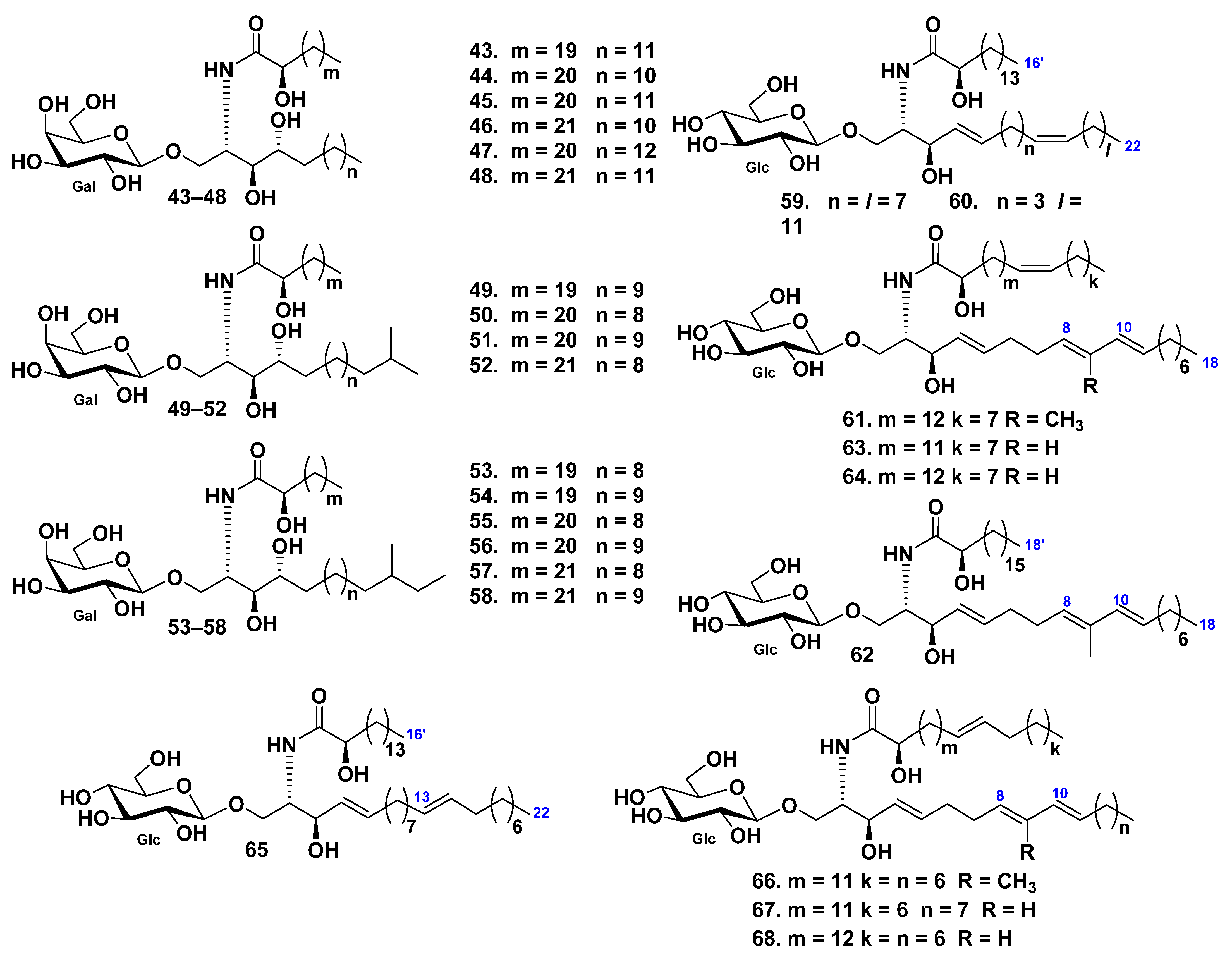
| Holothuriida | Holothuri-idae | Holothuria coronopertusa | 69–86 | [52] | |||
| Holothuria pervicax | 87–96 | 131 | Neuritogenic activity toward the rat pheochromocytoma PC12 cell line (131) | [53,79,80] | |||
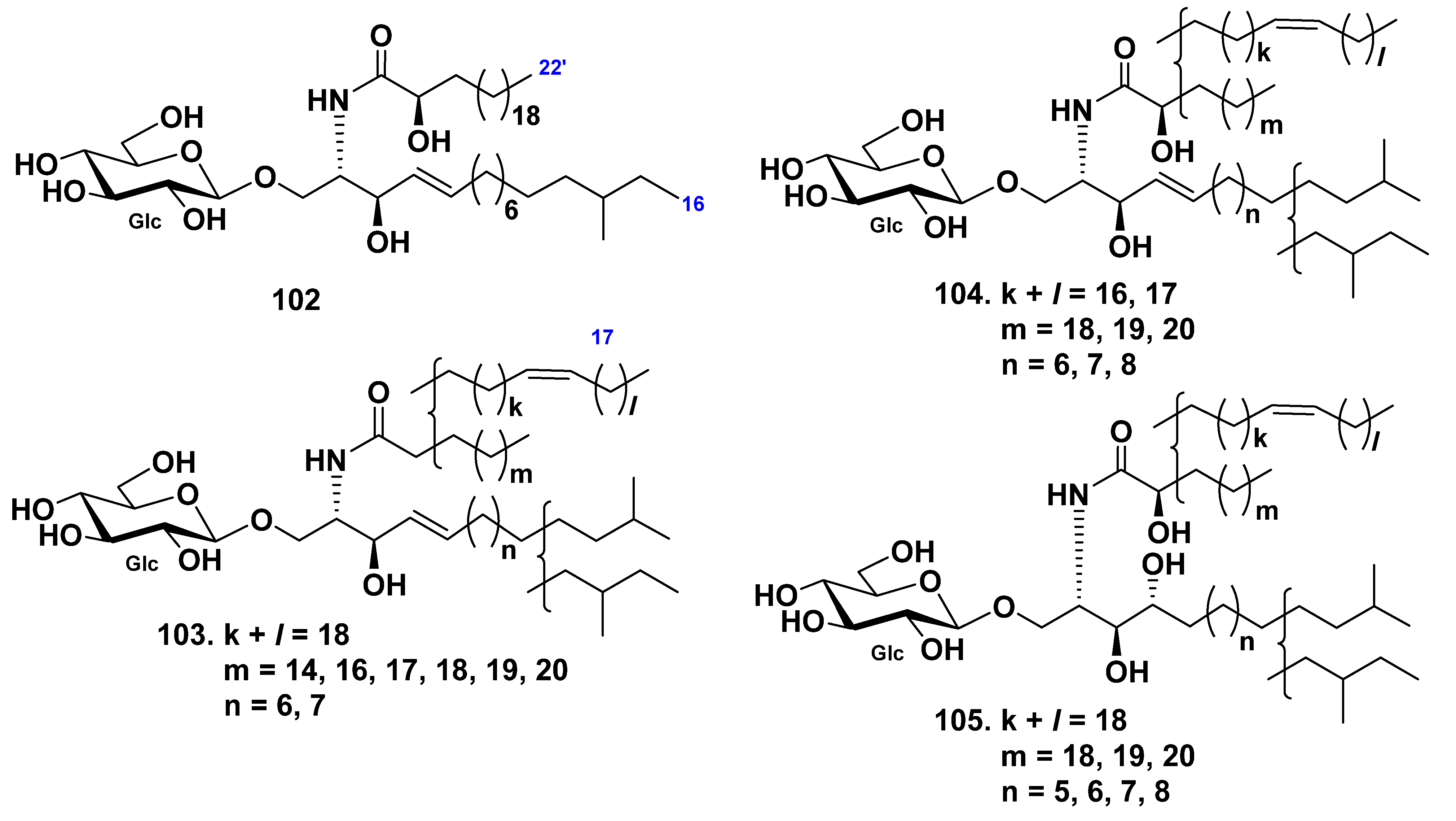
| Holothuria leucospilota | 102–105 | 132–134 | Neuritogenic activity toward the rat pheochromocytoma PC12 cell line (132–134) | [56,81] | |||
| Bohadschia argus | 106, 107 | [58] | |||||
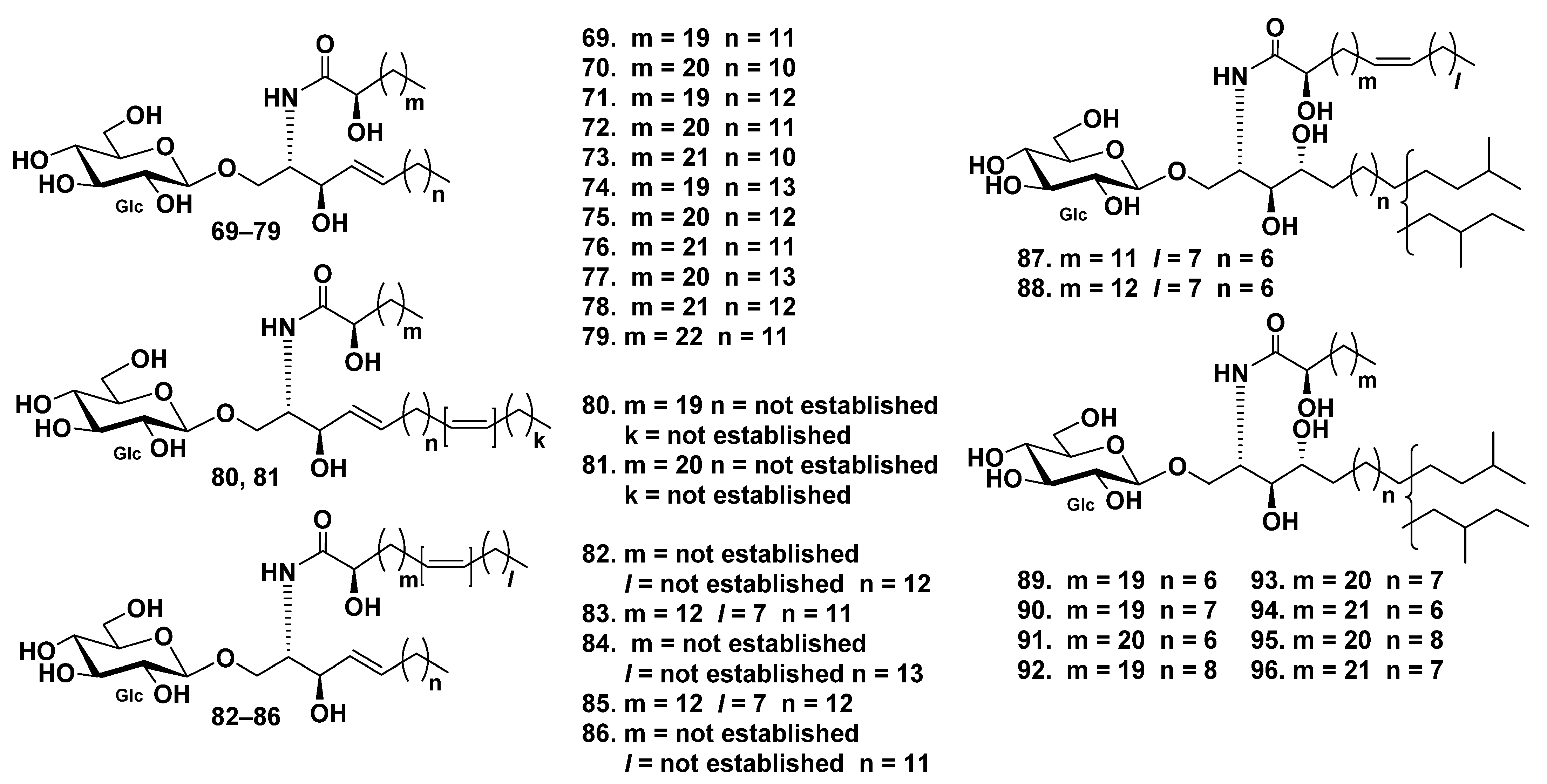
