

Optimization of the Preparation Process of Glucuronomannan Oligosaccharides and Their Effects on the Gut Microbiota in MPTP-Induced PD Model Mice

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Optimizing the Preparation Process and Preparing GMn
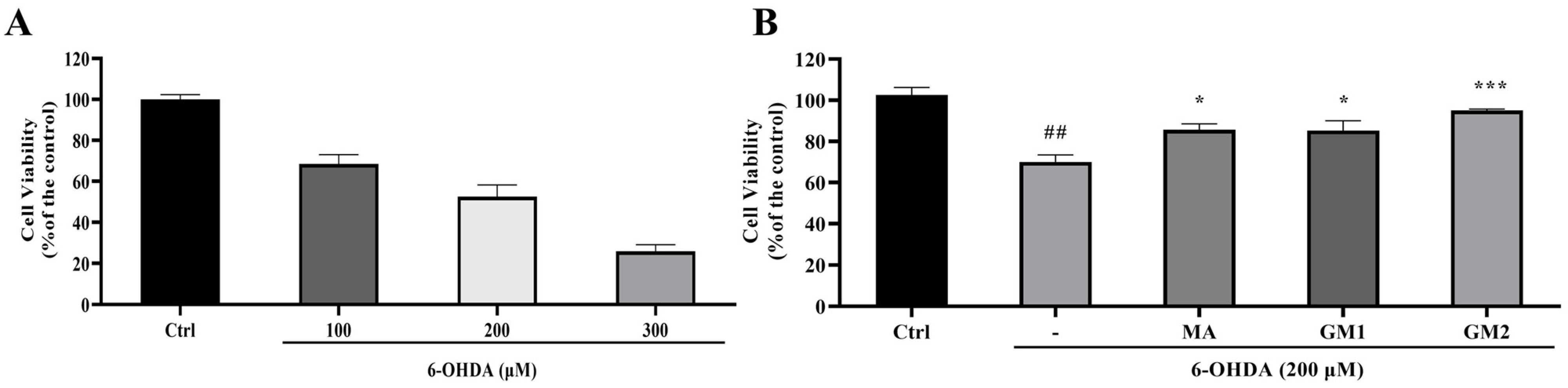
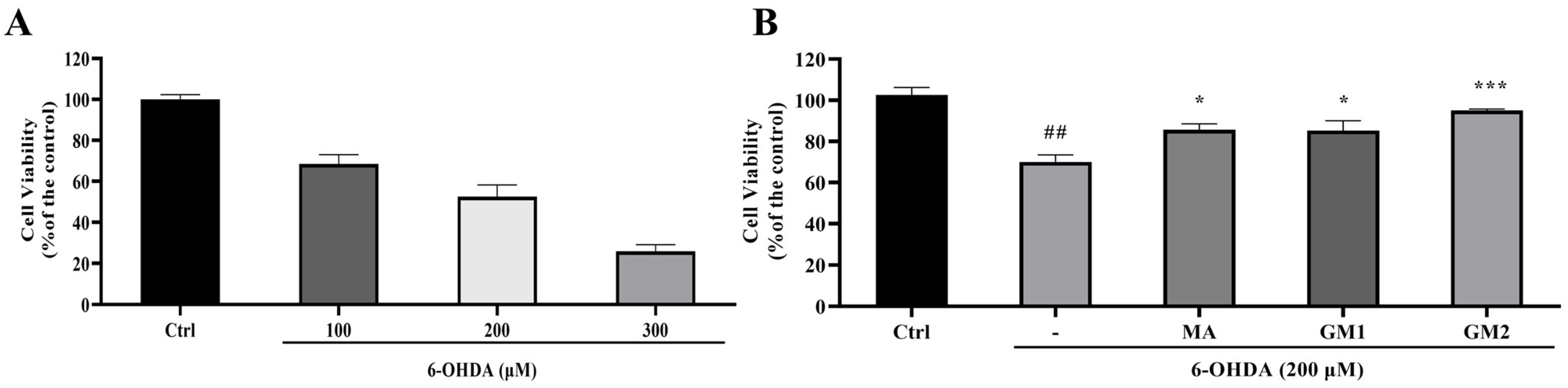
2.2. Protective Effects of GMn on 6-OHDA-Induced Neurotoxicity on SH-SY5Y Cells
2.3. GM2 Improved Motor Behavior in MPTP-Induced PD Mice
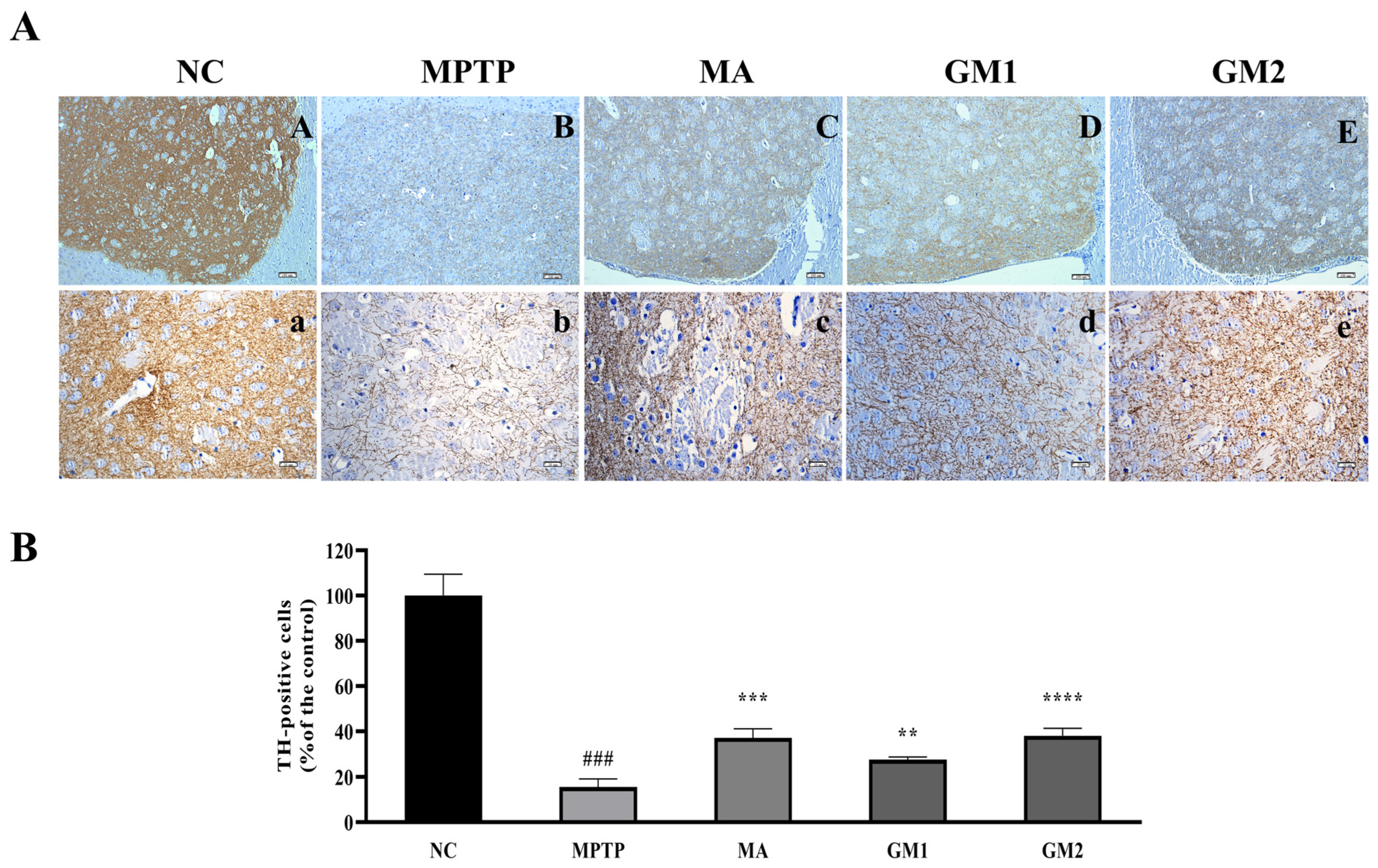
2.4. GMn Attenuated Dopamine Neuronal Damage in MPTP-Induced PD Mice
2.5. GMn Maintained Homeostasis of Monoamine Neurotransmitters
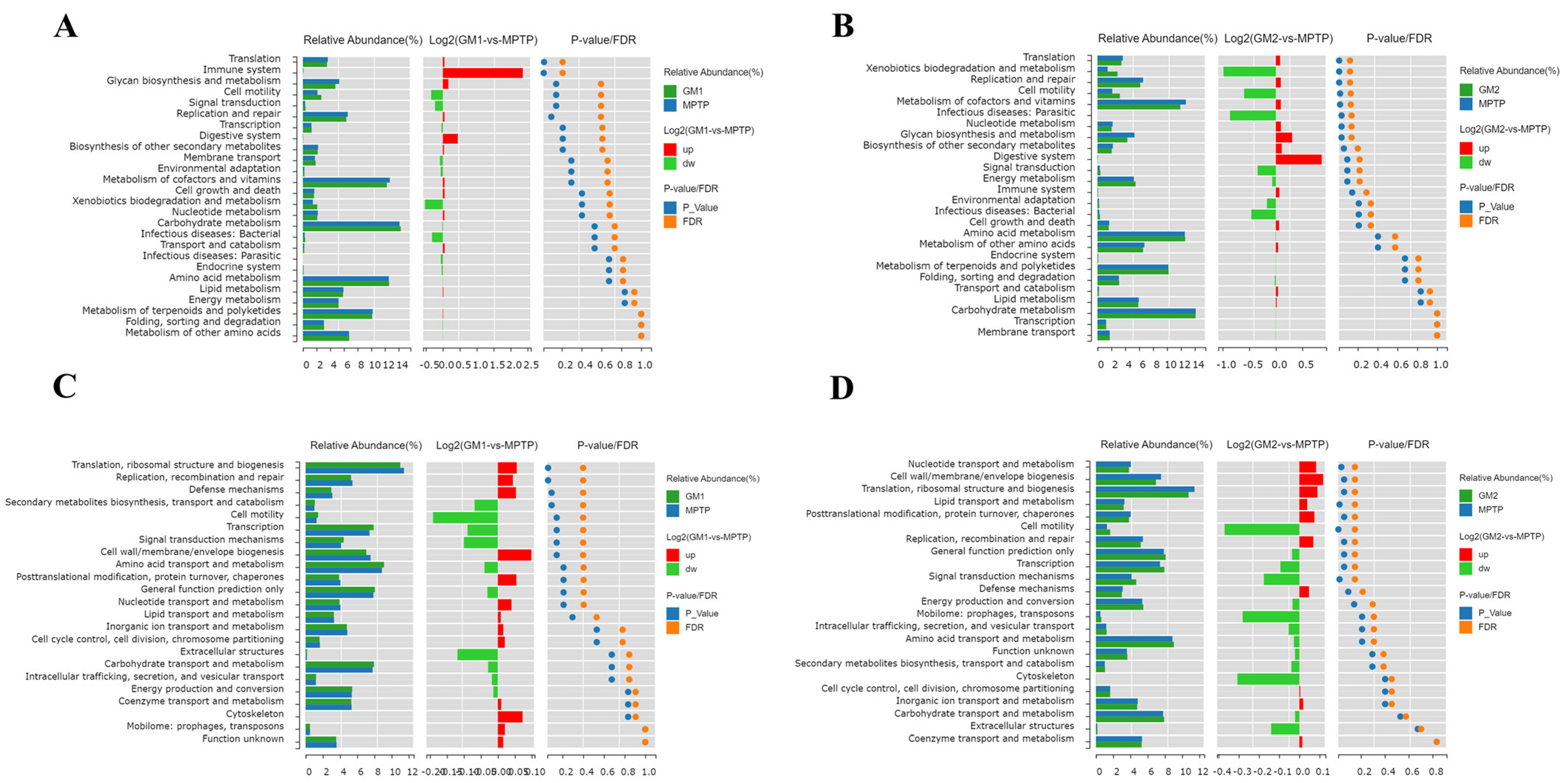
2.6. GMn Alleviated the Dysbiosis of Gut Microbiota in MPTP-Induced PD Model Mice
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
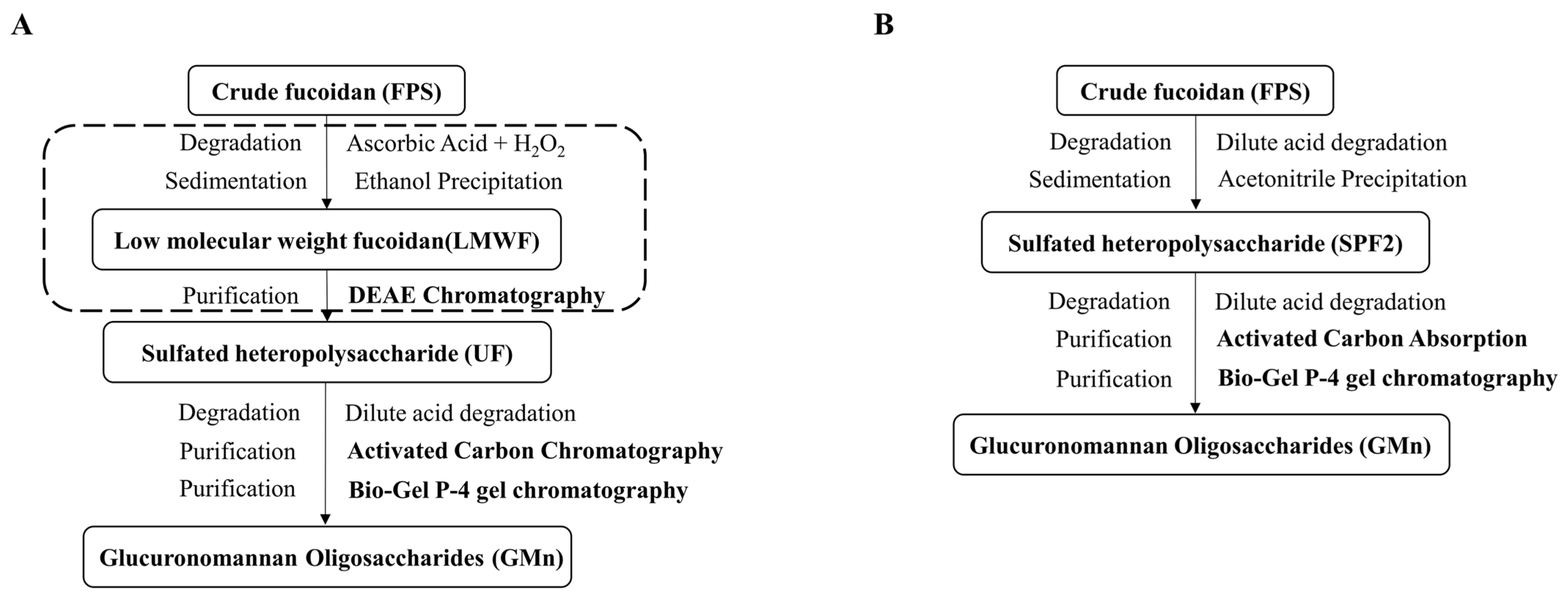
4.2. Preparation of Glucuronomannan Oligosaccharides (GMn)
4.3. Identification and Purity of GMn
4.4. Optimization of Degradation Conditions
4.5. Cell Culture and Treatment
4.6. Cell Viability Assay
4.7. Animals and Treatment
4.8. Open Field Test
4.9. Immunohistochemistry
4.10. HPLC-EDC Analysis
4.11. 16S rRNA Gene Sequencing
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kalia, L.V.; Lang, A.E. Parkinson’s Disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef] [PubMed]
- Aarsland, D.; Batzu, L.; Halliday, G.M.; Geurtsen, G.J.; Ballard, C.; Ray Chaudhuri, K.; Weintraub, D. Parkinson Disease-Associated Cognitive Impairment. Nat. Rev. Dis. Primer 2021, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Connolly, B.S.; Lang, A.E. Pharmacological Treatment of Parkinson Disease: A Review. JAMA 2014, 311, 1670–1683. [Google Scholar] [CrossRef] [PubMed]
- Hussain, G.; Rasul, A.; Anwar, H.; Aziz, N.; Razzaq, A.; Wei, W.; Ali, M.; Li, J.; Li, X. Role of Plant Derived Alkaloids and Their Mechanism in Neurodegenerative Disorders. Int. J. Biol. Sci. 2018, 14, 341–357. [Google Scholar] [CrossRef]
