

Effect of Post-Extraction Ultrasonication on Compositional Features and Antioxidant Activities of Enzymatic/Alkaline Extracts of Palmaria palmata





Abstract
1. Introduction
2. Results
2.1. Protein Content, Degree of Hydrolysis, Protein Recovery, and Nitrogen-to-Protein Conversion Factor
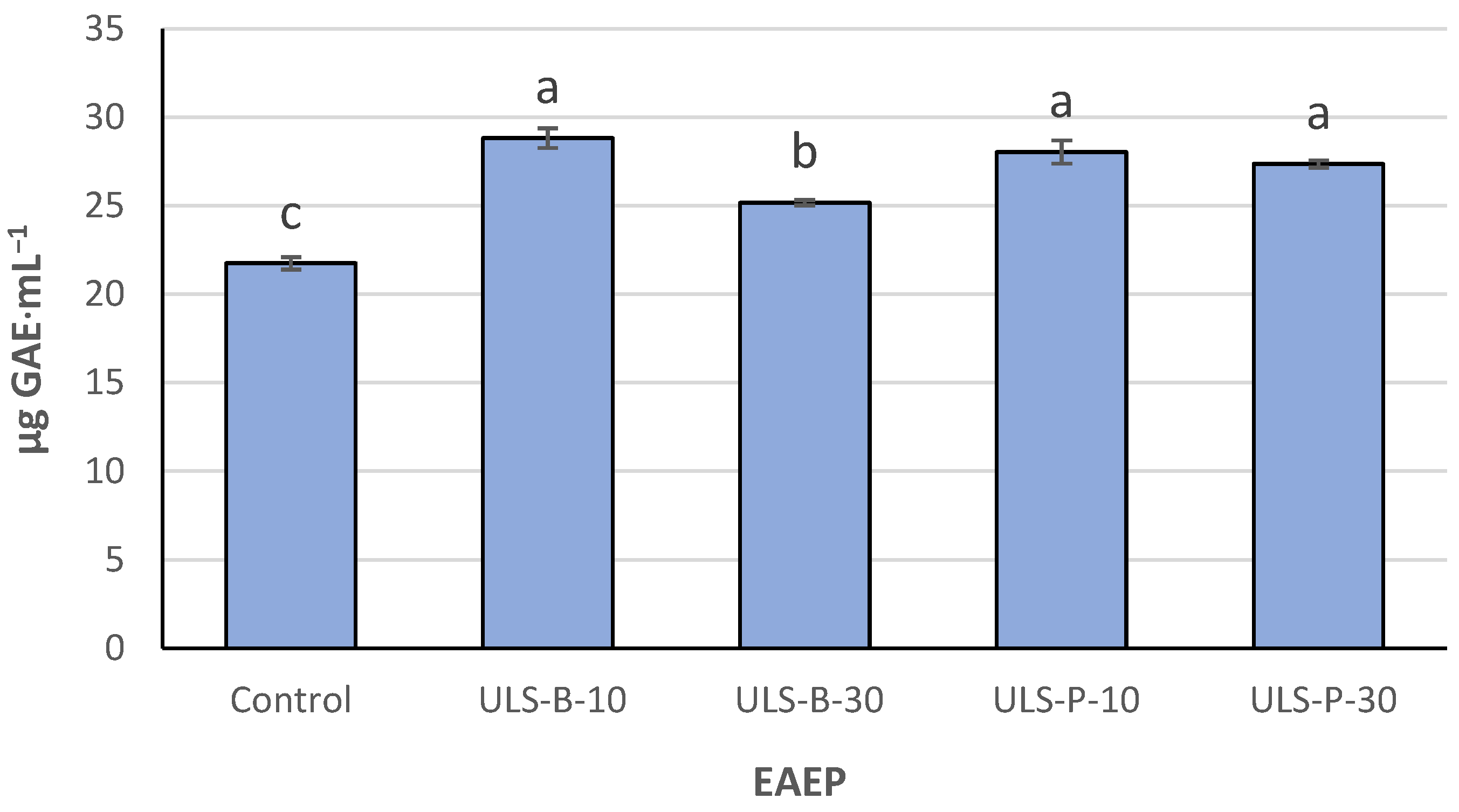
2.2. Total Phenolic Compounds
2.3. Amino Acid Composition
2.4. In Vitro Antioxidant Properties
