
Potential Anti-Obesity, Anti-Steatosis, and Anti-Inflammatory Properties of Extracts from the Microalgae Chlorella vulgaris and Chlorococcum amblystomatis under Different Growth Conditions


 , , , ,
, , , ,  and
and
Abstract
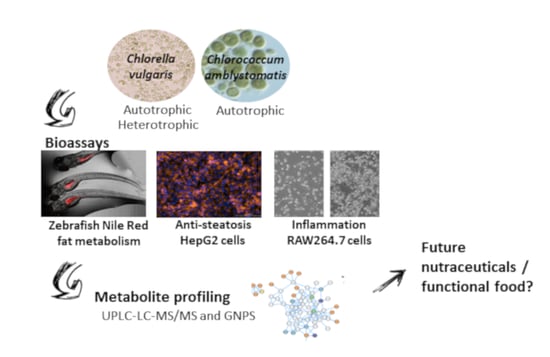
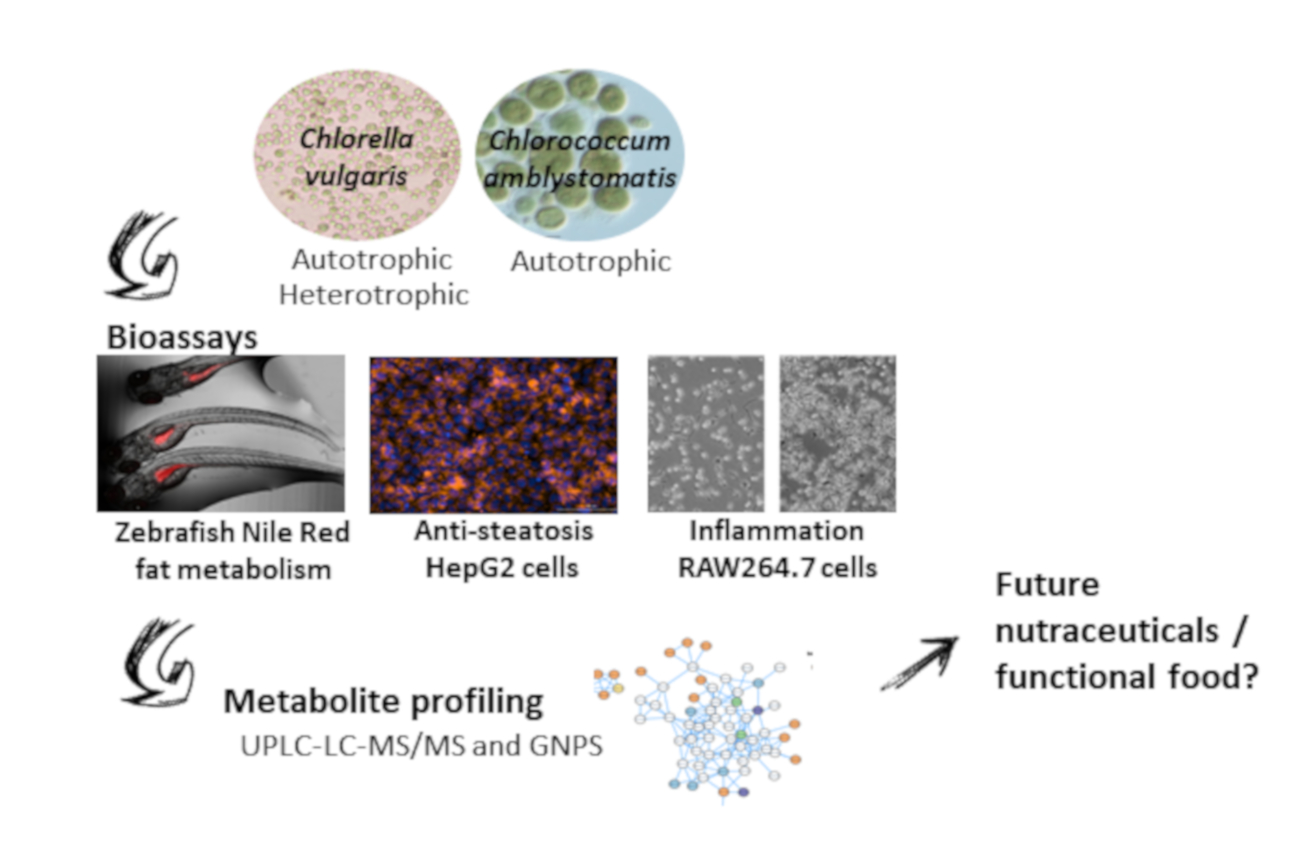
:
1. Introduction
2. Results
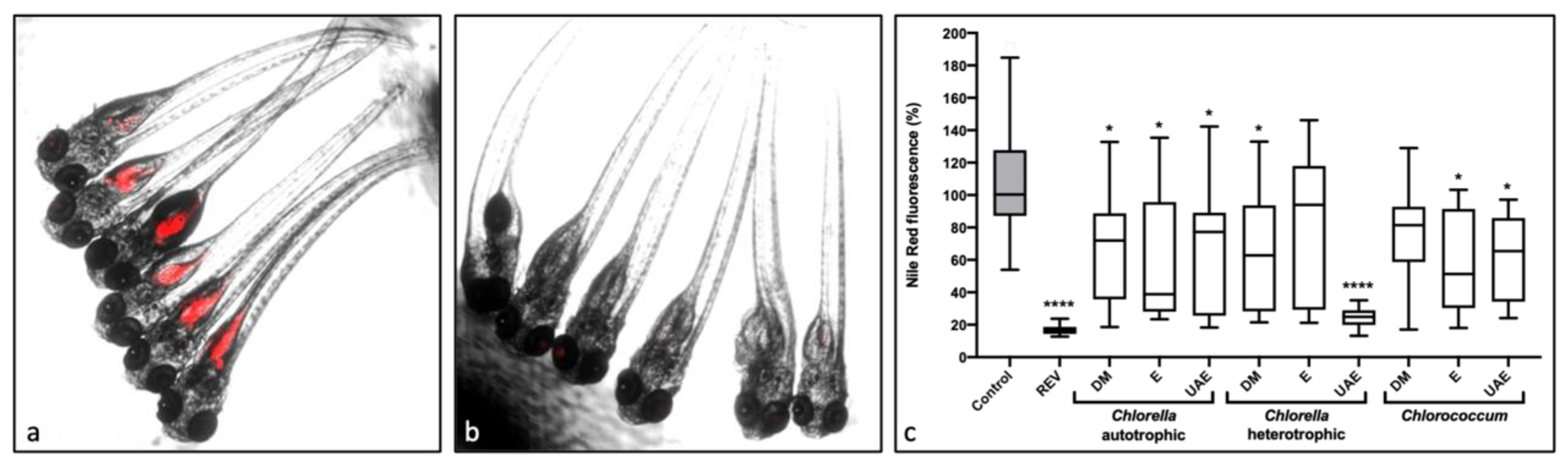
2.1. Lipid-Reducing Activity
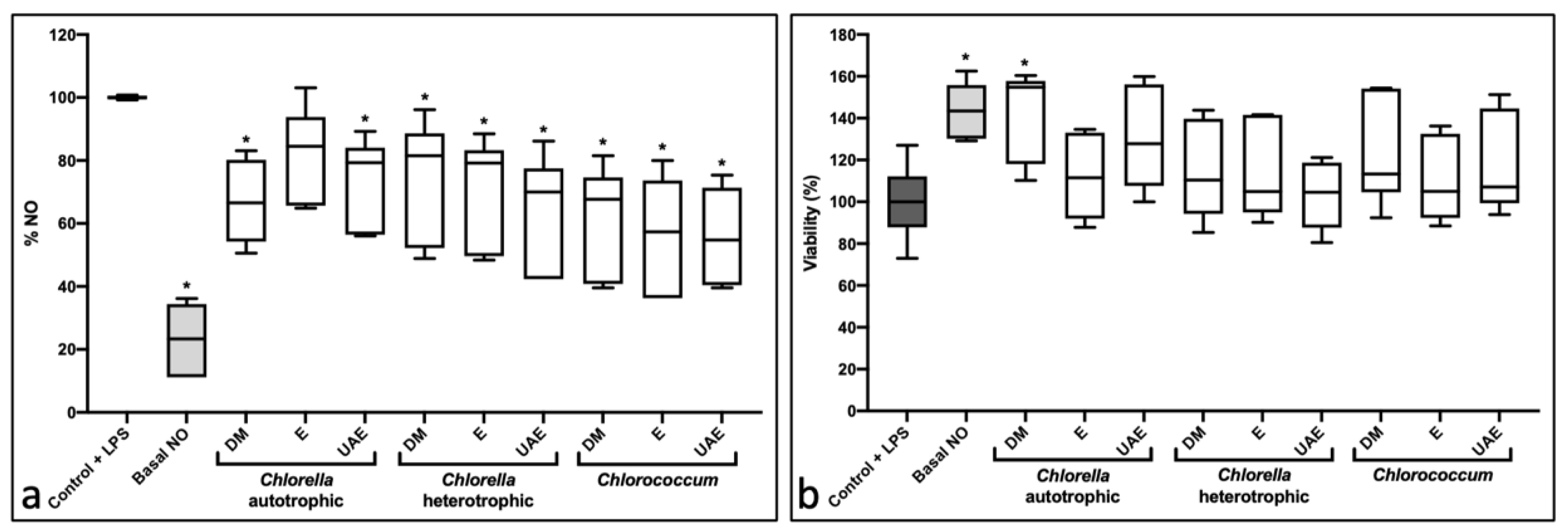
2.2. Anti-Inflammatory Activity
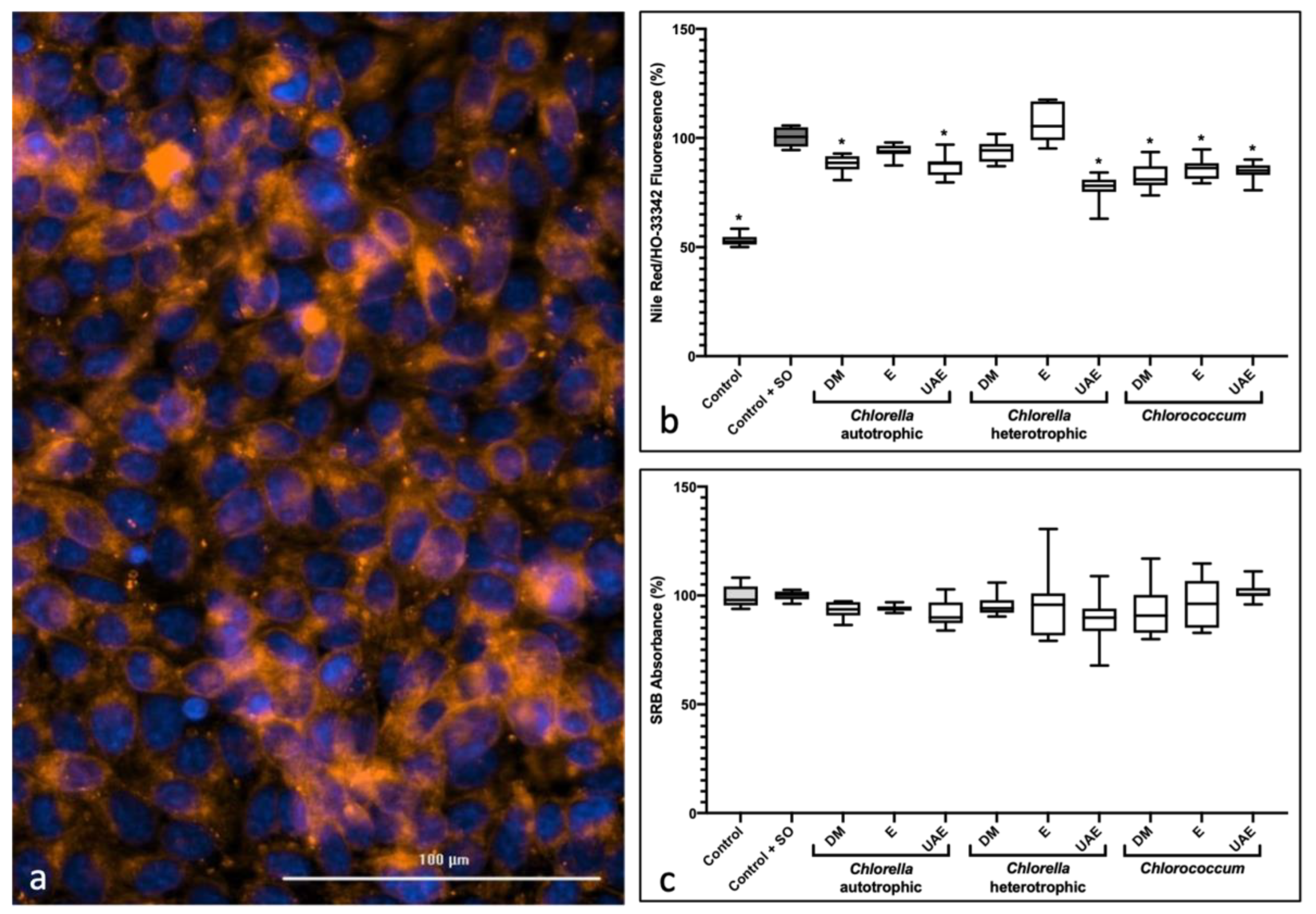
2.3. Anti-Steatosis Assay
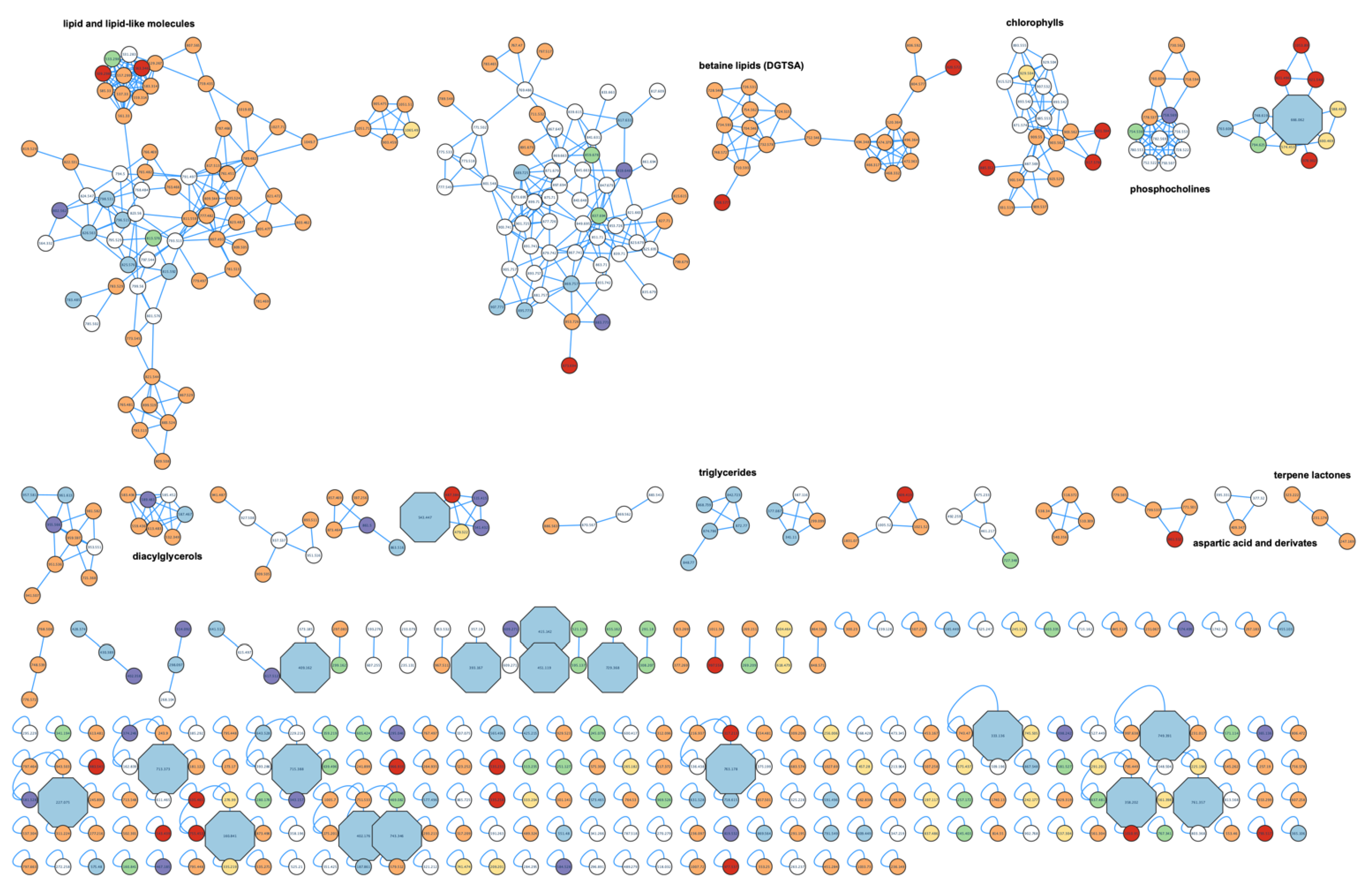
2.4. Metabolite Profiling
3. Discussion
4. Materials and Methods
4.1. Microalgae Biomass Production
4.2. Microalgae Extraction
4.3. Zebrafish Larvae Nile Red Fat Metabolism Assay
4.4. Cell Assays
4.4.1. Anti-Inflammatory Assay
4.4.2. Steatosis Assay
4.5. Metabolite Profiling
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- UNICEF, United Nations Children’s Fund; WHO, World Health Organization; WB, World Balk (WB) Joint Child Malnutrition Estimates—Levels and Trends; WHO: Geneva, Switzerland, 2017.
- González-Muniesa, P.; Mártinez-González, M.-A.; Hu, F.B.; Després, J.-P.; Matsuzawa, Y.; Loos, R.J.F.; Moreno, L.A.; Bray, G.A.; Martinez, J.A. Obesity. Nat. Rev. Dis. Primers 2017, 3, 17034. [Google Scholar] [CrossRef]
- Castro, M.; Preto, M.; Vasconcelos, V.; Urbatzka, R. Obesity: The Metabolic Disease, Advances on Drug Discovery and Natural Product Research. Curr. Top. Med. Chem. 2016, 16, 2577–2604. [Google Scholar] [CrossRef]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic Syndrome: Pathophysiology, Management, and Modulation by Natural Compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef]
