
Marine-Derived Indole Alkaloids and Their Biological and Pharmacological Activities †

 , , ,
, , ,  and
and 
Abstract
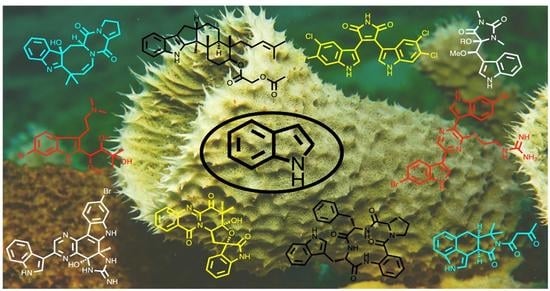
:
1. Introduction
2. Sources of Marine-Derived Indole Alkaloids Isolated from January 2016 to October 2021
2.1. Marine Microorganisms
2.1.1. Marine-Derived Fungi
Simple Indole Alkaloids
Prenylated Indoles
Bis-/Tris-Indoles
Annelated Indoles
2.1.2. Marine-Derived Bacteria
Simple Indoles
Bis-/Tris-Indoles
2.2. Marine Invertebrates
2.2.1. Marine Sponges
Simple Indole Alkaloids
Bis-/Tris-Indole Alkaloids
2.2.2. Bryozoans
Simple Indole Alkaloids
Prenylated Indoles
Annelated Indole Alkaloids
2.3. Marine Plants
2.3.1. Algae
2.3.2. Mangrove Trees
3. Biological and Pharmacological Activities of Indole Alkaloids
3.1. Antimicrobial Activity
3.2. Antiviral Activity
3.3. Anticancer Activity
3.4. Anti-Inflammatory Activity
3.5. Antidiabetic Activity
3.6. Antiparasitic Activity
3.7. Neuroprotective Activity
3.8. Enzyme Inhibitors
3.9. Other Activities
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Drugs and drug candidates from marine sources: An assessment of the current “state of play”. Planta Med. 2016, 82, 775–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gul, W.; Hamann, M.T. Indole alkaloid marine natural products: An established source of cancer drug leads with considerable promise for the control of parasitic, neurological and other diseases. Life Sci. 2005, 78, 442–453. [Google Scholar] [CrossRef] [PubMed]
- França, P.H.; Barbosa, D.P.; da Silva, D.L.; Ribeiro, Ê.A.; Santana, A.E.; Santos, B.V.; Barbosa-Filho, J.M.; Quintans, J.S.; Barreto, R.S.; Quintans-Júnior, L.J. Indole alkaloids from marine sources as potential leads against infectious diseases. BioMed Res. Int. 2014, 2014, 375423. [Google Scholar] [CrossRef]
- Netz, N.; Opatz, T. Marine indole alkaloids. Mar. Drugs 2015, 13, 4814–4914. [Google Scholar] [CrossRef] [Green Version]
- Kochanowska-Karamyan, A.J.; Hamann, M.T. Marine indole alkaloids: Potential new drug leads for the control of depression and anxiety. Chem. Rev. 2010, 110, 4489–4497. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.W.; Liu, X.J.; Yuan, J.; Li, H.J.; Mahmud, T.; Hong, M.J.; Yu, J.C.; Lan, W.J. l-Tryptophan induces a marine-derived Fusarium sp. to produce indole alkaloids with activity against the Zika virus. J. Nat. Prod. 2020, 83, 3372–3380. [Google Scholar] [CrossRef]
- Chen, Y.X.; Xu, M.Y.; Li, H.J.; Zeng, K.J.; Ma, W.Z.; Tian, G.B.; Xu, J.; Yang, D.P.; Lan, W.J. Diverse secondary metabolites from the marine-derived fungus Dichotomomyces cejpii F31-1. Mar. Drugs 2017, 15, 339. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.T.; Xu, M.Y.; Liu, W.; Li, H.J.; Xu, J.; Yang, D.P.; Lan, W.J.; Wang, L.Y. Two additional new compounds from the marine-derived fungus Pseudallescheria ellipsoidea F42-3. Molecules 2016, 21, 442. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.H.; Chen, H.Q.; Form, I.; Konuklugil, B.; Proksch, P.; Wang, B.G. New chromone, isocoumarin, and indole alkaloid derivatives from three sponge-derived fungal strains. Nat. Prod. Commun. 2016, 11, 1293–1296. [Google Scholar] [CrossRef]