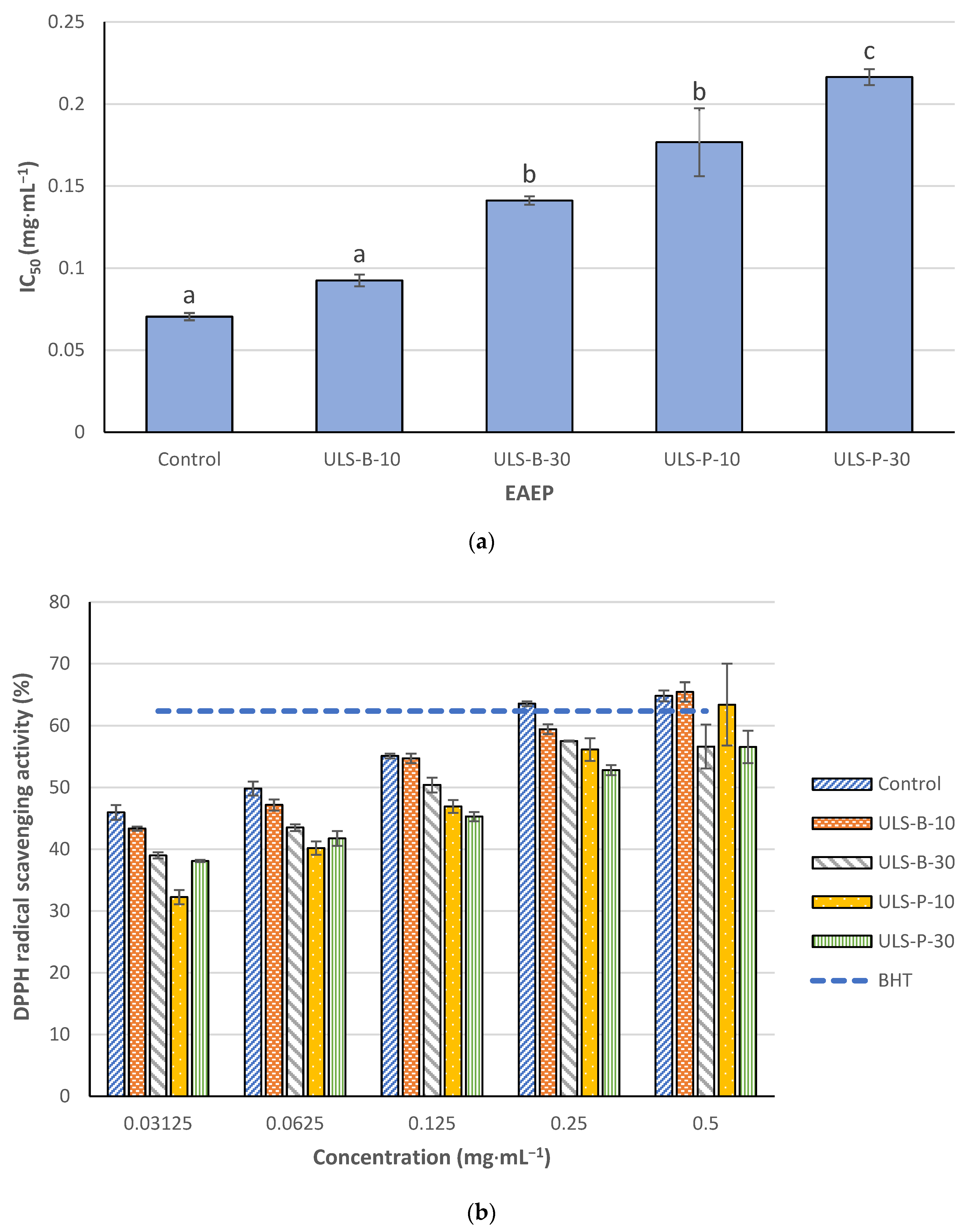
2.4.1. 1,1-Diphenyl-2-picrylhydrazyl (DPPH) Radical Scavenging Activity
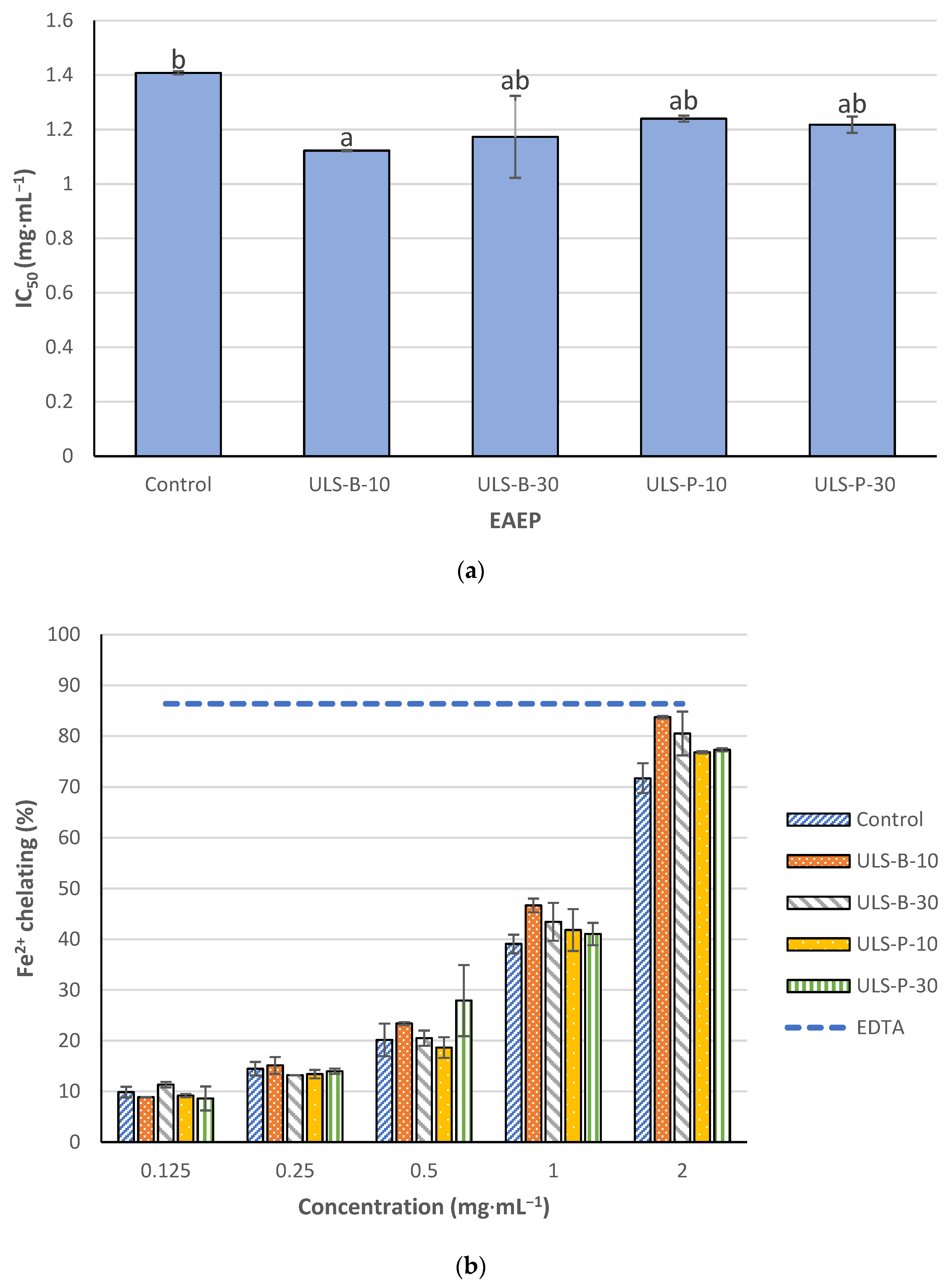
2.4.2. Fe2+ Chelating Activity
2.4.3. In Vitro Antioxidant Activity of EAEP versus Other Types of Extracts
3. Discussion
3.1. Protein Content, Degree of Hydrolysis, Protein Recovery, and Nitrogen-to-Protein Conversion Factor
3.2. Total Phenolic Compounds
3.3. Amino Acid Composition
3.4. In Vitro Antioxidant Properties
4. Materials and Methods
4.1. Seaweed Biomass Preparation
4.2. Enzymes and Chemicals
4.3. Preparation of EAEP
- Control: EAEP without ultrasonication
- ULS-B-10: EAEP ultrasonicated in the bath for 10 min.
- ULS-B-30: EAEP ultrasonicated in the bath for 30 min.
- ULS-P-10: EAEP ultrasonicated using the probe for 10 min.
- ULS-P-30: EAEP ultrasonicated using the probe for 30 min.