- Zhou, R.; Shi, X.-Y.; Bi, D.-C.; Fang, W.-S.; Wei, G.-B.; Xu, X. Alginate-Derived Oligosaccharide Inhibits Neuroinflammation and Promotes Microglial Phagocytosis of β-Amyloid. Mar. Drugs 2015, 13, 5828–5846. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Mao, J.; Ding, K.; Zhou, Y.; Zeng, X.; Yang, W.; Wang, P.; Zhao, C.; Yao, J.; Xia, P.; et al. Polysaccharides from Ganoderma Lucidum Promote Cognitive Function and Neural Progenitor Proliferation in Mouse Model of Alzheimer’s Disease. Stem Cell Rep. 2017, 8, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-M.; Zhou, N.; Jiang, N.; Lu, K.-M.; Wu, C.-F.; Bao, J.-K. Neuroprotective Effects of Oligosaccharides From Periplaneta Americana on Parkinson’s Disease Models in vitro and in vivo. Front. Pharmacol. 2022, 13, 936818. [Google Scholar] [CrossRef]
- Zhang, L.; Hao, J.; Zheng, Y.; Su, R.; Liao, Y.; Gong, X.; Liu, L.; Wang, X. Fucoidan Protects Dopaminergic Neurons by Enhancing the Mitochondrial Function in a Rotenone-Induced Rat Model of Parkinson’s Disease. Aging Dis. 2018, 9, 590–604. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, X.; Zheng, X.; Sun, S.; Zhao, J.; Long, Y.; Mao, Y. Acidic Oligosaccharide Sugar Chain Combined with Hyperbaric Oxygen Delays D-Galactose-Induced Brain Senescence in Mice via Attenuating Oxidative Stress and Neuroinflammation. Neurosci. Res. 2022, 185, 40–48. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Han, X.; Ma, Y.; Zhang, Z.; Zhao, L.; Guan, F.; Ma, S. Fucoidan: A Promising Agent for Brain Injury and Neurodegenerative Disease Intervention. Food Funct. 2021, 12, 3820–3830. [Google Scholar] [CrossRef]
- Batista, P.; Cunha, S.A.; Ribeiro, T.; Borges, S.; Baptista-Silva, S.; Oliveira-Silva, P.; Pintado, M. Fucoidans: Exploring Its Neuroprotective Mechanisms and Therapeutic Applications in Brain Disorders. Trends Food Sci. Technol. 2024, 143, 104300. [Google Scholar] [CrossRef]