- Li, C.J.; Chen, P.N.; Li, H.J.; Mahmud, T.; Wu, D.L.; Xu, J.; Lan, W.J. Potential antidiabetic fumiquinazoline alkaloids from the marine-derived fungus Scedosporium apiospermum F41-1. J. Nat. Prod. 2020, 83, 1082–1091. [Google Scholar] [CrossRef]
- Huang, L.H.; Xu, M.Y.; Li, H.J.; Li, J.Q.; Chen, Y.X.; Ma, W.Z.; Li, Y.P.; Xu, J.; Yang, D.P.; Lan, W.J. Amino acid-directed strategy for inducing the marine-derived fungus Scedosporium apiospermum F41–1 to maximize alkaloid diversity. Org. Lett. 2017, 19, 4888–4891. [Google Scholar] [CrossRef]
- May Zin, W.W.; Buttachon, S.; Dethoup, T.; Fernandes, C.; Cravo, S.; Pinto, M.M.M.; Gales, L.; Pereira, J.A.; Silva, A.M.S.; Sekeroglu, N.; et al. New cyclotetrapeptides and a new diketopiperzine derivative from the marine sponge-associated fungus Neosartorya glabra KUFA 0702. Mar. Drugs 2016, 14, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May Zin, W.W.; Buttachon, S.; Dethoup, T.; Pereira, J.A.; Gales, L.; Inácio, Â.; Costa, P.M.; Lee, M.; Sekeroglu, N.; Silva, A.M.S.; et al. Antibacterial and antibiofilm activities of the metabolites isolated from the culture of the mangrove-derived endophytic fungus Eurotium chevalieri KUFA 0006. Phytochemistry 2017, 141, 86–97. [Google Scholar] [CrossRef]
- Zhong, W.M.; Wang, J.F.; Shi, X.F.; Wei, X.Y.; Chen, Y.C.; Zeng, Q.; Yao Xiang, Y.; Chen, X.Y.; Tian, X.P.; Xiao, Z.H.; et al. Eurotiumins A–E, five new alkaloids from the marine-derived fungus Eurotium sp. SCSIO F452. Mar. Drugs 2018, 16, 136. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Feng, C.; Wang, S.Y.; Zhang, D.M.; Li, X.H.; Zhang, C.X. New indole diketopiperazine alkaloids from soft coral-associated epiphytic fungus Aspergillus sp. EGF 15-0-3. Chem. Biodivers. 2020, 17, e2000106. [Google Scholar] [CrossRef] [PubMed]
- Afiyatullov, S.H.; Zhuravleva, O.I.; Antonov, A.S.; Berdyshev, D.V.; Pivkin, M.V.; Denisenko, V.A.; Popov, R.S.; Gerasimenko, A.V.; von Amsberg, G.; Dyshlovoy, S.A.; et al. Prenylated indole alkaloids from co-culture of marine-derived fungi Aspergillus sulphureus and Isaria felina. J. Antibiot. 2018, 71, 846–853. [Google Scholar] [CrossRef]
- Li, H.; Sun, W.; Deng, M.; Zhou, Q.; Wang, J.; Liu, J.; Chen, C.; Qi, C.; Luo, Z.; Xue, Y.; et al. Asperversiamides, linearly fused prenylated indole alkaloids from the marine-derived fungus Aspergillus versicolor. J. Org. Chem. 2018, 83, 8483–8492. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.J.; Wang, K.T.; Xu, M.Y.; Zhang, J.J.; Lam, C.K.; Zhong, G.H.; Xu, J.; Yang, D.P.; Li, H.J.; Wang, L.Y. Secondary metabolites with chemical diversity from the marine-derived fungus Pseudallescheria boydii F19-1 and their cytotoxic activity. RSC Adv. 2016, 6, 76206–76213. [Google Scholar] [CrossRef]
