
(?)-Loliolide Isolated from Sargassum horneri Abate UVB-Induced Oxidative Damage in Human Dermal Fibroblasts and Subside ECM Degradation

 ,
, 
 ,
,  , and
, and
Abstract
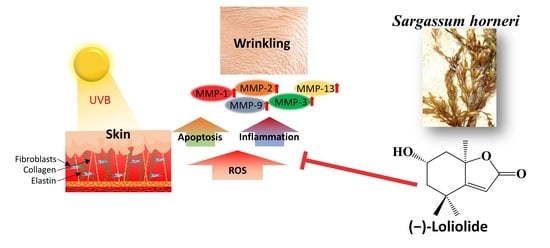
:
1. Introduction
2. Results
2.1. HTT Attenuated Intracellular ROS Level While Ameliorating HDFs Viability
2.2. HTT Abated UVB-Induced Mitochondrial Hyperpolarization and Apoptotic Body Formation
2.3. HTT Ameliorated UVB-Induced Irregularities in MAPK and NF-κB Signaling
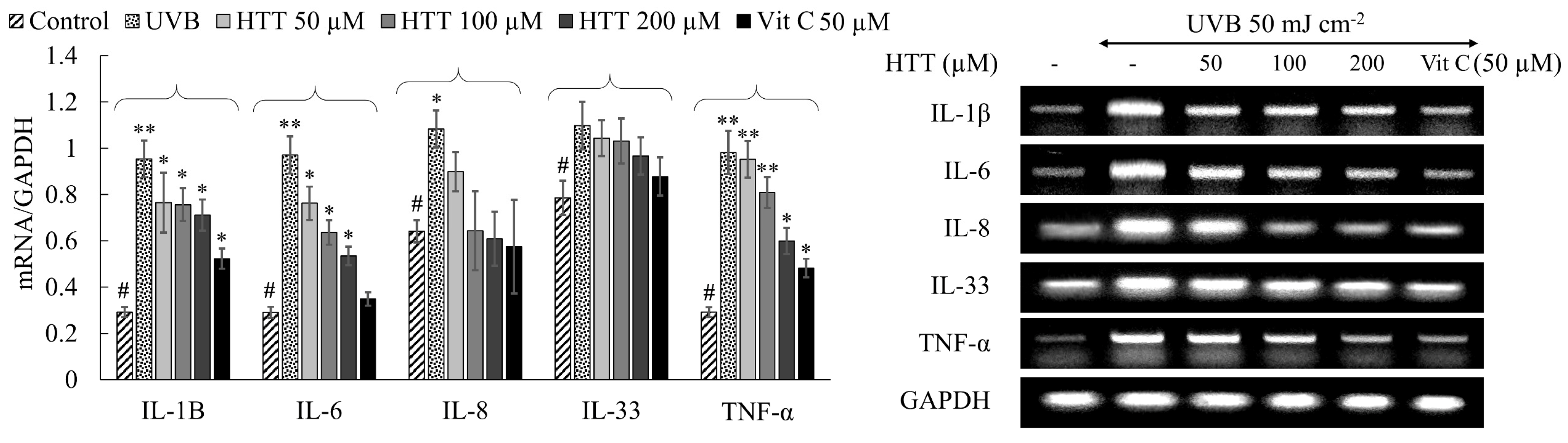
2.4. HTT Suppressed UVB-Induced Expression of Inflammatory Cytokines in HDFs
2.5. HTT Downregulated UVB-Induced MMP Expression in HDFs
2.6. HTT Reduced Collagenase and Elastase Activity in UVB-Stimulated HDFs
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. UVB Exposure of HDFs and Measuring Oxidative Stress
4.4. Evaluating Mitochondrial Depolarization by JC-1 Assay
4.5. Evaluating Apoptosis
4.6. Western Blot Analysis
4.7. RT PCR Analysis
4.8. Immunofluorescence Analysis
4.9. Analysis of Collagenase and Elastase Activity in Cell Lysates
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Makrantonaki, E.; Zouboulis, C.C. Skin anti-aging strategies. Dermatoendocrinology 2012, 4, 308–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.; Sherje, A.P. Development of resveratrol and green tea sunscreen formulation for combined photoprotective and antioxidant properties. J. Drug Deliv. Sci. Technol. 2020, 60, 102000. [Google Scholar] [CrossRef]
- Wang, L.; Kim, H.S.; Oh, J.Y.; Je, J.G.; Jeon, Y.-J.; Ryu, B. Protective effect of diphlorethohydroxycarmalol isolated from Ishige okamurae against UVB-induced damage in vitro in human dermal fibroblasts and in vivo in zebrafish. Food Chem. Toxicol. 2019, 136, 110963. [Google Scholar] [CrossRef]
- Jeanmaire, C.; Danoux, L.; Pauly, G. Glycation during human dermal intrinsic and actinic ageing: An in vivo and in vitro model study. Br. J. Dermatol. 2001, 145, 10–18. [Google Scholar] [CrossRef]
- Tanaka, N.; Tajima, S.; Ishibashi, A.; Uchida, K.; Shigematsu, T. Immunohistochemical detection of lipid peroxidation products, protein-bound acrolein and 4-hydroxynonenal protein adducts, in actinic elastosis of photodamaged skin. Arch. Dermatol. Res. 2001, 293, 363–367. [Google Scholar] [CrossRef]
- Jiratchayamaethasakul, C.; Ding, Y.; Hwang, O.; Im, S.-T.; Jang, Y.; Myung, S.-W.; Lee, J.M.; Kim, H.-S.; Ko, S.-C.; Lee, S.-H. In vitro screening of elastase, collagenase, hyaluronidase, and tyrosinase inhibitory and antioxidant activities of 22 halophyte plant extracts for novel cosmeceuticals. Fish. Aquat. Sci. 2020, 23, 6. [Google Scholar] [CrossRef]