- Luo, D.; Zhang, Q.; Wang, H.; Cui, Y.; Sun, Z.; Yang, J.; Zheng, Y.; Jia, J.; Yu, F.; Wang, X.; et al. Fucoidan Protects against Dopaminergic Neuron Death in Vivo and in Vitro. Eur. J. Pharmacol. 2009, 617, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.-Q.; Jia, Y.-J.; Zhang, T.; Zhang, Q.-B.; Wang, X.-M. Fucoidan Protects against Lipopolysaccharide-Induced Rat Neuronal Damage and Inhibits the Production of Proinflammatory Mediators in Primary Microglia. CNS Neurosci. Ther. 2012, 18, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, H.; Zhang, X.; Li, X.; Geng, L.; Zhang, H.; Zhang, Q. Sulfated Hetero-Polysaccharides Protect SH-SY5Y Cells from H2O2-Induced Apoptosis by Affecting the PI3K/Akt Signaling Pathway. Mar. Drugs 2017, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Wang, J.; Ren, S.; Song, N.; Zhang, Q. Structural Analysis of a Heteropolysaccharide from Saccharina japonica by Electrospray Mass Spectrometry in Tandem with Collision-Induced Dissociation Tandem Mass Spectrometry (ESI-CID-MS/MS). Mar. Drugs 2012, 10, 2138–2152. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jin, W.; Deng, Z.; Wang, J.; Zhang, Q. Preparation and Neuroprotective Activity of Glucuronomannan Oligosaccharides in an MPTP-Induced Parkinson’s Model. Mar. Drugs 2020, 18, 438. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jin, W.; Deng, Z.; Zhang, Q.; Wang, J. Glucuronomannan GM2 from Saccharina japonica Enhanced Mitochondrial Function and Autophagy in a Parkinson’s Model. Mar. Drugs 2021, 19, 58. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.-F.; Shen, Y.-Q. Dysbiosis of Gut Microbiota and Microbial Metabolites in Parkinson’s Disease. Ageing Res. Rev. 2018, 45, 53–61. [Google Scholar] [CrossRef]
- Tan, A.H.; Lim, S.Y.; Lang, A.E. The Microbiome–Gut–Brain Axis in Parkinson Disease—From Basic Research to the Clinic. Nat. Rev. Neurol. 2022, 18, 476–495. [Google Scholar] [CrossRef]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease. Cell 2016, 167, 1469–1480.e12. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, B.; Guo, J. Parkinson’s Disease and Gut Microbiota: From Clinical to Mechanistic and Therapeutic Studies. Transl. Neurodegener. 