- Marchisello, S.; Di Pino, A.; Scicali, R.; Urbano, F.; Piro, S.; Purrello, F.; Rabuazzo, A.M. Pathophysiological, Molecular and Therapeutic Issues of Nonalcoholic Fatty Liver Disease: An Overview. Int. J. Mol. Sci. 2019, 20, 1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettinelli, P.; Obregon, A.M.; Videla, L.A. Molecular Mechanisms of Steatosis in Nonalcoholic Fatty Liver Disease. Nutr. Hosp. 2011, 26, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Bessesen, D.H.; Van Gaal, L.F. Progress and challenges in anti-obesity pharmacotherapy. Lancet Diabetes Endocrinol. 2018, 6, 237–248. [Google Scholar] [CrossRef]
- Urbatzka, R.; Freitas, S.; Palmeira, A.; Almeida, T.; Moreira, J.; Azevedo, C.; Afonso, C.; Correia-da-Silva, M.; Sousa, E.; Pinto, M.; et al. Lipid Reducing Activity and Toxicity Profiles of a Library of Polyphenol Derivatives. Eur. J. Med. Chem. 2018, 151, 272–284. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Biodiversity: A Continuing Source of Novel Drug Leads. Pure Appl. Chem. 2005, 77, 7–24. [Google Scholar] [CrossRef] [Green Version]
- Becker, E.W. Micro-Algae as a Source of Protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R.; Folarin, A.; Huberty, A.; Interlandi, S.; Kilham, S.S.; McCauley, E.; Schulz, K.L.; et al. Nutritional Constraints in Terrestrial and Freshwater Food Webs. Nature 2000, 408, 578–580. [Google Scholar] [CrossRef]
- Koyande, A.K.; Chew, K.W.; Rambabu, K.; Tao, Y.; Chu, D.-T.; Show, P.-L. Microalgae: A Potential Alternative to Health Supplementation for Humans. Food Sci. Hum. 2019, 8, 16–24. [Google Scholar] [CrossRef]
- Rizwan, M.; Mujtaba, G.; Memon, S.A.; Lee, K.; Rashid, N. Exploring the Potential of Microalgae for New Biotechnology Applications and Beyond: A Review. Renew. Sustain. Energy Rev. 2018, 92, 394–404. [Google Scholar] [CrossRef]
- Ge, S.; Qiu, S.; Tremblay, D.; Viner, K.; Champagne, P.; Jessop, P.G. Centrate Wastewater Treatment With Chlorella vulgaris: Simultaneous Enhancement of Nutrient Removal, Biomass and Lipid Production. Chem. Eng. J. 2018, 342, 310–320. [Google Scholar] [CrossRef]
- Sydney, E.B.; Sydney, A.C.N.; de Carvalho, J.C.; Soccol, C.R. Chapter 4—Potential Carbon Fixation of Industrially Important Microalgae. In Biofuels from Algae (Second Edition); Pandey, A., Chang, J.-S., Soccol, C.R., Lee, D.-J., Chisti, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 67–88. [Google Scholar]