4.4. Protein Content and Recovery
4.5. Total Phenolic Content
4.6. Amino Acid Profile
4.7. Nitrogen-to-Protein Conversion Factor
4.8. DPPH Radical Scavenging Activity
4.9. Fe2+ Chelating Activity
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, H.; Zhang, Q.; Zhang, B.; Zhao, Y.; Wang, N. Potential Active Marine Peptides as Anti-Aging Drugs or Drug Candidates. Mar. Drugs 2023, 21, 144. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.M.; Zhang, Z.; Sheng, Y.; Chi, C.F.; Wang, B. A Systematic Review on Marine Umami Peptides: Biological Sources, Preparation Methods, Structure-Umami Relationship, Mechanism of Action and Biological Activities. Food Biosci. 2024, 57, 103637. [Google Scholar] [CrossRef]
- O’connor, J.; Garcia-Vaquero, M.; Meaney, S.; Tiwari, B.K. Bioactive Peptides from Algae: Traditional and Novel Generation Strategies, Structure-Function Relationships, and Bioinformatics as Predictive Tools for Bioactivity. Mar. Drugs 2022, 20, 317. [Google Scholar] [CrossRef] [PubMed]
- Martínez Leo, E.E.; Martín Ortega, A.M.; Segura Campos, M.R. Bioactive Peptides—Impact in Cancer Therapy. In Therapeutic, Probiotic, and Unconventional Foods; Academic Press: Cambridge, MA, USA, 2018; pp. 157–166. [Google Scholar] [CrossRef]
- Hajfathalian, M.; Ghelichi, S.; García-Moreno, P.J.; Moltke Sørensen, A.D.; Jacobsen, C. Peptides: Production, Bioactivity, Functionality, and Applications. Crit. Rev. Food Sci. Nutr. 2018, 58, 3097–3129. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Lloréns, J.L.; Critchley, A.T.; Cornish, M.L.; Mouritsen, O.G. Saved by Seaweeds (II): Traditional Knowledge, Home Remedies, Medicine, Surgery, and Pharmacopoeia. J. Appl. Phycol. 2023, 35, 2049–2068. [Google Scholar] [CrossRef]
- Menaa, F.; Wijesinghe, U.; Thiripuranathar, G.; Althobaiti, N.A.; Albalawi, A.E.; Khan, B.A.; Menaa, B. Marine Algae-Derived Bioactive Compounds: A New Wave of Nanodrugs? Mar. Drugs 2021, 19, 484. [Google Scholar] [CrossRef]
- Dai, N.; Wang, Q.; Xu, B.; Chen, H. Remarkable Natural Biological Resource of Algae for Medical Applications. Front. Mar. Sci. 2022, 9, 912924. [Google Scholar] [CrossRef]
- Dumay, J.; Cognie, B.; Fleurence, J.; Morançais, M.; Turpin, V.; Gavilan, M.C.; Le Strat, Y.; Decottignies, P. Palmaria Species: From Ecology and Cultivation to Its Use in Food and Health Benefits. In Sustainable Global Resources of Seaweeds Volume 1: Bioresources, Cultivation, Trade and Multifarious Applications; Springer International Publishing: Cham, Switzerland, 2022; Volume 1, pp. 45–61. [Google Scholar] [CrossRef]
- Mouritsen, O.G.; Dawczynski, C.; Duelund, L.; Jahreis, G.; Vetter, W.; Schröder, M. On the Human Consumption of the Red Seaweed Dulse (Palmaria palmata (L.) Weber & Mohr). J. Appl. Phycol. 2013, 25, 1777–1791. [Google Scholar] [CrossRef]
- Naseri, A.; Marinho, G.S.; Holdt, S.L.; Bartela, J.M.; Jacobsen, C. Enzyme-Assisted Extraction and Characterization of Protein from Red Seaweed Palmaria palmata. Algal Res. 2020, 47, 101849. [Google Scholar] [CrossRef]
- Harnedy, P.A.; FitzGerald, R.J. In Vitro Assessment of the Cardioprotective, Anti-Diabetic and Antioxidant Potential of Palmaria palmata Protein Hydrolysates. J. Appl. Phycol. 2013, 25, 1793–1803. [Google Scholar] [CrossRef]