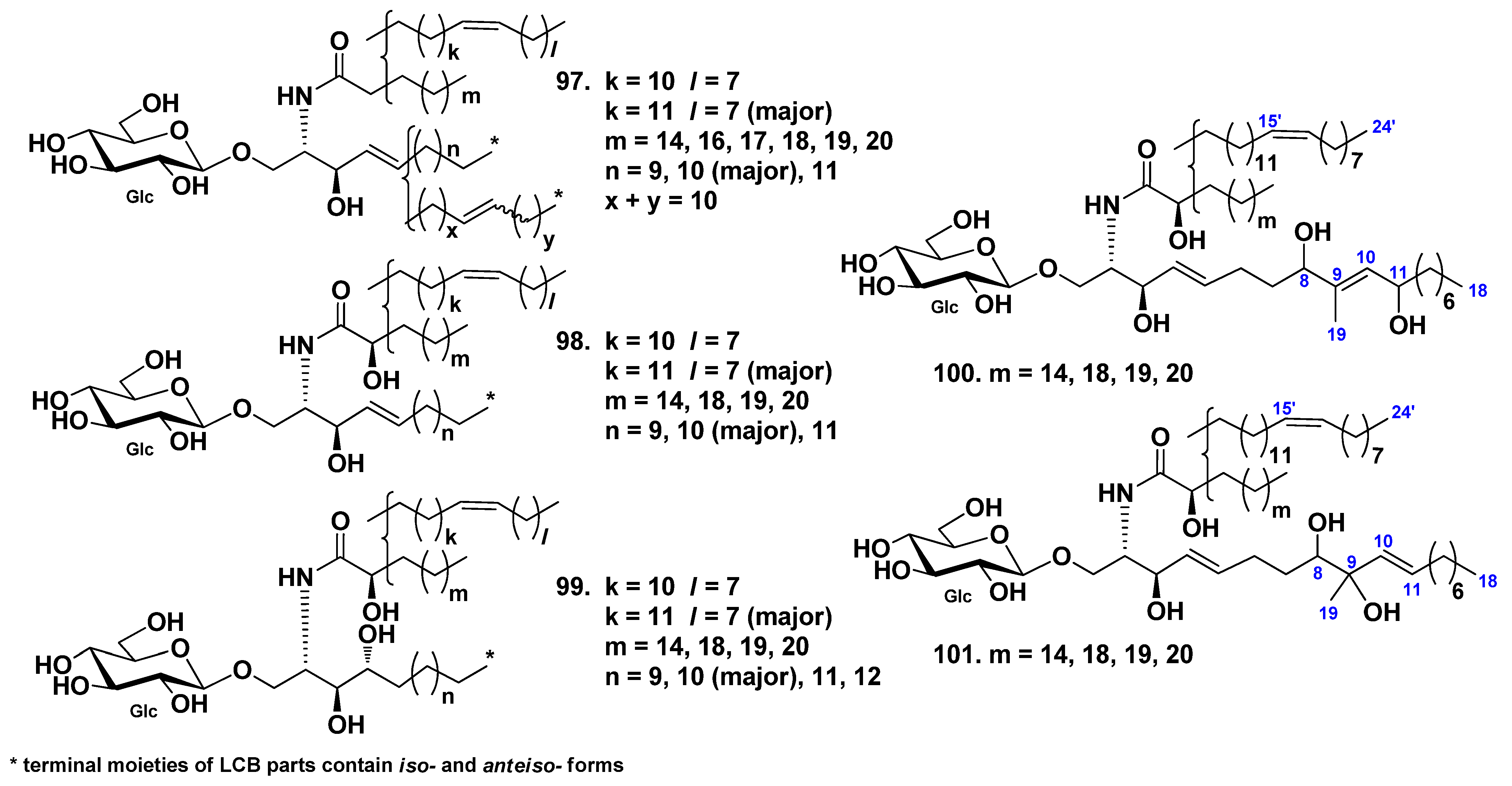
| Synallactida | Stichopodi-dae | Stichopus japonicus | 97–101 | 135 | (1) Absorption of cerebrosides in vivo and improving skin barrier functions (97–101); (2) neuritogenic activity toward the rat pheochromocytoma PC12 cells in the presence of NGF (135). | [54,55,82,83] | |
| Stichopus chloronotus | 136–138 | Neuritogenic activity toward the rat pheochromocytoma PC12 cells in the presence of NGF | [84] | ||||
| Molpadida | Caudinidae | Acaudina molpadioides | 108 | (1) Anti-fatty liver activity