2023, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Liu, X.; Ye, Y.; Yan, X.; Cheng, Y.; Zhao, L.; Chen, F.; Ling, Z. Gut Microbiota: A Novel Therapeutic Target for Parkinson’s Disease. Front. Immunol. 2022, 13, 937555. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Ren, L.; Liu, B.; Zhang, Q.; Zhong, W. Structural Features of Sulfated Glucuronomannan Oligosaccharides and Their Antioxidant Activity. Mar. Drugs 2018, 16, 291. [Google Scholar] [CrossRef] [PubMed]
- Ferlazzo, N.; Cirmi, S.; Maugeri, A.; Russo, C.; Lombardo, G.E.; Gangemi, S.; Calapai, G.; Mollace, V.; Navarra, M. Neuroprotective Effect of Bergamot Juice in 6-OHDA-Induced SH-SY5Y Cell Death, an in vitro Model of Parkinson’s Disease. Pharmaceutics 2020, 12, 326. [Google Scholar] [CrossRef] [PubMed]
- Franceschelli, S.; Lanuti, P.; Ferrone, A.; Gatta, D.M.P.; Speranza, L.; Pesce, M.; Grilli, A.; Cacciatore, I.; Ricciotti, E.; Di Stefano, A.; et al. Modulation of Apoptotic Cell Death and Neuroprotective Effects of Glutathione—L-Dopa Codrug Against H2O2-Induced Cellular Toxicity. Antioxidants 2019, 8, 319. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Chen, C.; Huang, J.; Wei, H.; Fan, Q. Neuroprotective Effect of Combined Therapy with Hyperbaric Oxygen and Madopar on 6-Hydroxydopamine-Induced Parkinson’s Disease in Rats. Neurosci. Lett. 2015, 600, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Haavik, J.; Toska, K. Tyrosine Hydroxylase and Parkinson’s Disease. Mol. Neurobiol. 1998, 16, 285–309. [Google Scholar] [CrossRef]
- Zhao, J.; Kumar, M.; Sharma, J.; Yuan, Z. Arbutin Effectively Ameliorates the Symptoms of Parkinson’s Disease: The Role of Adenosine Receptors and Cyclic Adenosine Monophosphate. Neural Regen. Res. 2021, 16, 2030. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Xu, S.; Du, N.; Liu, J.; Huang, Y.; Han, M. Neuroprotective Effects of Stemazole in the MPTP-Induced Acute Model of Parkinson’s Disease: Involvement of the Dopamine System. Neurosci. Lett. 2016, 616, 152–159. [Google Scholar] [CrossRef]
- Pajares, M.; Rojo, A.I.; Manda, G.; Boscá, L.; Cuadrado, A. Inflammation in Parkinson’s Disease: Mechanisms and Therapeutic Implications. Cells 2020, 9, 1687. [Google Scholar] [CrossRef]