- Zhuravleva, O.I.; Antonov, A.S.; Trang, V.T.D.; Pivkin, M.V.; Khudyakova, Y.V.; Denisenko, V.A.; Popov, R.S.; Kim, N.Y.; Yurchenko, E.A.; Gerasimenko, A.V.; et al. New deoxyisoaustamide derivatives from the coral-derived fungus Penicillium dimorphosporum KMM 4689. Mar. Drugs 2021, 19, 32. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X.; Nong, X.; Wang, J.; Qi, S. Brevianamides and mycophenolic acid derivatives from the deep-sea-derived fungus Penicillium brevicompactum DFFSCS025. Mar. Drugs 2017, 15, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Gong, L.; Guo, M.; Jiang, Y.; Ding, Y.; Wang, Z.; Xin, X.; An, F. Bioactive indole diketopiperazine alkaloids from the marine endophytic fungus Aspergillus sp. YJ191021. Mar. Drugs 2021, 19, 157. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.Y.; Shen, N.X.; Liang, Z.Y.; Shen, L.; Chen, M.; Wang, C.Y. Paraherquamide J, a new prenylated indole alkaloid from the marine-derived fungus Penicillium janthinellum HK1-6. Nat. Prod. Res. 2020, 34, 378–384. [Google Scholar] [CrossRef]
- Kong, F.D.; Fan, P.; Zhou, L.M.; Ma, Q.Y.; Xie, Q.Y.; Zheng, H.Z.; Zheng, Z.H.; Zhang, R.S.; Yuan, J.Z.; Dai, H.F.; et al. Penerpenes A–D, four indole terpenoids with potent protein tyrosine phosphatase inhibitory activity from the marine-derived fungus Penicillium sp. KFD28. Org. Lett. 2019, 21, 4864–4867. [Google Scholar] [CrossRef]
- Zhou, L.M.; Kong, F.D.; Fan, P.; Ma, Q.Y.; Xie, Q.Y.; Li, J.H.; Zheng, H.Z.; Zheng, Z.H.; Yuan, J.Z.; Dai, H.F.; et al. Indole-diterpenoids with protein tyrosine phosphatase inhibitory activities from the marine-derived fungus Penicillium sp. KFD28. J. Nat. Prod. 2019, 82, 2638–2644. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Tao, H.; Lin, X.; Wang, J.; Liao, S.; Dong, J.; Zhou, X.; Liu, Y. Prenylated indole alkaloids and chromone derivatives from the fungus Penicillium sp. SCSIO041218. Tetrahedron 2018, 74, 77–82. [Google Scholar] [CrossRef]
- Zhou, G.; Sun, C.; Hou, X.; Che, Q.; Zhang, G.; Gu, Q.; Liu, C.; Zhu, T.; Li, D. Ascandinines A–D, indole diterpenoids from the sponge-derived fungus Aspergillus candidus HDN15-152. J. Org. Chem. 2021, 86, 2431–2436. [Google Scholar] [CrossRef]
- Ivanets, E.V.; Yurchenko, A.N.; Smetanina, O.F.; Rasin, A.B.; Zhuravleva, O.I.; Pivkin, M.V.; Popov, R.S.; Von Amsberg, G.; Afiyatullov, S.H.; Dyshlovoy, S.A. Asperindoles A–D and a p-terphenyl derivative from the ascidian-derived fungus Aspergillus sp. KMM 4676. Mar. Drugs 2018, 16, 232. [Google Scholar] [CrossRef] [Green Version]
- Cai, R.; Jiang, H.; Xiao, Z.; Cao, W.; Yan, T.; Liu, Z.; Lin, S.E.; Long, Y.; She, Z. (−)-and (+)-Asperginulin A, a pair of indole diketopiperazine alkaloid dimers with a 6/5/4/5/6 pentacyclic skeleton from the mangrove endophytic fungus Aspergillus sp. SK-28. Org. Lett. 2019, 21, 9633–9636. [Google Scholar] [CrossRef]
- Li, P.; Zhang, M.; Li, H.; Wang, R.; Hou, H.; Li, X.; Liu, K.; Chen, H. New prenylated indole homodimeric and pteridine alkaloids from the marine-Derived fungus Aspergillus austroafricanus Y32-2. Mar. Drugs 2021, 19, 98. [Google Scholar] [CrossRef]
- Shaker, S.; Fan, R.Z.; Li, H.J.; Lan, W.J. A pair of novel bisindole alkaloid enantiomers from marine fungus Fusarium sp. XBB-9. Nat. Prod. Res. 2021, 35, 1497–1503. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.X.; Qiu, Y.; Ran, Y.Q.; Feng, G.K.; Deng, R.; Zhu, X.F.; Lan, W.J.; Li, H.J. Exploration of indole alkaloids from marine fungus Pseudallescheria boydii F44-1 using an amino acid-directed strategy. Mar. Drugs 2019, 17, 77. [Google Scholar] [CrossRef] [Green Version]
