
Unusual Ether Lipids and Branched Chain Fatty Acids in Sea Cucumber (Cucumaria frondosa) Viscera and Their Seasonal Variation

Abstract
:1. Introduction
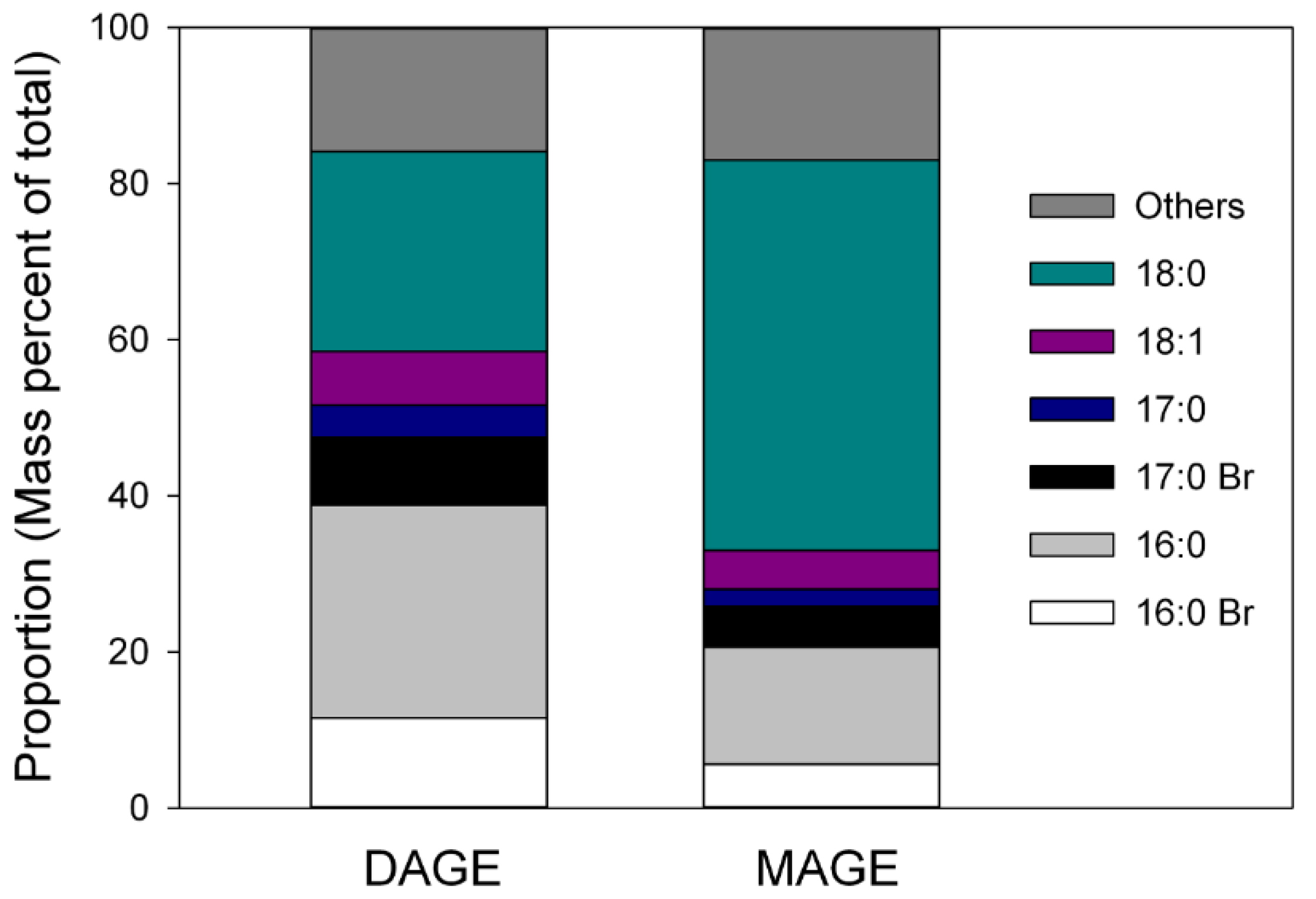
2. Results
3. Discussion
4. Materials and Methods
4.1. Moisture and Lipid Content
4.2. Lipid Class Analysis
4.3. Isolation of Lipid Classes by TLC
4.4. 1H NMR and 13C NMR Analysis
4.5. Ether Lipid Identification Using GC-MS
4.6. GC-FID Analysis of FAME
4.7. Analysis of 3-Ppyridylcarbinol Ester Derivatives
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Department of Fisheries and Aquaculture (DFA). Sea Cucumber Survey Conducted; Project Report FDP 358-4; Fisheries Diversification Program, Department of Fisheries and Aquaculture: St. John’s, NL, Canada, 2002.