- Gonçalves, C.; Ferreira, N.; Lourenço, L. Production of Low Molecular Weight Chitosan and Chitooligosaccharides (COS): A Review. Polymers 2021, 13, 2466. [Google Scholar] [CrossRef] [PubMed]
- Xing, R.; Liu, Y.; Li, K.; Yu, H.; Liu, S.; Yang, Y.; Chen, X.; Li, P. Monomer Composition of Chitooligosaccharides Obtained by Different Degradation Methods and Their Effects on Immunomodulatory Activities. Carbohydr. Polym. 2017, 157, 1288–1297. [Google Scholar] [CrossRef] [PubMed]
- Botelho, N.M.; Silveira, E.L.; Lopes, L.N.; Santos, F.A.F.; Teixeira, R.K.C.; Silva, T.T. da Copaiba Oil Effect under Different Pathways in Mice Subjected to Sepsis. Acta Cirúrgica Bras. 2014, 29, 528–531. [Google Scholar] [CrossRef]
- Zhou, Z.-L.; Jia, X.-B.; Sun, M.-F.; Zhu, Y.-L.; Qiao, C.-M.; Zhang, B.-P.; Zhao, L.-P.; Yang, Q.; Cui, C.; Chen, X.; et al. Neuroprotection of Fasting Mimicking Diet on MPTP-Induced Parkinson’s Disease Mice via Gut Microbiota and Metabolites. Neurotherapeutics 2019, 16, 741–760. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wei, J.; Zhao, X.; Liu, Y.; Chen, Z.; Wei, K.; Lu, J.; Chen, W.; Jiang, M.; Li, S.; et al. Neuroprotective Effects of an Engineered Escherichia coli Nissle 1917 on Parkinson’s Disease in Mice by Delivering GLP-1 and Modulating Gut Microbiota. Bioeng. Transl. Med. 2023, 8, e10351. [Google Scholar] [CrossRef] [PubMed]
- Aho, V.T.E.; Pereira, P.A.B.; Voutilainen, S.; Paulin, L.; Pekkonen, E.; Auvinen, P.; Scheperjans, F. Gut Microbiota in Parkinson’s Disease: Temporal Stability and Relations to Disease Progression. EBioMedicine 2019, 44, 691–707. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Zheng, W.; He, Y.; Tang, W.; Wei, X.; He, R.; Huang, W.; Su, Y.; Huang, Y.; Zhou, H.; et al. Gut Microbiota in Patients with Parkinson’s Disease in Southern China. Park. Relat. Disord. 2018, 53, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-J.; Liang, C.-Y.; Yang, L.-Q.; Ren, S.-M.; Xia, Y.-M.; Cui, L.; Li, X.-F.; Gao, B.