of 108 in the rats with fatty liver induced by orotic acid; (2) alleviating adipose atrophy in the cancer-associated cachexia mice; (3) effects of cerebrosides on the obesity-related metabolic disorders in mice. | [59,61,62] | ||
| Dendrochi-rotida | Cucu-mariidae | Cucumaria frondosa | 109–114 | (1) In vitro cytotoxic activity against Caco-2 colon cancer cells (112–114); (2) in vitro antiadipogenic activity of cerebrosides. | [63,64,65] | ||
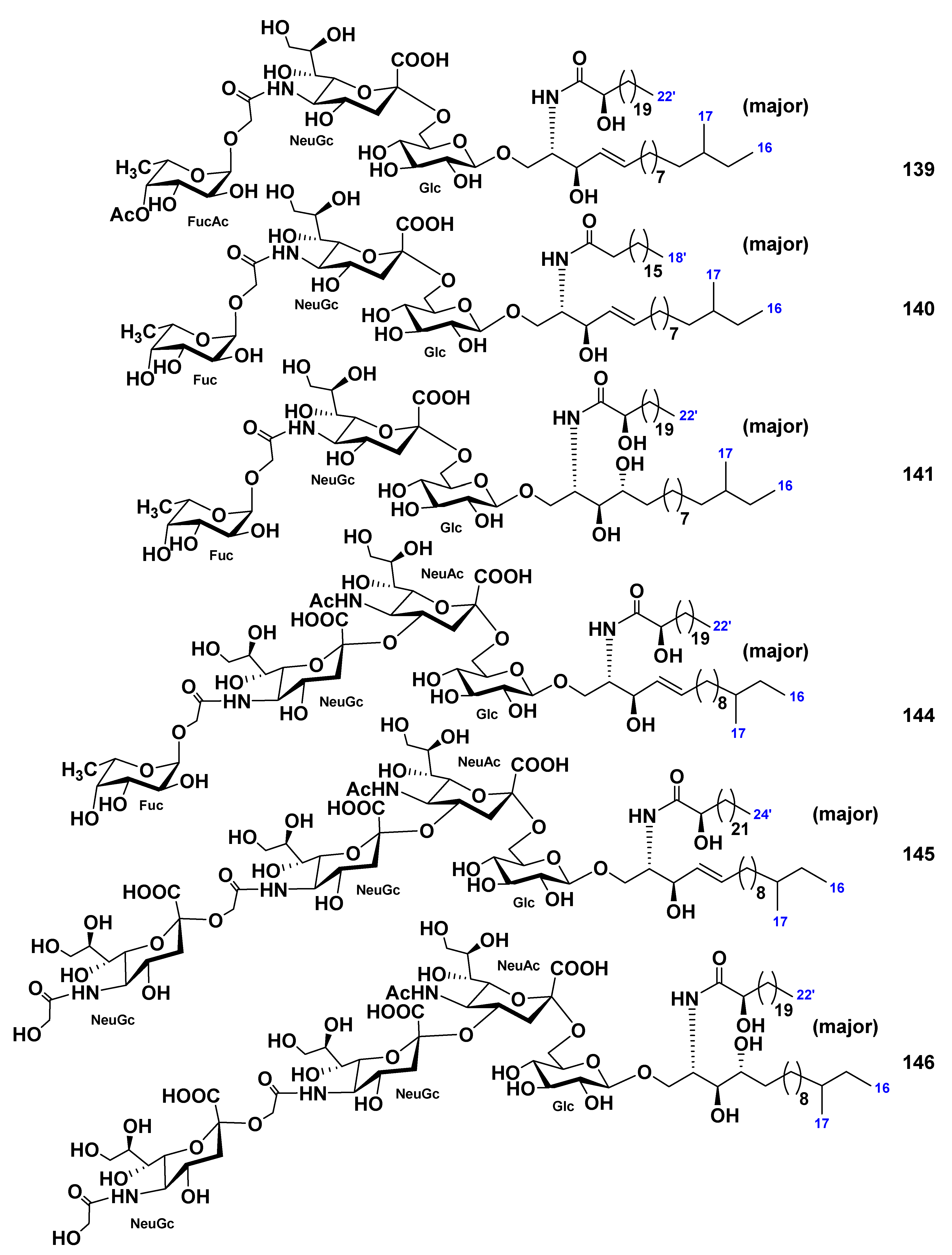