- Di Girolamo, N.; Wakefield, D.; Coroneo, M.T. UVB-mediated induction of cytokines and growth factors in pterygium epithelial cells involves cell surface receptors and intracellular signaling. Invest. Ophthalmol. Vis. Sci. 2006, 47, 2430–2437. [Google Scholar] [CrossRef]
- Matrisian, L.M. Metalloproteinases and their inhibitors in matrix remodeling. Trends Genet. 1990, 6, 121–125. [Google Scholar] [CrossRef]
- Fisher, G.J.; Choi, H.C.; Bata-Csorgo, Z.; Shao, Y.; Datta, S.; Wang, Z.Q.; Kang, S.; Voorhees, J.J. Ultraviolet Irradiation Increases Matrix Metalloproteinase-8 Protein in Human Skin In Vivo. J. Invest. Dermatol. 2001, 117, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, M.K.H.M.; Madusanka, D.M.D.; Han, E.J.; Kim, H.-S.; Jeon, Y.-J.; Jee, Y.; Kim, K.-N.; Lee, K.; Fernando, I.P.S.; Ahn, G. Sargassum horneri (Turner) C. Agardh ethanol extract attenuates fine dust-induced inflammatory responses and impaired skin barrier functions in HaCaT keratinocytes. J. Ethnopharmacol. 2021, 273, 114003. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Wang, L.; Fernando, I.P.S.; Je, J.-G.; Ko, S.-C.; Kang, M.C.; Lee, J.M.; Yim, M.-J.; Jeon, Y.-J.; Lee, D.-S. Antioxidant efficacy of (−)-loliolide isolated from Sargassum horneri against AAPH-induced oxidative damage in Vero cells and zebrafish models in vivo. J. Appl. Phycol. 2020, 32, 3341–3348. [Google Scholar] [CrossRef]
- Dias, M.K.H.M.; Madusanka, D.M.D.; Han, E.J.; Kim, M.J.; Jeon, Y.-J.; Kim, H.-S.; Fernando, I.P.S.; Ahn, G. (−)-Loliolide Isolated from Sargassum horneri Protects against Fine Dust-Induced Oxidative Stress in Human Keratinocytes. Antioxidants 2020, 9, 474. [Google Scholar] [CrossRef]
- Kim, H.-S.; Fernando, I.P.S.; Lee, S.-H.; Ko, S.-C.; Kang, M.C.; Ahn, G.; Je, J.-G.; Sanjeewa, K.K.A.; Rho, J.-R.; Shin, H.J.; et al. Isolation and characterization of anti-inflammatory compounds from Sargassum horneri via high-performance centrifugal partition chromatography and high-performance liquid chromatography. Algal Res. 2021, 54, 102209. [Google Scholar] [CrossRef]
- Zhu, X.; Li, N.; Wang, Y.; Ding, L.; Chen, H.; Yu, Y.; Shi, X. Protective effects of quercetin on UVB irradiation-induced cytotoxicity through ROS clearance in keratinocyte cells. Oncol. Rep. 2017, 37, 209–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Park, Y.G.; Lee, H.-J.; Lim, S.J.; Nho, C.W. Youngiasides A and C Isolated from Youngia denticulatum Inhibit UVB-Induced MMP Expression and Promote Type I Procollagen Production via Repression of MAPK/AP-1/NF-κB and Activation of AMPK/Nrf2 in HaCaT Cells and Human Dermal Fibroblasts. J. Agric. Food Chem. 2015, 63, 5428–5438. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.-H.; Kim, A.; Park, J.-M.; Kim, S.-H.; Chung, A.-S. Ultraviolet B-induced matrix metalloproteinase-1 and -3 secretions are mediated via PTEN/Akt pathway in human dermal fibroblasts. J. Cell. Physiol. 2006, 209, 775–785. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Dias, M.K.H.M.; Madusanka, D.M.D.; Han, E.J.; Kim, M.J.; Jeon, Y.J.; Lee, K.; Cheong, S.H.; Han, Y.S.; Park, S.R.; et al. Human Keratinocyte UVB-Protective Effects of a Low Molecular Weight Fucoidan from Sargassum horneri Purified by Step Gradient Ethanol Precipitation. Antioxidants 2020, 9, 340. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-S.; Han, E.J.; Fernando, I.P.S.; Sanjeewa, K.K.A.; Jayawardena, T.U.; Kim, H.-J.; Jee, Y.; Kang, S.-H.; Jang, J.-H.; Jang, J.-P.; et al. Anti-allergy effect of mojabanchromanol isolated from Sargassum horneri in bone marrow-derived cultured mast cells. Algal Res. 2020, 48, 101898. [Google Scholar] [CrossRef]
- Pettit, G.R.; Herald, C.L.; Ode, R.H.; Brown, P.; Gust, D.J.; Michel, C. The isolation of loliolide from an Indian Ocean opisthobranch mollusc. J. Nat. Prod. 1980, 43, 752–755. [Google Scholar] [CrossRef]
- Hodges, R.; Porte, A.L. The structure of loliolide: A terpene from lolium perenne. Tetrahedron 1964, 20, 1463–1467. [Google Scholar] [CrossRef]
- Kimura, J.; Maki, N. New Loliolide Derivatives from the Brown Alga Undaria pinnatifida. J. Nat. Prod. 2002, 65, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.H.; Hwangbo, K.; Zheng, M.S.; Cho, J.H.; Son, J.-K.; Kim, H.Y.; Baek, S.H.; Choi, H.C.; Park, S.Y.; Kim, J.-R. Inhibitory effects of (−)-loliolide on cellular senescence in human dermal fibroblasts. Arch. Pharmacal Res. 2015, 38, 876–884. [Google Scholar] [CrossRef]
- Lee, Y.R.; Bae, S.; Kim, J.Y.; Lee, J.; Cho, D.H.; Kim, H.S.; An, I.S.; An, S. Monoterpenoid Loliolide Regulates Hair Follicle Inductivity of Human Dermal Papilla Cells by Activating the Akt/β-Catenin Signaling Pathway. J. Microbiol. Biotechnol. 2019, 29, 1830–1840. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Kim, H.S.; Wang, L.; Lee, W.W.; Jeon, Y.J. Apoptotic and antiproliferative properties of 3beta-hydroxy-Delta5-steroidal congeners from a partially purified column fraction of Dendronephthya gigantea against HL-60 and MCF-7 cancer cells. J. Appl. Toxicol. 2018, 38, 527–536. [Google Scholar] [CrossRef]