-L. Association of Parkinson’s Disease With Microbes and Microbiological Therapy. Front. Cell. Infect. Microbiol. 2021, 11, 619354. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Paul, K.C.; Jacobs, J.P.; Chou, H.-C.; Folle, A.D.; Del Rosario, I.; Yu, Y.; Bronstein, J.M.; Keener, A.M.; Ritz, B. Parkinson’s Disease and the Gut Microbiome in Rural California. J. Park. Dis. 2022, 12, 2441–2452. [Google Scholar] [CrossRef] [PubMed]
- Bedarf, J.R.; Hildebrand, F.; Coelho, L.P.; Sunagawa, S.; Bahram, M.; Goeser, F.; Bork, P.; Wüllner, U. Functional Implications of Microbial and Viral Gut Metagenome Changes in Early Stage L-DOPA-Naïve Parkinson’s Disease Patients. Genome Med. 2017, 9, 39. [Google Scholar] [CrossRef]
- Haran, J.P.; Bhattarai, S.K.; Foley, S.E.; Dutta, P.; Ward, D.V.; Bucci, V.; McCormick, B.A. Alzheimer’s Disease Microbiome Is Associated with Dysregulation of the Anti-Inflammatory P-Glycoprotein Pathway. mBio 2019, 10, e00632-19. [Google Scholar] [CrossRef] [PubMed]
- Heintz-Buschart, A.; Pandey, U.; Wicke, T.; Sixel-Döring, F.; Janzen, A.; Sittig-Wiegand, E.; Trenkwalder, C.; Oertel, W.H.; Mollenhauer, B.; Wilmes, P. The Nasal and Gut Microbiome in Parkinson’s Disease and Idiopathic Rapid Eye Movement Sleep Behavior Disorder. Mov. Disord. 2018, 33, 88–98. [Google Scholar] [CrossRef]
- Scheperjans, F.; Aho, V.; Pereira, P.A.B.; Koskinen, K.; Paulin, L.; Pekkonen, E.; Haapaniemi, E.; Kaakkola, S.; Eerola-Rautio, J.; Pohja, M.; et al. Gut Microbiota Are Related to Parkinson’s Disease and Clinical Phenotype. Mov. Disord. 2015, 30, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Bayliss, J.; Stark, R.; Reichenbach, A.; Andrews, Z.B.; Bayliss, J.; Stark, R.; Reichenbach, A.; Andrews, Z.B. Gut Hormones Restrict Neurodegeneration in Parkinson’s Disease. In Advanced Understanding of Neurodegenerative Diseases; IntechOpen: London, UK, 2011; ISBN 978-953-307-529-7. [Google Scholar]
- Minato, T.; Maeda, T.; Fujisawa, Y.; Tsuji, H.; Nomoto, K.; Ohno, K.; Hirayama, M. Progression of Parkinson’s Disease Is Associated with Gut Dysbiosis: Two-Year Follow-up Study. PLoS ONE 2017, 12, e0187307. [Google Scholar] [CrossRef]