- Buttachon, S.; Ramos, A.A.; Inácio, Â.; Dethoup, T.; Gales, L.; Lee, M.; Costa, P.M.; Silva, A.M.S.; Sekeroglu, N.; Rocha, E.; et al. Bis-indolyl benzenoids, hydroxypyrrolidine derivatives and other constituents from cultures of the marine sponge-associated fungus Aspergillus candidus KUFA0062. Mar. Drugs 2018, 16, 119. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Chen, H.; Li, W.; Zhu, X.; Ding, W.; Li, C. Bioactive chaetoglobosins from the mangrove endophytic fungus Penicillium chrysogenum. Mar. Drugs 2016, 14, 172. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Zhou, D.; Liang, F.; Wu, Z.; She, Z.; Li, C. Penochalasin K, a new unusual chaetoglobosin from the mangrove endophytic fungus Penicillium chrysogenum V11 and its effective semi-synthesis. Fitoterapia 2017, 123, 23–28. [Google Scholar] [CrossRef]
- Qi, J.; Jiang, L.; Zhao, P.; Chen, H.; Jia, X.; Zhao, L.; Dai, H.; Hu, J.; Liu, C.; Shim, S.H.; et al. Chaetoglobosins and azaphilones from Chaetomium globosum associated with Apostichopus japonicus. Appl. Microbiol. Biot. 2020, 104, 1545–1553. [Google Scholar] [CrossRef]
- Luo, X.W.; Gao, C.H.; Lu, H.M.; Wang, J.M.; Su, Z.Q.; Tao, H.M.; Zhou, X.F.; Yang, B.; Liu, Y.H. HPLC-DAD-guided isolation of diversified chaetoglobosins from the coral-associated fungus Chaetomium globosum C2F17. Molecules 2020, 25, 1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Min, X.; Huang, J.; Zhong, Y.; Wu, Y.; Li, X.; Deng, Y.; Jiang, Z.; Shao, Z.; Zhang, L.; et al. Cytoglobosins H and I, new antiproliferative cytochalasans from deep-sea-derived fungus Chaetomium globosum. Mar. Drugs 2016, 14, 233. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Hu, Y.; Hao, X.; Tan, J.; Li, F.; Qiao, X.; Chen, S.; Xiao, C.; Chen, M.; Peng, Z.; et al. Raistrickindole A, an anti-HCV oxazinoindole alkaloid from Penicillium raistrickii IMB17-034. J. Nat. Prod. 2019, 82, 1391–1395. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.D.; Zhang, S.L.; Zhou, S.Q.; Ma, Q.Y.; Xie, Q.Y.; Chen, J.P.; Li, J.H.; Zhou, L.M.; Yuan, J.Z.; Hu, Z.; et al. Quinazoline-containing indole alkaloids from the marine-derived fungus Aspergillus sp. HNMF114. J. Nat. Prod. 2019, 82, 3456–3463. [Google Scholar] [CrossRef]
- Limbadri, S.; Luo, X.; Lin, X.; Liao, S.; Wang, J.; Zhou, X.; Yang, B.; Liu, Y. Bioactive novel indole alkaloids and steroids from deep sea-derived fungus Aspergillus fumigatus SCSIO 41012. Molecules 2018, 23, 2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.S.; Yang, L.; Kong, F.D.; Zhao, J.H.; Yao, L.; Yuchi, Z.G.; Ma, Q.Y.; Xie, Q.Y.; Zhou, L.M.; Guo, M.F.; et al. Three new quinazoline-containing indole alkaloids from the marine-derived fungus Aspergillus sp. HNMF114. Front. Microbiol. 2021, 12, 1111. [Google Scholar] [CrossRef]