- Du, C.; Fang, M.; Li, Y.; Li, L.; Wang, X. Smac, a mitochondrial protein that promotes cytochrome c–dependent caspase activation by eliminating IAP inhibition. Cell 2000, 102, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Hwang, B.-M.; Noh, E.-M.; Kim, J.-S.; Kim, J.-M.; Hwang, J.-K.; Kim, H.-K.; Kang, J.-S.; Kim, D.-S.; Chae, H.-J.; You, Y.-O.; et al. Decursin inhibits UVB-induced MMP expression in human dermal fibroblasts via regulation of nuclear factor-κB. Int. J. Mol. Med. 2013, 31, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Jackson, B.C.; Nebert, D.W.; Vasiliou, V. Update of human and mouse matrix metalloproteinase families. Hum Genom. 2010, 4, 194–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amano, S.; Ogura, Y.; Akutsu, N.; Matsunaga, Y.; Kadoya, K.; Adachi, E.; Nishiyama, T. Protective effect of matrix metalloproteinase inhibitors against epidermal basement membrane damage: Skin equivalents partially mimic photoageing process. Br. J. Dermatol. 2005, 153 (Suppl. 2), 37–46. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, R.K.; Fakurazi, S.; Ahmad, S.; Abas, F.; Ismail, I.S.; Rukayadi, Y.; Akhtar, M.T.; Shaari, K. Inhibition of UVB-induced pro-inflammatory cytokines and MMP expression by Zanthoxylum rhetsa bark extract and its active constituent hesperidin. Phytother. Res. 2018, 32, 1608–1616. [Google Scholar] [CrossRef]
- Hay, M.; Thomas, D.W.; Craighead, J.L.; Economides, C.; Rosenthal, J. Clinical development success rates for investigational drugs. Nature Biotechnol. 2014, 32, 40–51. [Google Scholar] [CrossRef]
- Potts, R.O.; Guy, R.H. Predicting skin permeability. Pharm. Res. 1992, 9, 663–669. [Google Scholar] [CrossRef]
- Weinmüllner, R.; Zbiral, B.; Becirovic, A.; Stelzer, E.M.; Nagelreiter, F.; Schosserer, M.; Lämmermann, I.; Liendl, L.; Lang, M.; Terlecki-Zaniewicz, L. Organotypic human skin culture models constructed with senescent fibroblasts show hallmarks of skin aging. NPJ Aging Mech. Dis. 2020, 6, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, I.P.S.; Dias, M.K.H.M.; Madusanka, D.M.D.; Han, E.J.; Kim, M.J.; Jeon, Y.-J.; Ahn, G. Fucoidan refined by Sargassum confusum indicate protective effects suppressing photo-oxidative stress and skin barrier perturbation in UVB-induced human keratinocytes. Int. J. Biol. Macromol. 2020, 164, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Fernando, I.P.S.; Han, E.J.; Kim, M.J.; Jung, K.; Kang, D.S.; Ahn, C.B.; Ahn, G. In Vivo Hepatoprotective Effects of a Peptide Fraction from Krill Protein Hydrolysates against Alcohol-Induced Oxidative Damage. Mar. Drugs 2019, 17, 690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suganuma, K.; Nakajima, H.; Ohtsuki, M.; Imokawa, G. Astaxanthin attenuates the UVA-induced up-regulation of matrix-metalloproteinase-1 and skin fibroblast elastase in human dermal fibroblasts. J. Dermatol. Sci. 2010, 58, 136–142. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence (5′ to 3′ Direction) | |