- Mamelona, J.; Saint-Louis, R.; Pelletier, E. Proximate composition and nutritional profile of by-products from green urchin and Atlantic sea cucumber processing plants. Int. J. Food Sci. Technol. 2010, 45, 2119–2126. [Google Scholar] [CrossRef]
- Department of Fisheries and Oceans (DFO). Integrated Fisheries Management Plan Summary: Sea Cucumber (Apostichopus californicus) by Dive Pacific Region 2021/2022; Department of Fisheries and Oceans (DFO): Ottawa, ON, Canada, 2022.
- Vaidya, H.; Cheema, S.K. Sea cucumber and blue mussel: New sources of phospholipid enriched omega-3 fatty acids with a potential role in 3T3-L1 adipocyte metabolism. Food Funct. 2014, 5, 3287–3295. [Google Scholar] [CrossRef] [PubMed]
- Rybin, V.; Pavel, K.; Mitrofanov, D. 1-O-Alkylglycerol ether lipids in two Holothurian species: Cucumaria japonica and C. okhotensis. Nat. Prod. Commun. 2007, 2, 1934578X0700200913. [Google Scholar] [CrossRef] [Green Version]
- Isay, S.V.; Makarchenko, M.A.; Vaskovsky, V.E. A study of glyceryl ethers—I. content of α-glyceryl ethers in marine invertebrates from the sea of Japan and tropical regions of the Pacific Ocean. Comp. Biochem. Physiol. 1976, 55B, 301–305. [Google Scholar] [CrossRef]
- Haraldsson, G.; Kristinsson, B. Separation of eicosapentaenoic acid and docosahexaenoic acid in fish oil by kinetic resolution using lipase. J. Am. Oil Chem. Soc. 1998, 75, 1551–1556. [Google Scholar] [CrossRef]
- Hayashi, K.; Kishimura, H. Amount and composition of diacyl glyceryl ethers in various tissue lipids of the deep-sea squid Berryteuthis magister. J. Oleo Sci. 2002, 51, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Mollinedo, F. Antitumour ether lipids: Proapoptotic agents with multiple therapeutic indications. Expert Opin. Ther. Pat. 2007, 17, 385–405. [Google Scholar] [CrossRef]
- Jaffrès, P.A.; Gajate, C.; Bouchet, A.M.; Couthon-Gourvès, H.; Chantôme, A.; Potier-Cartereau, M.; Besson, P.; Bougnoux, P.; Mollinedo, F.; Vandier, C. Alkyl ether lipids, ion channels and lipid raft reorganization in cancer therapy. Pharmacol. Ther. 2016, 165, 114–131. [Google Scholar] [CrossRef]
- Torres, C.F.; Vázquez, L.; Señoráns, F.J.; Reglero, G. Enzymatic synthesis of short-chain diacylated alkylglycerols: A kinetic study. Process Biochem. 2009, 44, 1025–1031. [Google Scholar] [CrossRef]
- Martin, D.; Morán-Valero, M.; Señoráns, F.; Reglero, G.; Torres, C. In vitro intestinal bioaccessibility of alkylglycerols versus triacylglycerols as vehicles of butyric acid. Lipids 2011, 46, 277–285. [Google Scholar] [CrossRef]