- Cho, K.H.; Sohn, J.H.; Oh, H. Isolation and structure determination of a new diketopiperazine dimer from marine-derived fungus Aspergillus sp. SF-5280. Nat. Prod. Res. 2018, 32, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.L.; Xia, J.M.; Su, R.Q.; Li, J.; Liu, Y.; Yang, X.W.; Yang, Q. Bacilsubteramide A, a new indole alkaloid, from the deep-sea-derived Bacillus subterraneus 11593. Nat. Prod. Res. 2018, 32, 2553–2557. [Google Scholar] [CrossRef]
- Anjum, K.; Kaleem, S.; Yi, W.; Zheng, G.; Lian, X.; Zhang, Z. Novel antimicrobial indolepyrazines A and B from the marine-associated Acinetobacter sp. ZZ1275. Mar. Drugs 2019, 17, 89. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Yang, J.; Yu, J.; Li, J.; Yuan, J.; Wong, N.K.; Ju, J. Chlorinated bis-indole alkaloids from deep-sea derived Streptomyces sp. SCSIO 11791 with antibacterial and cytotoxic activities. J. Antibiot. 2020, 73, 542–547. [Google Scholar] [CrossRef]
- Okada, M.; Sugita, T.; Wong, C.P.; Wakimoto, T.; Abe, I. Identification of pyridinium with three indole moieties as an antimicrobial agent. J. Nat. Prod. 2017, 80, 1205–1209. [Google Scholar] [CrossRef]
- Kwon, O.S.; Ahn, S.; Jeon, J.; Park, I.G.; Won, T.H.; Sim, C.J.; Park, H.; Oh, D.C.; Oh, K.B.; Noh, M.; et al. Psammocindoles A–C: Isolation, synthesis, and bioactivity of indole-γ-lactams from the sponge Psammocinia vermis. Org. Lett. 2021, 23, 4667–4671. [Google Scholar] [CrossRef] [PubMed]
- Guzii, A.G.; Makarieva, T.N.; Denisenko, V.A.; Gerasimenko, A.V.; Udovenko, A.A.; Popov, R.S.; Dmitrenok, P.S.; Golotin, V.A.; Fedorov, S.N.; Grebnev, B.B.; et al. Guitarrins A–E and aluminumguitarrin A: 5-azaindoles from the Northwestern Pacific marine sponge Guitarra fimbriata. J. Nat. Prod. 2019, 82, 1704–1709. [Google Scholar] [CrossRef]
- Wang, Q.; Tang, X.L.; Luo, X.C.; de Voog, N.J.; Li, P.L.; Li, G.Q. Aplysinopsin-type and bromotyrosine-derived alkaloids from the south China sea sponge Fascaplysinopsis reticulata. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Di, X.; Rouger, C.; Hardardottir, I.; Freysdottir, J.; Molinski, T.F.; Tasdemir, D.; Omarsdottir, S. 6-bromoindole derivatives from the Icelandic marine sponge Geodia barretti: Isolation and anti-Inflammatory activity. Mar. Drugs 2018, 16, 437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, A.E.; Killday, K.B.; Chakrabarti, D.; Guzmán, E.A.; Harmody, D.; McCarthy, P.J.; Pitts, T.; Pomponi, S.A.; Reed, J.K.; Roberts, B.F.; et al. Dragmacidin G, a bioactive bis-indole alkaloid from a deep-water sponge of the genus Spongosorites. Mar. Drugs 2017, 15, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hitora, Y.; Takada, K.; Ise, Y.; Okada, S.; Matsunaga, S. Dragmacidins G and H, bisindole alkaloids tethered by a guanidino ethylthiopyrazine moiety, from a Lipastrotethya sp. marine sponge. J. Nat. Prod. 2016, 79, 2973–2976. [Google Scholar] [CrossRef]
- Liu, H.B.; Lauro, G.; O’Connor, R.D.; Lohith, K.; Kelly, M.; Colin, P.; Bifulco, G.; Bewley, C.A. Tulongicin, an antibacterial tri-indole alkaloid from a deep-water Topsentia sp. sponge. J. Nat. Prod. 2017, 80, 2556–2560. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Feng, Y.; Murtaza, M.; Wood, S.; Mellick, G.; Hooper, J.N.; Quinn, R.J. A grand challenge: Unbiased phenotypic function of metabolites from Jaspis splendens against Parkinson’s disease. J. Nat. Prod. 2016, 79, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Ragini, K.; Piggott, A.M.; Karuso, P. Bisindole alkaloids from a New Zealand deep-sea marine sponge Lamellomorpha strongylata. Mar. Drugs 2019, 17, 683. [Google Scholar] [CrossRef] [Green Version]