|---|---|---|
| IL-1β | Forward | TGT CCT GCG TGT TGA AAG ATG A |
| Reverse | CAG GCA GTT GGG CAT TGG TG | |
| IL-6 | Forward | GAT GGC TGA AAA AGA TGG ATG C |
| Reverse | TGG TTG GGT CAG GGG TGG TT | |
| IL-8 | Forward | ACA CTG CGC CAA CAC AGA AAT TA |
| Reverse | CAG GCA GTT GGG CAT TGG TG | |
| IL-33 | Forward | GAT GAG ATG TCT CGG CTG CTT G |
| Reverse | AGC CGT TAC GGA TAT GGT GGT C | |
| TNF-α | Forward | GGC AGT CAG ATC ATC TTC TCG AA |
| Reverse | GAA GGC CTA AGG TCC ACT TGT GT | |
| MMP1 | Forward | CTGAAGGTGATGAAGCAGCC |
| Reverse | AGTCCAAGAGAATGGCCGAG | |
| MMP2 | Forward | GCGACAAGAAGTATGGCTTC |
| Reverse | TGCCAAGGTCAATGTCAGGA | |
| MMP3 | Forward | CTCACAGACCTGACTCGGTT |
| Reverse | CACGCCTGAAGGAAGAGATG | |
| MMP8 | Forward | ATGGACCAACACCTCCGCAA |
| Reverse | GTCAATTGCTTGGACGCTGC | |
| MMP9 | Forward | CGCAGACATCGTCATCCAGT |
| Reverse | GGATTGGCCTTGGAAGATGA | |
| MMP13 | Forward | CTATGGTCCAGGAGATGAAG |
| Reverse | AGAGTCTTGCCTGTATCCTC | |
| GAPDH | Forward | CGT CTA GAA AAA CCT GCC AA |
| Reverse | TGA AGT CAA AGG AGA CCA CC- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernando, I.P.S.; Heo, S.-J.; Dias, M.K.H.M.; Madusanka, D.M.D.; Han, E.-J.; Kim, M.-J.; Sanjeewa, K.K.A.; Lee, K.; Ahn, G. (?)-Loliolide Isolated from Sargassum horneri Abate UVB-Induced Oxidative Damage in Human Dermal Fibroblasts and Subside ECM Degradation. Mar. Drugs 2021, 19, 435. https://doi.org/10.3390/md19080435
Fernando IPS, Heo S-J, Dias MKHM, Madusanka DMD, Han E-J, Kim M-J, Sanjeewa KKA, Lee K, Ahn G. (?)-Loliolide Isolated from Sargassum horneri Abate UVB-Induced Oxidative Damage in Human Dermal Fibroblasts and Subside ECM Degradation. Marine Drugs. 2021; 19(8):435. https://doi.org/10.3390/md19080435
Chicago/Turabian StyleFernando, Ilekuttige Priyan Shanura, Soo-Jin Heo, Mawalle Kankanamge Hasitha Madhawa Dias, Dissanayaka Mudiyanselage Dinesh Madusanka, Eui-Jeong Han, Min-Ju Kim, Kalu Kapuge Asanka Sanjeewa, Kyounghoon Lee, and Ginnae Ahn. 2021. "(?)-Loliolide Isolated from Sargassum horneri Abate UVB-Induced Oxidative Damage in Human Dermal Fibroblasts and Subside ECM Degradation" Marine Drugs 19, no. 8: 435. https://doi.org/10.3390/md19080435
APA StyleFernando, I. P. S., Heo, S.-J., Dias, M. K. H. M., Madusanka, D. M. D., Han, E.-J., Kim, M.-J., Sanjeewa, K. K. A., Lee, K., & Ahn, G. (2021). (?)-Loliolide Isolated from Sargassum horneri Abate UVB-Induced Oxidative Damage in Human Dermal Fibroblasts and Subside ECM Degradation. Marine Drugs, 19(8), 435. https://doi.org/10.3390/md19080435