- Harnedy-Rothwell, P.A.; McLaughlin, C.M.; Le Gouic, A.V.; Mullen, C.; Parthsarathy, V.; Allsopp, P.J.; McSorley, E.M.; FitzGerald, R.J.; O’Harte, F.P.M. In Vitro and In Vivo Effects of Palmaria palmata Derived Peptides on Glucose Metabolism. Int. J. Pept. Res. Ther. 2021, 27, 1667–1676. [Google Scholar] [CrossRef]
- Yousof, S.M.; Alghamdi, B.S.; Alqurashi, T.; Alam, M.Z.; Tash, R.; Tanvir, I.; Kaddam, L.A.G. Modulation of Gut Microbiome Community Mitigates Multiple Sclerosis in a Mouse Model: The Promising Role of Palmaria palmata Alga as a Prebiotic. Pharmaceuticals 2023, 16, 1355. [Google Scholar] [CrossRef]
- Ghelichi, S.; Sørensen, A.D.M.; García-Moreno, P.J.; Hajfathalian, M.; Jacobsen, C. Physical and Oxidative Stability of Fish Oil-in-Water Emulsions Fortified with Enzymatic Hydrolysates from Common Carp (Cyprinus carpio) Roe. Food Chem. 2017, 237, 1048–1057. [Google Scholar] [CrossRef][Green Version]
- Yesiltas, B.; García-Moreno, P.J.; Gregersen, S.; Olsen, T.H.; Jones, N.C.; Hoffmann, S.V.; Marcatili, P.; Overgaard, M.T.; Hansen, E.B.; Jacobsen, C. Antioxidant Peptides Derived from Potato, Seaweed, Microbial and Spinach Proteins: Oxidative Stability of 5% Fish Oil-in-Water Emulsions. Food Chem. 2022, 385, 132699. [Google Scholar] [CrossRef]
- Sabeena Farvin, K.H.; Jacobsen, C. Phenolic Compounds and Antioxidant Activities of Selected Species of Seaweeds from Danish Coast. Food Chem. 2013, 138, 1670–1681. [Google Scholar] [CrossRef]
- Angell, A.R.; Mata, L.; de Nys, R.; Paul, N.A. The Protein Content of Seaweeds: A Universal Nitrogen-to-Protein Conversion Factor of Five. J. Appl. Phycol. 2016, 28, 511–524. [Google Scholar] [CrossRef]
- Biancarosa, I.; Espe, M.; Bruckner, C.G.; Heesch, S.; Liland, N.; Waagbø, R.; Torstensen, B.; Lock, E.J. Amino Acid Composition, Protein Content, and Nitrogen-to-Protein Conversion Factors of 21 Seaweed Species from Norwegian Waters. J. Appl. Phycol. 2017, 29, 1001–1009. [Google Scholar] [CrossRef]
- Hadidi, M.; Aghababaei, F.; McClements, D.J. Enhanced Alkaline Extraction Techniques for Isolating and Modifying Plant-Based Proteins. Food Hydrocoll. 2023, 145, 109132. [Google Scholar] [CrossRef]
- Rodrigues, D.; Sousa, S.; Silva, A.; Amorim, M.; Pereira, L.; Rocha-Santos, T.A.P.; Gomes, A.M.P.; Duarte, A.C.; Freitas, A.C. Impact of Enzyme- and Ultrasound-Assisted Extraction Methods on Biological Properties of Red, Brown, and Green Seaweeds from the Central West Coast of Portugal. J. Agric. Food Chem. 2015, 63, 3177–3188. [Google Scholar] [CrossRef] [PubMed]
- Echave, J.; Fraga-Corral, M.; Garcia-Perez, P.; Popović-Djordjević, J.; Avdović, E.H.; Radulović, M.; Xiao, J.; Prieto, M.A.; Simal-Gandara, J. Seaweed Protein Hydrolysates and Bioactive Peptides: Extraction, Purification, and Applications. Mar. Drugs 2021, 19, 500. [Google Scholar] [CrossRef] [PubMed]
- Lavilla, I.; Bendicho, C. Fundamentals of Ultrasound-Assisted Extraction. Water Extraction of Bioactive Compounds: From Plants to Drug Development. In Water Extraction of Bioactive Compounds; Elsevier: Amsterdam, The Netherlands, 2017; pp. 291–316. [Google Scholar] [CrossRef]