- Jennings, L.K.; Khan, N.; Kaur, N.; Rodrigues, D.; Morrow, C.; Boyd, A.; Thomas, O.P. Brominated bisindole alkaloids from the Celtic Sea sponge Spongosorites calcicola. Molecules 2019, 24, 3890. [Google Scholar] [CrossRef] [Green Version]
- Cruz, P.G.; Leal, J.F.M.; Daranas, A.H.; Pérez, M.; Cuevas, C. On the mechanism of action of dragmacidins I and J, two new representatives of a new class of protein phosphatase 1 and 2A inhibitors. ACS Omega 2018, 3, 3760–3767. [Google Scholar] [CrossRef]
- Park, J.S.; Cho, E.; Hwang, J.Y.; Park, S.C.; Chung, B.; Kwon, O.S.; Sim, C.J.; Oh, D.C.; Oh, K.B.; Shin, J. Bioactive bis (indole) alkaloids from a Spongosorites sp. sponge. Mar. Drugs 2021, 19, 3. [Google Scholar] [CrossRef] [PubMed]
- El-Hawary, S.S.; Sayed, A.M.; Mohammed, R.; Hassan, H.M.; Rateb, M.E.; Amin, E.; Mohammed, T.A.; El-Mesery, M.; Muhsinah, A.B.; Alsayari, A.; et al. Bioactive brominated oxindole alkaloids from the Red Sea sponge Callyspongia siphonella. Mar. Drugs 2019, 17, 465. [Google Scholar] [CrossRef] [Green Version]
- Moosmann, P.; Taniguchi, T.; Furihata, K.; Utsumi, H.; Ise, Y.; Morii, Y.; Yamawaki, N.; Takatani, T.; Arakawa, O.; Okada, S.; et al. Myrindole A, an antimicrobial bis-indole from a marine sponge Myrmekioderma sp. Org. Lett. 2021, 23, 3477–3480. [Google Scholar] [CrossRef] [PubMed]
- Kleks, G.; Holland, D.C.; Kennedy, E.K.; Avery, V.M.; Carroll, A.R. Antiplasmodial alkaloids from the Australian bryozoan Amathia lamourouxi. J. Nat. Prod. 2020, 83, 3435–3444. [Google Scholar] [CrossRef]
- Di, X.; Wang, S.; Oskarsson, J.T.; Rouger, C.; Tasdemir, D.; Hardardottir, I.; Freysdottir, J.; Wang, X.; Molinski, T.F.; Omarsdottir, S. Bromotryptamine and imidazole alkaloids with anti-inflammatory activity from the bryozoan Flustra foliacea. J. Nat. Prod. 2020, 83, 2854–2866. [Google Scholar] [CrossRef]
- Hansen, K.Ø.; Isaksson, J.; Bayer, A.; Johansen, J.A.; Andersen, J.H.; Hansen, E. Securamine derivatives from the Arctic bryozoan Securiflustra securifrons. J. Nat. Prod. 2017, 80, 3276–3283. [Google Scholar] [CrossRef]
- Li, M.C.; Sun, W.S.; Cheng, W.; Liu, D.; Liang, H.; Zhang, Q.Y.; Lin, W.H. Four new minor brominated indole related alkaloids with antibacterial activities from Laurencia similis. Bioorg. Med. Chem. Lett. 2016, 26, 3590–3593. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.S.; Sun, J.Z.; Tang, Q.Q.; Fan, F.; Guo, Y.W. Acanthiline A, a pyrido[1,2-a] indole alkaloid from Chinese mangrove Acanthus ilicifolius. J. Asian Nat. Prod. Res. 2018, 20, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Aoki, S.; Gato, K.; Matsunami, K.; Kurosu, M.; Kitagawa, I. Marine natural products. XXXIV. Trisindoline, a new antibiotic indole trimer, produced by a bacterium of Vibrio sp. separated from the marine sponge Hyrtios altum. Chem. Pharm. Bull. 1994, 42, 2449–2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organisms | Biological/Pharmacological Activities | Compound | Reference |