| Cucumaria echinata | 139–141, 144–146 142, 143 | Neuritogenic activity toward the rat pheochromocytoma PC12 cells in the presence of NGF | [85,86] | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malyarenko, T.V.; Kicha, A.A.; Stonik, V.A.; Ivanchina, N.V. Sphingolipids of Asteroidea and Holothuroidea: Structures and Biological Activities. Mar. Drugs 2021, 19, 330. https://doi.org/10.3390/md19060330
Malyarenko TV, Kicha AA, Stonik VA, Ivanchina NV. Sphingolipids of Asteroidea and Holothuroidea: Structures and Biological Activities. Marine Drugs. 2021; 19(6):330. https://doi.org/10.3390/md19060330
Chicago/Turabian StyleMalyarenko, Timofey V., Alla A. Kicha, Valentin A. Stonik, and Natalia V. Ivanchina. 2021. "Sphingolipids of Asteroidea and Holothuroidea: Structures and Biological Activities" Marine Drugs 19, no. 6: 330. https://doi.org/10.3390/md19060330
APA StyleMalyarenko, T. V., Kicha, A. A., Stonik, V. A., & Ivanchina, N. V. (2021). Sphingolipids of Asteroidea and Holothuroidea: Structures and Biological Activities. Marine Drugs, 19(6), 330. https://doi.org/10.3390/md19060330