- Darsih, C.; Indrianingsih, A.W.; Poeloengasih, C.D.; Prasetyo, D.J.; Indirayati, N. In Vitro Antioxidant Activity of Macroalgae Sargassum duplicatum and Palmaria palmata Extracts Collected from Sepanjang Beach, Gunungkidul, Yogyakarta. IOP Conf. Ser. Mater. Sci. Eng. 2021, 1011, 012052. [Google Scholar] [CrossRef]
- Galland-Irmouli, A.V.; Fleurence, J.; Lamghari, R.; Luçon, M.; Rouxel, C.; Barbaroux, O.; Bronowicki, J.P.; Villaume, C.; Guéant, J.L. Nutritional Value of Proteins from Edible Seaweed Palmaria palmata (Dulse). J. Nutr. Biochem. 1999, 10, 353–359. [Google Scholar] [CrossRef]
- Gallagher, J.A.; Adams, J.M.M.; Turner, L.B.; Kirby, M.E.; Toop, T.A.; Mirza, M.W.; Theodorou, M.K. Bio-Processing of Macroalgae Palmaria palmata: Metabolite Fractionation from Pressed Fresh Material and Ensiling Considerations for Long-Term Storage. J. Appl. Phycol. 2021, 33, 533–544. [Google Scholar] [CrossRef]
- Bjarnadóttir, M.; Aðalbjörnsson, B.V.; Nilsson, A.; Slizyte, R.; Roleda, M.Y.; Hreggviðsson, G.Ó.; Friðjónsson, Ó.H.; Jónsdóttir, R. Palmaria palmata as an Alternative Protein Source: Enzymatic Protein Extraction, Amino Acid Composition, and Nitrogen-to-Protein Conversion Factor. J. Appl. Phycol. 2018, 30, 2061–2070. [Google Scholar] [CrossRef]
- Hayes, M. Measuring Protein Content in Food: An Overview of Methods. Foods 2020, 9, 1340. [Google Scholar] [CrossRef]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Siar, E.H.; Tavano, O.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R. Use of Alcalase in the Production of Bioactive Peptides: A Review. Int. J. Biol. Macromol. 2020, 165, 2143–2196. [Google Scholar] [CrossRef]
- Derkach, S.R.; Kuchina, Y.A.; Kolotova, D.S.; Petrova, L.A.; Volchenko, V.I.; Glukharev, A.Y.; Grokhovsky, V.A. Properties of Protein Isolates from Marine Hydrobionts Obtained by Isoelectric Solubilisation/Precipitation: Influence of Temperature and Processing Time. Int. J. Mol. Sci. 2022, 23, 14221. [Google Scholar] [CrossRef]
- Veide Vilg, J.; Undeland, I. pH-Driven Solubilization and Isoelectric Precipitation of Proteins from the Brown Seaweed Saccharina latissima—Effects of Osmotic Shock, Water Volume and Temperature. J. Appl. Phycol. 2017, 29, 585–593. [Google Scholar] [CrossRef]
- Bernardi, S.; Lupatini-Menegotto, A.L.; Kalschne, D.L.; Moraes Flores, É.L.; Bittencourt, P.R.S.; Colla, E.; Canan, C. Ultrasound: A Suitable Technology to Improve the Extraction and Techno-Functional Properties of Vegetable Food Proteins. Plant Foods Hum. Nutr. 2021, 76, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Goncąlves, A.M.M.; Da Silva, G.J.; Pereira, L. Seaweed Phenolics: From Extraction to Applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jónsdóttir, R.; Kristinsson, H.G.; Hreggvidsson, G.O.; Jónsson, J.Ó.; Thorkelsson, G.; Ólafsdóttir, G. Enzyme-Enhanced Extraction of Antioxidant Ingredients from Red Algae Palmaria palmata. LWT Food Sci. Technol. 2010, 43, 1387–1393. [Google Scholar] [CrossRef]
- Holeček, M. Histidine in Health and Disease: Metabolism, Physiological Importance, and Use as a Supplement. Nutrients 2020, 12, 848. [Google Scholar] [CrossRef]