- Panahi, Y.; Darvishi, B.; Jowzi, N.; Beiraghdar, F.; Sahebkar, A. Chlorella vulgaris: A Multifunctional Dietary Supplement with Diverse Medicinal Properties. Curr. Pharm. Des. 2016, 22, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Couto, D.; Melo, T.; Conde, T.A.; Costa, M.; Silva, J.; Domingues, M.R.M.; Domingues, P. Chemoplasticity of the Polar Lipid Profile of the Microalgae Chlorella vulgaris Grown Under Heterotrophic and Autotrophic Conditions. Algal Res. 2021, 53, 102128. [Google Scholar] [CrossRef]
- Pantami, H.A.; Ahamad Bustamam, M.S.; Lee, S.Y.; Ismail, I.S.; Mohd Faudzi, S.M.; Nakakuni, M.; Shaari, K. Comprehensive GCMS and LC-MS/MS Metabolite Profiling of Chlorella vulgaris. Mar. Drugs 2020, 18, 367. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Wang, X.; Liu, W.; Feng, J.; Liu, Q.; Nan, F.; Jiao, X.; Xie, S. The Performance of a Self-Flocculating Microalga Chlorococcum sp. GD in Wastewater with Different Ammonia Concentrations. Int. J. Environ. Res. 2018, 15, 434. [Google Scholar] [CrossRef] [Green Version]
- Laje, K.; Seger, M.; Dungan, B.; Cooke, P.; Polle, J.; Holguin, F.O. Phytoene Accumulation in the Novel Microalga Chlorococcum sp. Using the Pigment Synthesis Inhibitor Fluridone. Mar. Drugs 2019, 17, 187. [Google Scholar] [CrossRef] [Green Version]
- Conde, T.A.; Couto, D.; Melo, T.; Costa, M.; Silva, J.; Domingues, M.R.; Domingues, P. Polar Lipidomic Profile Shows Chlorococcum amblystomatis as a Promising Source of Value-Added Lipids. Sci. Rep.-UK 2021, 11, 4355. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Cholinesterase Inhibitory Activity, Antioxidant Properties, and Phytochemical Composition of Chlorococcum sp. Extracts. J. Food Biochem. 2021, 45, e13395. [Google Scholar] [CrossRef]