|---|---|---|---|
| 1. Marine-derived fungi | |||
| Antibacterial activity | |||
| Aspergillus sp. YJ191021 | Xanthomonas oryzae pv. oryzae X. oryzae pv. Oryzicola Edwardsiella tarda Vibrio anguillarum V. parahaemolyticus Aeromonas hydrophilia | 41 | [22] |
| A. fumigatus SCSIO 41012 | Acinetoobacter baumannii ATCC 19606, A. baumannii ATCC 15122, Klebsiella pneumoniae ATCC 14578 Staphylococcus aureus ATCC 16339 | 93 | [41] |
| Antibiofilm activity | |||
| Eurotium chevalieri KUFA 0006 | S. aureus ATCC 25923 | 9 | [14] |
| Anti-Quorum sensing activity | |||
| Aspergillus sp. HNMF114 | Chromobacterium violaceum CV026 | 91, 92 | [40] |
| Antifungal activity | |||
| Aspergillus sp. YJ191021 | Rhizoctonia solani | 45 | [22] |
| A. fumigatus SCSIO 41012 | Fusarium oxysporum f. sp. cucumerinum F. oxysporum f. sp. momordicae | 93 | [41] |
| Penicillium chrysogenum V11 | Colletotrichum gloeosporioides R. solani | 82, 83 | [34,35] |
| Antiviral activity | |||
| Fusarium sp. L1 | Anti-Zika virus | 48, 49, 74, 75 | [7] |
| A. candidus HDN15-152 | Anti-influenza A virus (H1N1) | 65 | [27] |
| P. raistrickii IMB17-034 | Anti-hepatitis C virus | 90 | [39] |
| Scedosporium apiospermum F41 | Anti-hepatitis C virus | 106, 114 | [12] |
| Anticancer activity | |||
| Aspergillus sp. KMM4676 | 22Rv1, PC-3, and LNCaP | 66, 67 | [28] |
| A. candidus KUFA0062 | Hep G2, HT29, HCT116, A549, A375, MCF7 and U251 | 80 | [33] |
| P. chrysogenum V11 | MDA-MB-435, SGC-7901,A549 | 81, 83 | [34,35] |
| Anti-inflammatory activity | |||
| Aspergillus versicolor | iNOS inhibitory activity | 17, 18, 35, 36, | [18] |
| Aspergillus sp. YJ191021 | Inhibit secretion of 1L-1β by THP-1 cells | 41, 45, 46 | [22] |
| Antidiabetic activity | |||
| S. apiospermum F41-1 | promote triglyceride accumulation in 3T3-L1 | 6 | [12] |
| Penicillium sp. KFD28 | Inhibition of protein tyrosine | 50, 51, 54, | [24] |
| Aspergillus sp. SF-5280 | phosphatases (PTPs) | 55, 57 | [25] |
| Inhibition of non-transmembrane PTPs (PTP1B) | 116 | [43] | |
| Neuroprotective activity | |||
| P. dimorphosporum KMM 4689 | increased a viability of paraquat-treated cells | 27, 29 | [30] |
| Pro-angiogenic activity | |||
| A. austroafricanus Y32-2 | Pro-angiogenic activity in a vatalanib (PTK787)-induced vascular injury zebrafish model | 73 | [30] |
| 2.Marine-derived bacteria | |||
| Antibacterial activity | |||
| Streptomyces sp. SCSIO 11791 | Micrococcus luteus ML01, S. aureus ATCC 29213, and a panel of MRSA isolated from human patients (MRSA 991, MRSA 1862, MRSA 669 A, MRSA A2) and pig (MRSA GDQ6P012P, MRSA GDE4P037P) | 119, 120 | [46] |
| Acinetobacter sp. ZZ1275 | S. aureus (MRSA), E. coli | 118, 121 | [45] |
| E. coli transfected by metagenomic DNA prepared from the marine sponge Dicderma calyx | Bacillus cereus, S. aureus (MSSA) | 122 | [47] |
| Antifungal activity | |||
| Acinetobacter sp. ZZ1275 | Candida albicans | 118, 121 | [45] |
| Anticancer activity | |||