- Zhong, Y.; Khan, M.A.; Shahidi, F. Compositional characteristics and antioxidant properties of fresh and processed sea cucumber (Cucumaria frondosa). J. Agric. Food Chem. 2007, 55, 1188–1192. [Google Scholar] [CrossRef] [PubMed]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 fatty acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Collin, P.D.; Yang, P.; Newman, R. Methods and Compositions for Treating Lipoxygenase—Mediated Disease States. U.S. Patent 6,541,519, 1 April 2003. [Google Scholar]
- Dalsgaard, J.; St John, M.; Kattner, G.; Müller-Navarra, D.; Hagen, W. Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.B.; Shurvell, H.F.; Lightner, D.A.; Cooks, R.G. The chemical shift. In Introduction to Organic Spectroscopy; Macmillan Publishing Company: New York, NY, USA, 1987; pp. 44–66. [Google Scholar]
- Santos, V.L.C.S.; Billett, D.S.M.; Wolff, G.A. 1-O-alkylglyceryl ether lipids of the gut walls and contents of an Abyssal Holothurian (Oneirophanta mutabilis). J. Braz. Chem. Soc. 2002, 13, 653–657. [Google Scholar] [CrossRef] [Green Version]
- Hallgren, B.; Larsson, S. The glyceryl ethers in the liver oils of elasmobranch fish. J. Lipid Res. 1962, 3, 31–38. [Google Scholar] [CrossRef]
- Guella, G.; Mancini, I.; Pietra, F. (+)-Raspailyne-a, a novel, acid-sensitive acetylenic enol ether glyceride from the marine sponge Raspialia pumila. J. Chem. Soc. Chem. Commun. 1986, 1, 77–78. [Google Scholar] [CrossRef]
- Myers, B.L.; Crews, P. Chiral ether glycerides from a marine sponge. J. Organ. Chem. 1983, 48, 3583–3585. [Google Scholar] [CrossRef]
- Smith, G.M.; Djerassi, C. Phospholipid studies of marine organisms: 14. Ether lipids of the sponge Tethya aurantia. Lipids 1987, 22, 236–240. [Google Scholar] [CrossRef]
- Boer, M.; Gannefors, C.; Kattner, G.; Graeve, M.; Hop, H.; Falk-Petersen, S. The arctic pteropod Clione limacina: Seasonal lipid dynamics and life-strategy. Mar. Biol. 2005, 147, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Imbs, A.; Demina, O.; Demidkova, D. Lipid class and fatty acid composition of the boreal soft coral soft coral Gersemia rubiformis. Lipids 2006, 41, 721–725. [Google Scholar] [CrossRef]
- Bakes, M.J.; Nichols, P.D. Lipid, fatty acid and squalene composition of liver oil from six species of deep-sea sharks collected in southern Australian water. Comp. Biochem. Physiol. B 1995, 110, 267–275. [Google Scholar] [CrossRef]
- Reichwald-Hacker, I. Substrate specificity of enzymes catalyzing the biosynthesis of ether lipids. In Ether Lipids: Biochemical and Biomedical Aspects; Mangold, H.K., Paltauf, F., Eds.; Academic Press: New York, NY, USA, 1983; pp. 129–140. [Google Scholar]