- Jones, K.S.; Alimov, A.P.; Rilo, H.L.; Jandacek, R.J.; Woollett, L.A.; Penberthy, W.T. A High Throughput Live Transparent Animal Bioassay to Identify Non-Toxic Small Molecules or Genes That Regulate Vertebrate Fat Metabolism for Obesity Drug Development. Nutr. Metab. (Lond) 2008, 5, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.; Rosa, F.; Ribeiro, T.; Hernandez-Bautista, R.; Bonaldo, M.; Gonçalves Silva, N.; Eiríksson, F.; Thorsteinsdóttir, M.; Ussar, S.; Urbatzka, R. Identification of Cyanobacterial Strains With Potential for the Treatment of Obesity-Related Co-Morbidities by Bioactivity, Toxicity Evaluation and Metabolite Profiling. Mar. Drugs 2019, 17, 280. [Google Scholar] [CrossRef] [Green Version]
- Freitas, S.; Silva, N.G.; Sousa, M.L.; Ribeiro, T.; Rosa, F.; Leão, P.N.; Vasconcelos, V.; Reis, M.A.; Urbatzka, R. Chlorophyll Derivatives from Marine Cyanobacteria with Lipid-Reducing Activities. Mar. Drugs 2019, 17, 229. [Google Scholar] [CrossRef] [Green Version]
- Sano, T.; Tanaka, Y. Effect of Dried, Powdered Chlorella vulgaris on Experimental Atherosclerosis and Alimentary Hypercholesterolemia in Cholesterol-Fed Rabbits. Artery 1987, 14, 76–84. [Google Scholar] [CrossRef]
- Shibata, S.; Oda, K.; Onodera-Masuoka, N.; Matsubara, S.; Kikuchi-Hayakawa, H.; Ishikawa, F.; Iwabuchi, A.; Sansawa, H. Hypocholesterolemic Effect of Indigestible Fraction of Chlorella regularis in Cholesterol-Fed Rats. J. Nutr. Sci. Vitaminol. 2001, 47, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Park, H.J.; Kim, M.K. Effect of Chlorella vulgaris on Lipid Metabolism in Wistar Rats Fed High Fat Diet. Nutr. Res. Pract. 2008, 2, 204–210. [Google Scholar] [CrossRef]
- Chen, B.; Wan, C.; Mehmood, M.A.; Chang, J.-S.; Bai, F.; Zhao, X. Manipulating Environmental Stresses and Stress Tolerance of Microalgae for Enhanced Production of Lipids and Value-Added Products–A Review. Bioresour. Technol. 2017, 244, 1198–1206. [Google Scholar] [CrossRef]