| Streptomyces sp. SCSIO 11791 | MDA-MB-435, MDA-MB-231 NCI-H460, HCT-116, HepG2. MCF10A | 119, 120 | [46] |
| 3.Marine sponges | |||
| Antibacterial activity | |||
| Spongosorites sp. | Mycobacterium tuberculosis CDC1551 | 141 | [52] |
| Spongosorites sp. | S. aureus | 156–158, 159 | [59] |
| S. entérica | 158, 159 | [59] | |
| Topsentia sp. | S. aureus ATCC 29213 | 143, 161 | [54] |
| Callyspongia siphonella | S. aureus and B. subtilis | 162, 163, | [60] |
| Myrmekioderma sp. | E. coli and B. subtilis | 164 | [67] |
| Antiviral activity | |||
| Topsentia sp. | Anti-HIV activity | 143, 161 | [54] |
| Anticancer activity | |||
| Fascaplysinopsis reticulata | HeLa | 133a, 133b | [50] |
| Spongosorites sp. | Human pancreatic cell lines: PANC-1, MIA PaCa-2, BxPC-3, ASPC-1 | 141 | [52] |
| Dragmacidon sp. | A549, HT29, and MDA-MB-231 | 153, 154 | [58] |
| Spongosorites sp. | A549 and K562 | 155–158 | [59] |
| Anti-inflammatory activity | |||
| Geodia barretti | Decrease dendritic cell secretion of pro-inflammatory cytokine IL-12p40 and anti-inflammatory cytokine IL-10 production | 139, 140 | [59] |
| Antidiabetic activity | |||
| Psammocinia vermis | Increase adiponectin secretion during adipogenesis in hBM-MSCs | 123–125 | [48] |
| Fascaplysinopsis reticulata | inhibitory activity against PTP1B | 132a, 132b | [50] |
| Antiparasitic activity | |||
| Callyspongia siphonella | Antitrypanosomal activity against Trypanosoma brucei | 162, 163 | [60] |
| Enzyme inhibitors | |||
| Guitarra fimbriata | Inhibitor of alkaline phosphatase | 128 | [49] |
| Spongosorites sp. | Inhibit sortase A | 156, 157, 159, 160 | [59] |
| 4.Bryozoans | |||
| Anticancer activity | |||
| Securiflustra securifrons | A2058, HT-29, MCF-7, MRC-5 | 179, 180 | [64] |
| Anti-inflammatory activity | |||
| Flustra foliácea | Decrease DC secretion of the pro-inflammatory cytokine IL-12p40 | 166, 168, 170, 178 | [63] |
| 5.Algae | |||
| Antibacterial activity | |||
| Laurencia similis | S. aureus, B. subtilis, B. thuringensis, Pseudomonas lachrymans, Agrobacterium tumefaciens, Xanthomonas vesicatória, Ralstonia solanacearum | 182, 183 | [65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wibowo, J.T.; Ahmadi, P.; Rahmawati, S.I.; Bayu, A.; Putra, M.Y.; Kijjoa, A. Marine-Derived Indole Alkaloids and Their Biological and Pharmacological Activities. Mar. Drugs 2022, 20, 3. https://doi.org/10.3390/md20010003
Wibowo JT, Ahmadi P, Rahmawati SI, Bayu A, Putra MY, Kijjoa A. Marine-Derived Indole Alkaloids and Their Biological and Pharmacological Activities. Marine Drugs. 2022; 20(1):3. https://doi.org/10.3390/md20010003
Chicago/Turabian StyleWibowo, Joko Tri, Peni Ahmadi, Siti Irma Rahmawati, Asep Bayu, Masteria Yunovilsa Putra, and Anake Kijjoa. 2022. "Marine-Derived Indole Alkaloids and Their Biological and Pharmacological Activities" Marine Drugs 20, no. 1: 3. https://doi.org/10.3390/md20010003
APA StyleWibowo, J. T., Ahmadi, P., Rahmawati, S. I., Bayu, A., Putra, M. Y., & Kijjoa, A. (2022). Marine-Derived Indole Alkaloids and Their Biological and Pharmacological Activities. Marine Drugs, 20(1), 3. https://doi.org/10.3390/md20010003