- Cheng, J.B.; Russell, D.W. Mammalian wax biosynthesis. I. Identification of two fatty acyl-coenzyme A reductases with different substrate specificities and tissue distributions. J. Biol. Chem. 2004, 279, 37789–37797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartvigsen, K.; Ravandi, A.; Harkewicz, R.; Kamido, H.; Bukhave, K.; Holmer, G.; Kuksis, A. 1-O-alkyl-2-(-oxo)acyl-sn-glycerols from shark oil and human milk fat are potential precursors of PAF mimics and GHB. Lipids 2006, 41, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, C.D.; Haraldsson, G.G. Ether lipids. Chem. Phys. Lipids 2011, 164, 315–340. [Google Scholar] [CrossRef] [PubMed]
- Brites, P.; Waterham, H.R.; Ronald, J.A.; Wanders, R.J.A. Functions and biosynthesis of plasmalogens in health and disease. Biochim. Biophys. Acta 2004, 1636, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Ratnayake, W.M.N.; Olsson, B.; Ackman, R.G. Novel branched chain fatty acids in certain fish oils. Lipids 1989, 24, 630–637. [Google Scholar] [CrossRef]
- Hauff, S.; Vetter, W. Quantification of branched chain fatty acids in polar and neutral lipids of cheese and fish samples. J. Agric. Food Chem. 2010, 58, 707–712. [Google Scholar] [CrossRef]
- Ackman, R.G. Fatty acids. In Marine Biogenic Lipids Fats and Oils; Ackman, R.G., Ed.; CRC Press: Boca Raton, FL, USA, 1989; pp. 103–138. [Google Scholar]
- Wallace, M.; Green, C.R.; Roberts, L.S.; Lee, Y.M.; McCarville, J.L.; Sanchez-Gurmaches, J.; Meurs, N.; Gengatharan, J.M.; Hover, J.D.; Phillips, S.A.; et al. Enzyme promiscuity drives branched-chain fatty acid synthesis in adipose tissues. Nat. Chem. Biol. 2018, 14, 1021–1031. [Google Scholar] [CrossRef]
- Kaneda, T. iso-Fatty and anteiso-fatty acids in bacteria—Biosynthesis, function, and taxonomic significance. Microbiol. Rev. 1991, 55, 288–302. [Google Scholar] [CrossRef]
- Budge, S.M.; Iverson, S.J.; Bowen, W.D.; Ackman, R.G. Among- and within-species variability in fatty acid signatures of marine fish and invertebrates on the Scotian Shelf, Georges Bank, and southern Gulf of St. Lawrence. Can. J. Fish. Aquat. Sci. 2002, 59, 886–898. [Google Scholar] [CrossRef] [Green Version]
- Iverson, S.J.; Frost, K.J.; Lang, S.L.C. Fat content and fatty acid composition of forage fish and invertebrates in Prince William Sound, Alaska: Factors contributing to among and within species variability. Mar. Ecol. Progr. Ser. 2002, 241, 161–181. [Google Scholar] [CrossRef] [Green Version]