- Xie, T.; Xia, Y.; Zeng, Y.; Li, X.; Zhang, Y. Nitrate Concentration-Shift Cultivation to Enhance Protein Content of Heterotrophic Microalga Chlorella vulgaris: Over-Compensation Strategy. Bioresour. Technol. 2017, 233, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Barros, A.; Pereira, H.; Campos, J.; Marques, A.; Varela, J.; Silva, J. Heterotrophy as a Tool to Overcome the Long and Costly Autotrophic Scale-Up Process for Large Scale Production of Microalgae. Sci. Rep.-UK 2019, 9, 13935. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Sun, Z.; Gerken, H. Recent Advances in Microalgal Biotechnology; OMICS Group eBooks: Foster City, CA, USA, 2016. [Google Scholar]
- Xu, H.; Miao, X.; Wu, Q. High Quality Biodiesel Production From a Microalga Chlorella protothecoides by Heterotrophic Growth in Fermenters. J. Biotechnol. 2006, 126, 499–507. [Google Scholar] [CrossRef]
- Bellver, M.; Costa, S.L.D.; Sanchez, B.A.; Vasconcelos, V.; Urbatzka, R. Inhibition of Intestinal Lipid Absorption by Cyanobacterial Strains in Zebrafish Larvae. Mar. Drugs 2021, 19, 161. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.D.; Vitorino, I.; De la Cruz, M.; Díaz, C.; Cautain, B.; Annang, F.; Pérez-Moreno, G.; Gonzalez Martinez, I.; Tormo, J.R.; Martín, J.M.; et al. Bioactivities and Extract Dereplication of Actinomycetales Isolated From Marine Sponges. Front. Microbiol. 2019, 10, 727. [Google Scholar] [CrossRef] [PubMed]
- Bel Mabrouk, S.; Reis, M.; Sousa, M.L.; Ribeiro, T.; Almeida, J.R.; Pereira, S.; Antunes, J.; Rosa, F.; Vasconcelos, V.; Achour, L.; et al. The Marine Seagrass Halophila stipulacea as a Source of Bioactive Metabolites Against Obesity and Biofouling. Mar. Drugs 2020, 18, 88. [Google Scholar] [CrossRef] [Green Version]
- Sibi, G.; Rabina, S. Inhibition of Pro-Inflammatory Mediators and Cytokines by Chlorella vulgaris Extracts. Pharmacogn. Res. 2016, 8, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Kwak, J.H.; Baek, S.H.; Woo, Y.; Han, J.K.; Kim, B.G.; Kim, O.Y.; Lee, J.H. Beneficial Immunostimulatory Effect of Short-Term Chlorella Supplementation: Enhancement of Natural Killercell Activity and Early Inflammatory Response (Randomized, Double-Blinded, Placebo-Controlled Trial). Nutr. J. 2012, 11, 53. [Google Scholar] [CrossRef] [Green Version]