- Mæhre, H.K.; Jensen, I.J.; Eilertsen, K.E. Enzymatic Pre-Treatment Increases the Protein Bioaccessibility and Extractability in Dulse (Palmaria palmata). Mar. Drugs 2016, 14, 196. [Google Scholar] [CrossRef]
- Dong, C.; Zhao, G.; Tao, L.; Qiu, F.; Wang, S.; Wang, B.; Liu, J.; Duan, S. Antioxidant Interactions between S-Allyl-L-Cysteine and Polyphenols Using Interaction Index and Isobolographic Analysis. Molecules 2022, 27, 4089. [Google Scholar] [CrossRef]
- Currell, K. Cysteine and Cystine. In Nutritional Supplements in Sport, Exercise and Health: An A-Z Guide; Routledge: London, UK, 2015; p. 102. [Google Scholar] [CrossRef]
- Fan, C.; Wang, X.; Song, X.; Sun, R.; Liu, R.; Sui, W.; Jin, Y.; Wu, T.; Zhang, M. Identification of a Novel Walnut Iron Chelating Peptide with Potential High Antioxidant Activity and Analysis of Its Possible Binding Sites. Foods 2023, 12, 226. [Google Scholar] [CrossRef]
- Sumampouw, G.A.; Jacobsen, C.; Getachew, A.T. Optimization of Phenolic Antioxidants Extraction from Fucus vesiculosus by Pressurized Liquid Extraction. J. Appl. Phycol. 2021, 33, 1195–1207. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative Properties of Xanthan on the Autoxidation of Soybean Oil in Cyclodextrin Emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Dinis, T.C.P.; Madeira, V.M.C.; Almeida, L.M. Action of Phenolic Derivatives (Acetaminophen, Salicylate, and 5-Aminosalicylate) as Inhibitors of Membrane Lipid Peroxidation and as Peroxyl Radical Scavengers. Arch. Biochem. Biophys. 1994, 315, 161–169. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Dried Seaweed | Control | ULS-B-10 | ULS-B-30 | ULS-P-10 | ULS-P-30 | |
|---|---|---|---|---|---|---|
| Protein (%) | 12.85 ± 0.63 | 11.20 ± 0.16 | 10.96 ± 0.20 | 10.25 ± 0.66 | 10.75 ± 0.15 | 10.67 ± 0.34 |
| DH (%) | - | 28.88 ± 1.34 | 33.19 ± 4.86 | 29.43 ± 0.15 | 25.39 ± 2.41 | 26.86 ± 3.95 |
| PRE * (%) | - | 93.00 ± 0.59 | 93.60 ± 0.66 | 93.91 ± 0.52 | 93.83 ± 1.50 | 91.39 ± 0.17 |
| PRSR ** (%) | - | 3.30 ± 1.65 | 3.99 ± 0.69 | 4.17 ± 1.20 | 4.65 ± 0.00 | 3.97 ± 0.22 |
| Protein (%): NCF 6.25 | Protein (%): NCF 5 | Protein (%): NCF 4 | Protein (%) Based on TAA * | NCF Based on TAA | Protein (%) Based on TAAPC ** | NCF Based on TAAPC | |
|---|---|---|---|---|---|---|---|
| Dried seaweed | 16.05 ± 0.55 a | 12.85 ± 0.63 a | 10.27 ± 0.35 a | 12.34 | 4.80 | 11.68 | 4.54 |
| Control | 14.00 ± 0.14 ab | 11.20 ± 0.16 ab | 8.96 ± 0.09 ab | 10.31 | 4.60 | 8.94 | 3.99 |
| ULS-B-10 | 13.70 ± 0.18 b | 10.96 ± 0.20 b | 8.76 ± 0.11 b | 10.25 | 4.67 | 8.72 | 3.97 |
| ULS-B-30 | 12.80 ± 0.59 b | 10.25 ± 0.66 b | 8.19 ± 0.37 b | 9.67 | 4.71 | 8.23 | 4.01 |
| ULS-P-10 | 13.44 ± 0.13 b | 10.75 ± 0.15 b | 8.60 ± 0.08 b | 10.13 | 4.71 | 8.41 | 3.91 |
| ULS-P-30 | 13.33 ± 0.30 b | 10.67 ± 0.34 b | 8.53 ± 0.19 b | 9.82 | 4.60 | 8.97 | 4.20 |
| Dried Seaweed | Control | ULS-B-10 | ULS-B-30 | ULS-P-10 | ULS-P-30 | |
|---|---|---|---|---|---|---|
| PHE * | 4.61 ± 0.10 | 2.99 ± 0.18 | 3.01 ± 0.07 | 2.79 ± 0.19 | 3.02 ± 0.09 | 2.87 ± 0.16 |
| LEU * | 8.60 ± 0.25 | 5.71 ± 0.41 | 5.86 ± 0.15 | 5.43 ± 0.34 | 5.73 ± 0.03 | 5.70 ± 0.54 |