- Bimbo, A.P. Chapter 4—Processing of marine oils. In Long-Chain Omega-3 Specialty Oils; Breivik, H., Ed.; The Oily Press Ltd.: Dundee, UK, 2012; pp. 77–109. [Google Scholar]
- American Oil Chemists’ Society (AOCS). Official Methods and Recommended Practices of the American Oil Chemists’ Society, 4th ed.; AOCS Press: Champaign, IL, USA, 1990. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Guillen, M.D.; Ruiz, A. Study of the oxidative stability of salted and unsalted salmon fillets by H nuclear magnetic resonance. Food Chem. 2004, 86, 297–304. [Google Scholar] [CrossRef]
- Christie, W.W. Gas Chromatography and Lipids; The Oily Press Ltd.: Dundee, UK, 1989; p. 36. [Google Scholar]
- Renkonen, O. Individual molecular species of phospholipids: III. Molecular species of ox-brain lecithins. Biochim. Biophys. Acta 1966, 125, 288–309. [Google Scholar] [CrossRef]
- Budge, S.M.; Iverson, S.J.; Koopman, H.N. Studying trophic ecology in marine ecosystems using fatty acids: A primer on analysis and interpretation. Mar. Mammal Sci. 2006, 22, 759–801. [Google Scholar] [CrossRef]
- Destaillats, F.; Angers, P. One-step methodology for the synthesis of FA picolinyl esters from intact lipids. J. Am. Oil Chem. Soc. 2002, 79, 253–256. [Google Scholar] [CrossRef]
- Dubois, N.; Barthomeuf, C.; Bergé, J.P. Convenient preparation of picolinyl derivatives from fatty acid esters. Eur. J. Lipid Sci. Technol. 2006, 108, 28–32. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Component | Winter | Spring | Summer | |||
|---|---|---|---|---|---|---|
| Total lipid (wwb *) | 7.36 | (0.21) a | 4.93 | (0.34) b | 5.30 | (0.29) b |
| Total lipid (dwb *) | 28.87 | (1.92) a | 20.81 | (1.22) b | 23.83 | (1.46) b |
| Moisture content (%) | 74.47 | (1.09) b | 76.29 | (0.08) a | 77.75 | (0.37) a |
| Total EPA (%) | 24.74 | (0.68) b | 25.38 | (0.18) b | 28.23 | (0.40) a |
| Total PUFA (%) | 27.31 | (0.62) c | 28.03 | (0.27) b | 29.72 | (0.42) a |
| 12-MTA (%) | 21.27 | (0.77) a | 19.17 | (0.93) b | 11.79 | (0.78) c |
| Total Branched FA (%) | 31.13 | (0.75) a | 27.84 | (0.94) b | 18.21 | (0.87) c |
| MAGE | DAGE | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Winter | Spring | Summer | Winter | Spring | Summer | |||||||
| 4,8,12-Me-13:0 | 0.42 | (0.00) b | 0.38 | (0.02) c | 0.57 | (0.05) a | 3.65 | (0.05) e | 3.32 | (0.19) f | 2.51 | (0.34) g |
| Me-14:0(a) * | 0.10 | (0.00) a | 0.09 | (0.00) a | 0.11 | (0.01) a | 0.85 | (0.03) e | 0.84 | (0.04) e | 0.80 | (0.01) f |
| Me-14:0(b) * | 0.17 | (0.03) b | 0.17 | (0.02) b | 0.39 | (0.04) a | 0.75 | (0.01) e | 0.75 | (0.02) e | 0.74 | (0.04) e |