- Soontornchaiboon, W.; Joo, S.S.; Kim, S.M. Anti-Inflammatory Effects of Violaxanthin Isolated From Microalga Chlorella ellipsoidea in Raw 264. 7 Macrophages. Biol. Pharm. Bull. 2012, 35, 1137–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polyzos, S.A.; Kountouras, J.; Mantzoros, C.S. Obesity and Nonalcoholic Fatty Liver Disease: From Pathophysiology to Therapeutics. Metabolism 2019, 92, 82–97. [Google Scholar] [CrossRef] [PubMed]
- Marcheafave, G.G.; Tormena, C.D.; Pauli, E.D.; Rakocevic, M.; Bruns, R.E.; Scarminio, I.S. Experimental Mixture Design Solvent Effects on Pigment Extraction and Antioxidant Activity from Coffea arabica L. Leaves. Microchem. J. 2019, 146, 713–721. [Google Scholar] [CrossRef]
- Ribeiro de Souza, E.B.; da Silva, R.R.; Afonso, S.; Scarminio, I.S. Enhanced Extraction Yields and Mobile Phase Separations by Solvent Mixtures for the Analysis of Metabolites in Annona muricata L. Leaves. J. Sep. Sci. 2009, 32, 4176–4185. [Google Scholar] [CrossRef]
- Cai, Z.; Qu, Z.; Lan, Y.; Zhao, S.; Ma, X.; Wan, Q.; Jing, P.; Li, P. Conventional, Ultrasound-Assisted, and Accelerated-Solvent Extractions of Anthocyanins From Purple Sweet Potatoes. Food Chem. 2016, 197, 266–272. [Google Scholar] [CrossRef]
- Medina-Torres, N.; Ayora-Talavera, T.; Espinosa-Andrews, H.; Sánchez-Contreras, A.; Pacheco, N. Ultrasound Assisted Extraction for the Recovery of Phenolic Compounds from Vegetable Sources. Agronomy 2017, 7, 47. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, C.; Quirantes-Piné, R.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Optimization of Extraction Method to Obtain a Phenolic Compounds-Rich Extract from Moringa oleifera Lam Leaves. Ind. Crops Prod. 2015, 66, 246–254. [Google Scholar] [CrossRef]
- Correia, N.; Pereira, H.; Silva, J.T.; Santos, T.; Soares, M.; Sousa, C.B.; Schüler, L.M.; Costa, M.; Varela, J.; Pereira, L.; et al. Isolation, Identification and Biotechnological Applications of a Novel, Robust, Free-living Chlorococcum (Oophila) amblystomatis Strain Isolated from a Local Pond. Appl. Sci. 2020, 10, 3040. [Google Scholar] [CrossRef]