| ILE * | 3.96 ± 1.48 | 3.65 ± 0.16 | 3.59 ± 0.03 | 3.39 ± 0.25 | 3.57 ± 0.10 | 3.54 ± 0.32 |
| MET * | 2.04 ± 0.10 | 1.61 ± 0.02 | 1.66 ± 0.01 | 1.58 ± 0.13 | 1.58 ± 0.08 | 1.59 ± 0.11 |
| TYR * | 4.05 ± 0.25 | 2.75 ± 0.25 | 2.79 ± 0.05 | 2.65 ± 0.33 | 2.71 ± 0.22 | 2.67 ± 0.11 |
| PRO | 7.19 ± 0.16 | 5.26 ± 0.44 | 5.20 ± 0.20 | 4.99 ± 0.41 | 5.21 ± 0.10 | 5.25 ± 0.41 |
| VAL * | 8.60 ± 0.21 | 6.57 ± 0.57 | 6.30 ± 0.49 | 6.04 ± 0.53 | 6.19 ± 0.24 | 6.39 ± 0.90 |
| ALA | 11.17 ± 0.34 | 8.11 ± 0.01 | 8.25 ± 0.34 | 7.54 ± 0.10 | 8.25 ± 0.12 | 8.14 ± 1.25 |
| THR * | 5.47 ± 0.12 | 4.22 ± 0.56 | 3.70 ± 0.27 | 3.59 ± 0.01 | 3.81 ± 0.08 | 4.00 ± 0.14 |
| GLY | 8.80 ± 0.22 | 6.28 ± 0.87 | 6.10 ± 0.15 | 6.04 ± 0.10 | 6.40 ± 0.56 | 6.48 ± 0.78 |
| SER | 8.08 ± 0.12 | 6.09 ± 0.30 | 6.33 ± 0.09 | 5.47 ± 0.02 | 6.31 ± 0.03 | 5.63 ± 0.52 |
| ARG | 6.83 ± 0.19 | 3.87 ± 0.41 | 3.75 ± 0.17 | 3.65 ± 0.01 | 3.63 ± 0.19 | 3.72 ± 0.82 |
| HIS * | 1.22 ± 0.06 | ND ** | ND | ND | ND | ND |
| LYS * | 4.37 ± 0.19 | 3.36 ± 0.19 | 3.52 ± 0.21 | 3.24 ± 0.05 | 3.28 ± 0.04 | 3.20 ± 0.05 |
| GLU | 19.46 ± 0.46 | 15.39 ± 0.08 | 15.19 ± 0.83 | 14.41 ± 0.67 | 15.47 ± 0.52 | 14.18 ± 1.17 |
| C-C * | 0.51 ± 0.16 | 13.17 ± 1.06 | 13.19 ± 1.22 | 12.49 ± 0.80 | 11.99 ± 0.98 | 11.75 ± 0.09 |
| ASP | 18.44 ± 0.57 | 14.07 ± 0.01 | 14.09 ± 0.30 | 13.42 ± 0.02 | 14.16 ± 0.09 | 13.09 ± 0.76 |
| TAA *** | 123.4 | 103.1 | 102.53 | 96.72 | 101.31 | 98.2 |
| EAA | 43.43 | 44.03 | 43.62 | 41.2 | 41.88 | 41.71 |
| EAA/TAA | 0.351 | 0.427 | 0.425 | 0.425 | 0.413 | 0.424 |
| EAEP | Ethanol Extract [1] | Water Extract [1] | Chloroform Extract [2] | Ethyl Acetate Extract [2] | Methanol Extract [2] | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | ULS-B-10 | ULS-B-30 | ULS-P-10 | ULS-P-30 | ||||||
| DPPH * | 0.07 | 0.09 | 0.14 | 0.17 | 0.21 | 1.16 | 0.57 | >1 | 0.78 | >1 |
| Fe2+ Ch ** | 1.40 | 1.12 | 1.17 | 1.24 | 1.21 | 0.84 | 0.75 | NR *** | NR | NR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghelichi, S.; Sørensen, A.-D.M.; Hajfathalian, M.; Jacobsen, C. Effect of Post-Extraction Ultrasonication on Compositional Features and Antioxidant Activities of Enzymatic/Alkaline Extracts of Palmaria palmata. Mar. Drugs 2024, 22, 179. https://doi.org/10.3390/md22040179
Ghelichi S, Sørensen A-DM, Hajfathalian M, Jacobsen C. Effect of Post-Extraction Ultrasonication on Compositional Features and Antioxidant Activities of Enzymatic/Alkaline Extracts of Palmaria palmata. Marine Drugs. 2024; 22(4):179. https://doi.org/10.3390/md22040179
Chicago/Turabian StyleGhelichi, Sakhi, Ann-Dorit Moltke Sørensen, Mona Hajfathalian, and Charlotte Jacobsen. 2024. "Effect of Post-Extraction Ultrasonication on Compositional Features and Antioxidant Activities of Enzymatic/Alkaline Extracts of Palmaria palmata" Marine Drugs 22, no. 4: 179. https://doi.org/10.3390/md22040179
APA StyleGhelichi, S., Sørensen, A.-D. M., Hajfathalian, M., & Jacobsen, C. (2024). Effect of Post-Extraction Ultrasonication on Compositional Features and Antioxidant Activities of Enzymatic/Alkaline Extracts of Palmaria palmata. Marine Drugs, 22(4), 179. https://doi.org/10.3390/md22040179