| Me-14:0(c) * | 0.08 | (0.00) b | 0.06 | (0.00) c | 0.11 | (0.00) a | 0.67 | (0.03) e | 0.57 | (0.02) f | 0.45 | (0.04) g |
| 12-MTA | 4.81 | (0.19) a | 4.95 | (0.28) a | 5.79 | (0.32) a | 42.21 | (0.61) e | 41.09 | (0.81) e | 37.12 | (0.01) f |
| 8,12-Me-14:0 | 0.44 | (0.03) a | 0.30 | (0.03) b | 0.52 | (0.02) a | 3.33 | (0.45) e | 2.14 | (0.29) f | 2.74 | (0.14) g |
| 12-Me-15:0 | 2.09 | (0.38) b | 2.31 | (0.24) b | 3.98 | (0.20) a | 7.06 | (0.22) e | 6.84 | (0.48) e | 5.19 | (0.02) f |
| 8,12-Me-15:0 | 0.13 | (0.04) ab | 0.11 | (0.01) b | 0.19 | (0.04) a | 0.48 | (0.01) e | 0.44 | (0.07) e | 0.31 | (0.01) f |
| ai-17:0 | 0.11 | (0.01) ab | 0.09 | (0.01) b | 0.13 | (0.00) a | 0.56 | (0.02) f | 0.57 | (0.03) f | 0.77 | (0.02) e |
| 14:0 | 0.16 | (0.01) b | 0.17 | (0.02) b | 0.56 | (0.05) a | 0.53 | (0.03) f | 0.61 | (0.01) f | 1.40 | (0.16) e |
| 15:0 | 0.03 | (0.01) b | 0.13 | (0.01) a | 0.00 | (0.00) b | 0.23 | (0.02) f | 1.37 | (0.22) e | 0.00 | (0.00) g |
| 16:0 | 0.76 | (0.05) b | 0.75 | (0.08) b | 3.15 | (0.17) a | 0.68 | (0.05) f | 0.72 | (0.07) f | 1.99 | (0.14) e |
| 18:0 | 0.99 | (0.02) b | 0.92 | (0.11) b | 2.43 | (0.08) a | 0.50 | (0.07) f | 0.49 | (0.07) f | 1.14 | (0.47) e |
| 20:0 | 0.03 | (0.01) b | 0.05 | (0.04) b | 0.19 | (0.06) a | 0.04 | (0.01) f | 0.08 | (0.02) e | 0.03 | (0.02) f |
| 22:0 | 0.09 | (0.06) b | 0.14 | (0.12) ab | 0.24 | (0.00) a | 0.10 | (0.04) f | 0.12 | (0.02) ef | 0.16 | (0.02) e |
| 16:1n-9 | 1.19 | (0.45) ab | 1.39 | (0.29) a | 0.90 | (0.06) b | 0.15 | (0.03) f | 0.17 | (0.03) f | 0.27 | (0.02) e |
| 16:1n-7 | 11.16 | (0.32) b | 11.20 | (0.32) b | 13.00 | (0.47) a | 20.65 | (0.51) e | 20.85 | (0.30) e | 19.54 | (0.82) f |
| 18:1n-9 | 0.48 | (0.03) b | 0.44 | (0.05) b | 1.02 | (0.01) a | 0.86 | (0.04) f | 0.96 | (0.14) f | 2.16 | (0.12) e |
| 18:1n-7 | 2.50 | (0.14) b | 2.41 | (0.29) b | 3.11 | (0.08) a | 2.74 | (0.06) e | 2.84 | (0.17) e | 2.77 | (0.11) e |
| 20:1n-11 | 0.21 | (0.02) a | 0.22 | (0.02) ab | 0.19 | (0.01) b | 0.20 | (0.01) g | 0.26 | (0.03) f | 0.32 | (0.01) e |
| 20:1n-9 | 0.44 | (0.01) a | 0.43 | (0.05) a | 0.53 | (0.02) a | 0.29 | (0.00) g | 0.40 | (0.04) f | 0.53 | (0.03) e |
| 20:1n-7 | 0.08 | (0.01) b | 0.01 | (0.01) c | 0.37 | (0.01) a | 0.07 | (0.01) f | 0.04 | (0.00) g | 1.42 | (0.51) e |
| 22:1n-9 | 0.14 | (0.05) a | 0.12 | (0.03) a | 0.19 | (0.00) a | 0.09 | (0.02) f | 0.14 | (0.03) f | 0.26 | (0.03) e |
| 22:1n-7 | 0.11 | (0.04) a | 0.10 | (0.02) a | 0.15 | (0.00) a | 0.13 | (0.02) f | 0.15 | (0.02) ef | 0.19 | (0.01) e |