- Figueiredo, A.R.P.; da Costa, E.; Silva, J.; Domingues, M.R.; Domingues, P. The Effects of Different Extraction Methods of Lipids from Nannochloropsis oceanica on the Contents of Omega-3 Fatty Acids. Algal Res. 2019, 41, 101556. [Google Scholar] [CrossRef]
- Lopes, G.; Clarinha, D.; Vasconcelos, V. Carotenoids from Cyanobacteria: A Biotechnological Approach for the Topical Treatment of Psoriasis. Microorganisms 2020, 8, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, I.; Girão, M.; Alexandrino, D.A.M.; Ribeiro, T.; Santos, C.; Pereira, F.; Mucha, A.P.; Urbatzka, R.; Leão, P.N.; Carvalho, M.F. Diversity and Bioactive Potential of Actinobacteria Isolated from a Coastal Marine Sediment in Northern Portugal. Microorganisms 2020, 8, 1691. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Mohimani, H.; Gurevich, A.; Mikheenko, A.; Garg, N.; Nothias, L.-F.; Ninomiya, A.; Takada, K.; Dorrestein, P.C.; Pevzner, P.A. Dereplication of Peptidic Natural Products Through Database Search of Mass Spectra. Nat. Chem. Biol. 2017, 13, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Mohimani, H.; Gurevich, A.; Shlemov, A.; Mikheenko, A.; Korobeynikov, A.; Cao, L.; Shcherbin, E.; Nothias, L.-F.; Dorrestein, P.C.; Pevzner, P.A. Dereplication of Microbial Metabolites Through Database Search of Mass Spectra. Nat. Commun. 2018, 9, 4035. [Google Scholar] [CrossRef] [Green Version]
- Feunang, Y.D.; Eisner, R.; Knox, C.; Chepelev, L.; Hastings, J.; Owen, G.; Fahy, E.; Steinbeck, C.; Subramanian, S.; Bolton, E.; et al. ClassyFire: Automated Chemical Classification with a Comprehensive, Computable Taxonomy. J. Cheminform. 2016, 8, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dictionary of Natural Products 30.1. Available online: https://dnp.chemnetbase.com/faces/chemical/ChemicalSearch.xhtml (accessed on 30 November 2021).