| 24:1 | 1.29 | (0.08) a | 1.32 | (0.48) a | 1.06 | (0.02) a | 0.62 | (0.04) e | 0.65 | (0.07) e | 0.62 | (0.07) e |
| EPA | 61.79 | (0.32) a | 62.26 | (0.22) a | 53.17 | (0.71) b | 5.27 | (0.28) g | 6.29 | (0.85) f | 9.10 | (0.22) e |
| DHA | 1.06 | (0.02) a | 1.12 | (0.06) a | 1.08 | (0.05) b | 0.30 | (0.02) f | 0.39 | (0.08) e | 0.43 | (0.01) e |
| Others | 7.81 | (0.43) a | 7.07 | (0.44) a | 4.91 | (0.69) b | 5.36 | (0.15) e | 5.32 | (0.22) e | 5.22 | (0.37) e |
| Sum | 100 | 100 | 100 | 100 | 100 | 100 | ||||||
| PL | TAG | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Winter | Spring | Summer | Winter | Spring | Summer | |||||||
| 4,8,12-Me-13:0 | 0.08 | (0.01) a | 0.06 | (0.01) a | 0.10 | (0.04) a | 3.98 | (0.21) e | 4.11 | (0.53) e | 2.87 | (0.37) f |
| Me-14:0(a) * | 0.00 | (0.00) b | 0.00 | (0.00) b | 0.44 | (0.03) a | 0.57 | (0.04) e | 0.61 | (0.03) e | 0.29 | (0.03) f |
| Me-14:0(b) * | 0.14 | (0.01) a | 0.14 | (0.01) a | 0.06 | (0.04) b | 0.52 | (0.02) e | 0.55 | (0.02) e | 0.42 | (0.13) e |
| Me-14:0(c) | 0.02 | (0.01) a | 0.00 | (0.00) a | 0.03 | (0.01) a | 0.30 | (0.02) e | 0.28 | (0.05) e | 0.12 | (0.02) f |
| 12-MTA | 0.46 | (0.03) a | 0.32 | (0.04) b | 0.45 | (0.14) b | 19.15 | (0.81) e | 19.87 | (0.92) e | 9.99 | (0.74) f |
| 8,12-Me-14:0 | 0.04 | (0.02) b | 0.02 | (0.01) b | 0.17 | (0.06) a | 1.75 | (0.25) e | 1.27 | (0.12) f | 1.04 | (0.08) g |
| 12-Me-15:0 | 0.21 | (0.03) b | 2.39 | (0.88) a | 3.58 | (0.30) a | 2.56 | (0.29) e | 2.51 | (0.30) e | 1.33 | (0.01) f |
| 8,12-Me-15:0 | 0.00 | (0.00) b | 0.04 | (0.01) a | 0.03 | (0.00) ab | 0.18 | (0.04) e | 0.17 | (0.04) e | 0.06 | (0.02) f |
| ai-17:0 | 0.44 | (0.09) a | 0.38 | (0.00) a | 0.43 | (0.21) a | 1.25 | (0.07) g | 1.31 | (0.01) e | 0.91 | (0.02) f |
| 14:0 | 0.25 | (0.11) a | 0.16 | (0.02) b | 0.28 | (0.11) ab | 3.43 | (0.07) g | 3.82 | (0.10) f | 5.94 | (0.25) e |
| 15:0 | 0.03 | (0.02) a | 0.04 | (0.01) a | 0.00 | (0.00) b | 0.45 | (0.24) f | 0.91 | (0.28) e | 0.00 | (0.00) g |
| 16:0 | 5.06 | (0.24) b | 2.09 | (0.31) c | 5.18 | (1.06) a | 3.11 | (0.07) g | 3.43 | (0.13) f | 4.35 | (0.32) e |
| 18:0 | 12.14 | (0.65) a | 6.43 | (0.13) b | 7.99 | (1.11) b | 3.00 | (0.06) f | 2.97 | (0.10) f | 3.71 | (0.44) e |
| 20:0 | 1.03 | (0.09) a | 1.21 | (0.02) a | 0.23 | (0.07) b | 0.26 | (0.02) f | 0.36 | (0.01) e | 0.26 | (0.02) f |
| 22:0 | 0.96 | (0.05) b | 1.08 | (0.04) b | 1.65 | (0.24) a | 0.31 | (0.01) ef | 0.35 | (0.09) e | 0.25 | (0.02) f |
| 16:1n-9 | 0.27 | (0.08) c | 2.96 | (0.85) a | 1.93 | (0.26) b | 0.14 | (0.01) f | 0.15 | (0.01) ef | 0.17 | (0.00) e |
| 16:1n-7 | 1.85 | (0.24) b | 1.51 | (0.06) c | 3.08 | (0.07) a | 23.73 | (0.41) f | 24.15 | (0.31) f | 27.61 | (0.62) e |
| 18:1n-9 | 1.17 | (0.04) a | 1.31 | (0.06) a | 1.77 | (0.62) a | 1.99 | (0.05) g | 2.18 | (0.07) f | 2.75 | (0.17) e |
| 18:1n-7 | 4.21 | (0.42) a | 4.16 | (0.25) a | 3.38 | (0.24) b | 2.50 | (0.04) ef | 2.79 | (0.14) e | 2.34 | (0.27) f |
| 20:1n-11 | 2.30 | (0.08) a | 2.74 | (0.06) a | 1.90 | (0.47) b | 0.65 | (0.05) f | 0.85 | (0.04) e | 0.38 | (0.04) g |
| 20:1n-9 | 1.03 | (0.10) a | 1.02 | (0.04) a | 1.77 | (0.58) a | 0.60 | (0.01) e | 0.70 | (0.04) e | 0.63 | (0.08) e |
| 20:1n-7 | 1.18 | (0.18) a | 0.60 | (0.13) b | 0.98 | (0.33) b | 0.20 | (0.08) f | 0.12 | (0.01) f | 1.06 | (0.23) e |
| 22:1n-9 | 1.20 | (0.35) a | 1.20 | (0.04) a | 1.16 | (0.37) a | 0.47 | (0.03) f | 0.57 | (0.03) e | 0.48 | (0.01) f |
| 22:1n-7 | 1.38 | (0.17) b | 1.46 | (0.04) b | 2.07 | (0.06) a | 0.50 | (0.03) f | 0.56 | (0.03) e | 0.38 | (0.02) g |
| 24:1 | 1.80 | (0.15) a | 1.50 | (0.12) c | 2.09 | (0.04) b | 0.90 | (0.07) e | 0.96 | (0.05) e | 0.62 | (0.19) f |
| EPA | 51.79 | (2.82) a | 55.20 | (0.72) a | 43.45 | (0.43) b | 16.71 | (0.20) f | 14.06 | (0.82) g | 20.06 | (0.41) e |
| DHA | 1.66 | (0.24) a | 1.73 | (0.11) a | 1.38 | (0.33) b | 0.92 | (0.07) e | 0.86 | (0.03) e | 0.79 | (0.02) f |
| Others | 8.04 | (0.57) a | 9.01 | (0.68) a | 11.57 | (0.72) a | 5.76 | (0.16) f | 5.73 | (0.14) f | 6.22 | (0.29) e |
| Sum | 100 | 100 | 100 | 100 | 100 | 100 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abuzaytoun, R.; Budge, S.M.; Xia, W.; MacKinnon, S. Unusual Ether Lipids and Branched Chain Fatty Acids in Sea Cucumber (Cucumaria frondosa) Viscera and Their Seasonal Variation. Mar. Drugs 2022, 20, 435. https://doi.org/10.3390/md20070435
Abuzaytoun R, Budge SM, Xia W, MacKinnon S. Unusual Ether Lipids and Branched Chain Fatty Acids in Sea Cucumber (Cucumaria frondosa) Viscera and Their Seasonal Variation. Marine Drugs. 2022; 20(7):435. https://doi.org/10.3390/md20070435
Chicago/Turabian StyleAbuzaytoun, Reem, Suzanne M. Budge, Wei Xia, and Shawna MacKinnon. 2022. "Unusual Ether Lipids and Branched Chain Fatty Acids in Sea Cucumber (Cucumaria frondosa) Viscera and Their Seasonal Variation" Marine Drugs 20, no. 7: 435. https://doi.org/10.3390/md20070435
APA StyleAbuzaytoun, R., Budge, S. M., Xia, W., & MacKinnon, S. (2022). Unusual Ether Lipids and Branched Chain Fatty Acids in Sea Cucumber (Cucumaria frondosa) Viscera and Their Seasonal Variation. Marine Drugs, 20(7), 435. https://doi.org/10.3390/md20070435