- Van Santen, J.A.; Jacob, G.; Singh, A.L.; Aniebok, V.; Balunas, M.J.; Bunsko, D.; Neto, F.C.; Castaño-Espriu, L.; Chang, C.; Clark, T.N.; et al. The Natural Products Atlas: An Open Access Knowledge Base for Microbial Natural Products Discovery. ACS Cent. Sci. 2019, 5, 1824–1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M + H+ | RT | Putative Identification | ppm | Formula | Source |
|---|---|---|---|---|---|
| 358.202 | 519.548 | Benzanoid | GNPS | ||
| 409.162 | 549.5865 | 2,6-Diamino-2,6-dideoxyidose; L-form, Dibenzyl dithioacetal or 3-(4-Hydroxybenzyl)-3,6-bis(methylthio)-2,5-piperazinedione; (3R,6R)-form, O-(3-Methyl-2-butenyl), 1,4-N-di-Me | 0.1 | C20H28N2O3S2 | DNP |
| Urauchimycin C | 2.2 | C19H24N2O8 | DNP | ||
| 333.136 | 749.917 | Anhydrodehydrotylophorinidine; 3-O-De-Me | −1.5 | C21H18NO3 | DNP |
| Pandangolide 2; Me ester | −3.6 | C15H24O6S | DNP | ||
| Xanthine; 7H-form, 1,7-Dibenzyl | 2.5 | C19H16N4O2 | DNP | ||
| 393.167 | 652.9033 | 7,8-Dihydroxy-1-methyl-β-carboline; 3,4-Dihydro, O7-Me, 8-O-β-D-Glucopyranoside | 2.1 | C19H24N2O7 | DNP |
| 749.391 | 550.2385 | Biscarpamontamine A or Conodiparine A; 19’-Ketone or Conodiparine B; 19’-Ketone or Conodirinine A or Conodirinine B or Coryzeylamine or Tabercorymine A or Tabernaricatine B; 19R,20S-Epoxide or Tabernaricatine B; 19S,20R-Epoxide or Tabernaricatine D; Δ1’,2’-Isomer, 7’β-hydroxy | −0.6 | C44H52N4O7 | DNP |
| 451.119 | 735.559 | Aspergillazine B or Aspergillazine B; 2-Epimer | 3.3 | C20H22N2O8S | DNP |
| 2,2’,3,3’,7,7’-Hexahydroxy-1,1’-biphenanthrene or 2,2’,4,4’,7,7’-Hexahydroxy-1,1’-biphenanthrene or 2,2’,4,4’,7,7’-Hexahydroxy-1,3’-biphenanthrene or 2,4,4’,5,5’,7’-Hexahydroxy-1,1’-biphenanthrene or 3,3’,4,4’,7,7’-Hexahydroxy-1,1’-biphenanthrene or 2,4,4’,7,7’-Pentahydroxy-1,2’-biphenanthrene ether or 2,4,5’,7,7’-Pentahydroxy-1,2’-biphenanthrene ether | 1.8 | C28H18O6 | DNP | ||
| Rhizoferrin; (R,R)-form, 2-Oxo | −2.3 | C16H22N2O13 | DNP | ||
| Aspergillazine C or Penispirozine C ou Perispirozine D | −3.3 | C20H22N2O8S | NPA | ||
| 729.368 | 732.944500 | 2,15-Dihydroxy-18-nor-16-kauren-19-oic acid; (ent-2α,15β)-form, 2-O-[β-D-Glucopyranosyl-(1→3)-2-O-(3-methylbutanoyl)-β-D-glucopyranoside] or 3,5,11,14-Tetrahydroxycard-20(22)-enolide; (3β,5β,11α,14β)-form, 3-O-[3-O-Methyl-β-D-glucopyranosyl-(1→4)-6-deoxy-α-L-glucopyranoside] | −2.4 | C36H56O15 | DNP |
| 227.075 | 648.9525 | 3-Buten-1-ol; 4-Methylbenzenesulfonyl or 3-Buten-1-ol; 4-Methylbenzenesulfonyl or 2,4-Dihydroxy-3,5,6-trimethylthiobenzoic acid; S-Me ester or 4-Phenyl-3-buten-1-ol; (Z)-form, Methanesulfonyl | 3.6 | C11H14O3S | DNP |
| 1-(2’,4’-dihydroxy-5’-methyl-3’-methylsulfanylmethylphenyl)ethanone or Mortivinacin A | 3.4 | C11H14O3S | NPA | ||
| 666.062 | 979.23 | ||||
| 543.447 | 887.89275 | ||||
| 415.142 | 734.2515 | ||||
| 743.346 | 590.652499 | ||||
| 402.176 | 711.7333 | ||||
| 160.841 | 387.5715 | ||||
| 761.357 | 554.2535 | ||||
| 763.178 | 827.886 | ||||
| 715.388 | 650.252 | ||||
| 713.373 | 775.4403 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Regueiras, A.; Huguet, Á.; Conde, T.; Couto, D.; Domingues, P.; Domingues, M.R.; Costa, A.M.; Silva, J.L.d.; Vasconcelos, V.; Urbatzka, R. Potential Anti-Obesity, Anti-Steatosis, and Anti-Inflammatory Properties of Extracts from the Microalgae Chlorella vulgaris and Chlorococcum amblystomatis under Different Growth Conditions. Mar. Drugs 2022, 20, 9. https://doi.org/10.3390/md20010009
Regueiras A, Huguet Á, Conde T, Couto D, Domingues P, Domingues MR, Costa AM, Silva JLd, Vasconcelos V, Urbatzka R. Potential Anti-Obesity, Anti-Steatosis, and Anti-Inflammatory Properties of Extracts from the Microalgae Chlorella vulgaris and Chlorococcum amblystomatis under Different Growth Conditions. Marine Drugs. 2022; 20(1):9. https://doi.org/10.3390/md20010009
Chicago/Turabian StyleRegueiras, Ana, Álvaro Huguet, Tiago Conde, Daniela Couto, Pedro Domingues, Maria Rosário Domingues, Ana Margarida Costa, Joana Laranjeira da Silva, Vitor Vasconcelos, and Ralph Urbatzka. 2022. "Potential Anti-Obesity, Anti-Steatosis, and Anti-Inflammatory Properties of Extracts from the Microalgae Chlorella vulgaris and Chlorococcum amblystomatis under Different Growth Conditions" Marine Drugs 20, no. 1: 9. https://doi.org/10.3390/md20010009