- Liu, J.; Lv, X.; Ye, T.; Zhao, M.; Chen, Z.; Zhang, Y.; Yang, W.; Xie, H.; Zhan, L.; Chen, L.; et al. Microbiota-Microglia Crosstalk between Blautia Producta and Neuroinflammation of Parkinson’s Disease: A Bench-to-Bedside Translational Approach. Brain. Behav. Immun. 2024, 117, 270–282. [Google Scholar] [CrossRef]
- Husted, A.S.; Trauelsen, M.; Rudenko, O.; Hjorth, S.A.; Schwartz, T.W. GPCR-Mediated Signaling of Metabolites. Cell Metab. 2017, 25, 777–796. [Google Scholar] [CrossRef]
- Liu, J.; Wang, F.; Liu, S.; Du, J.; Hu, X.; Xiong, J.; Fang, R.; Chen, W.; Sun, J. Sodium Butyrate Exerts Protective Effect against Parkinson’s Disease in Mice via Stimulation of Glucagon like Peptide-1. J. Neurol. Sci. 2017, 381, 176–181. [Google Scholar] [CrossRef]
- Haikal, C.; Chen, Q.-Q.; Li, J.-Y. Microbiome Changes: An Indicator of Parkinson’s Disease? Transl. Neurodegener. 2019, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lu, G.; Li, Z.; Wu, B.; Luo, E.; Qiu, X.; Guo, J.; Xia, Z.; Zheng, C.; Su, Q.; et al. Altered Actinobacteria and Firmicutes Phylum Associated Epitopes in Patients With Parkinson’s Disease. Front. Immunol. 2021, 12, 632482. [Google Scholar] [CrossRef]
- Geng, L.; Zhang, Q.; Wang, J.; Jin, W.; Zhao, T.; Hu, W. Glucofucogalactan, a Heterogeneous Low-Sulfated Polysaccharide from Saccharina japonica and Its Bioactivity. Int. J. Biol. Macromol. 2018, 113, 90–97. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Du, G.-H. Ginsenoside Rg1 Inhibits β-Secretase Activity in vitro and Protects against Aβ-Induced Cytotoxicity in PC12 Cells. J. Asian Nat. Prod. Res. 2009, 11, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, Z.; Abbassian, H.; Sheibani, V.; Haghani, M.; Nazeri, M.; Aghaei, I.; Shabani, M. The Therapeutic Potential of Berberine Chloride Hydrate against Harmaline-Induced Motor Impairments in a Rat Model of Tremor. Neurosci. Lett. 2015, 590, 84–90. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Zhao, S.; Liu, X.; Dong, S.; Lv, J.; Liu, D.; Wang, J.; Meng, Z. ReSeqTools: An Integrated Toolkit for Large-Scale next-Generation Sequencing Based Resequencing Analysis. Genet. Mol. Res. 2013, 12, 6275–6283. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The NO. | A-Hydrolysis Temperature | B-Hydrolysis Time | GM1 Relative Abundance (%) | GM2 Relative Abundance (%) |
|---|---|---|---|---|
| 1 | 1 | 1 | 1.27 | 0.89 |
| 2 | 1 | 2 | 2.23 | 1.4 |
| 3 | 1 | 3 | 1.78 | 1.17 |
| 4 | 2 | 1 | 3.62 | 2.97 |
| 5 | 2 | 2 | 12.36 | 8.85 |
| 6 | 2 | 3 | 7.83 | 6.14 |
| 7 | 3 | 1 | 17.36 | 13.44 |
| 8 | 3 | 2 | 5.37 | 4.15 |
| 9 | 3 | 3 | 7.78 | 6.08 |
| GM1 Relative Abundance (%) | GM2 Relative Abundance (%) | |||
|---|---|---|---|---|
| A-Hydrolysis Temperature | B-Hydrolysis Time | A-Hydrolysis Temperature | B-Hydrolysis Time | |
| K1 | 5.28 | 22.25 | 3.46 | 17.30 |
| K2 | 23.81 | 19.96 | 17.96 | 14.40 |
| K3 | 30.51 | 17.39 | 23.67 | 13.39 |
| k1 | 1.76 | 7.42 | 1.15 | 5.77 |
| k2 | 7.94 | 6.65 | 5.99 | 4.80 |
| k3 | 10.17 | 5.80 | 7.89 | 4.46 |
| R | 8.41 | 1.62 | 6.74 | 1.30 |
| Source | Sum of Squares | DF | Mean Square | F Value | p Value | Significance |
|---|---|---|---|---|---|---|
| GM1 | ||||||
| A | 113.867 | 2 | 56.934 | 3.954 | 0.064 | ns |
| B | 3.941 | 2 | 1.970 | 0.137 | 0.874 | ns |
| Error | 115.180 | 8 | 14.397 | |||
| GM2 | ||||||
| A | 72.366 | 2 | 36.183 | 4.611 | 0.047 | * |
| B | 2.746 | 2 | 1.373 | 0.175 | 0.843 | ns |
| Error | 62.773 | 8 | 7.847 |
| Groups | DA | DOPAC | HVA | 5-HIAA | 5-HT | (DOPAC + HVA)/DA |
|---|---|---|---|---|---|---|
| NC | 0.511 ± 0.224 | 0.795 ± 0.116 | 0.983 ± 0.131 | 0.686 ± 0.101 | 0.042 ± 0.014 | 4.82 |
| MPTP | 0.161 ± 0.035 # | 0.294 ± 0.029 # | 0.701 ± 0.075 # | 0.586 ± 0.044 | 0.019 ± 0.001 | 7.55 |
| GM1 | 0.417 ± 0.065 | 0.579 ± 0.042 * | 0.971 ± 0.076 * | 0.560 ± 0.033 | 0.027 ± 0.007 | 3.84 * |
| GM2 | 0.323 ± 0.101 | 0.589 ± 0.060 * | 1.103 ± 0.052 * | 0.687 ± 0.015 | 0.024 ± 0.003 | 5.31 |
| Factors | Levels | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| A-hydrolysis temperature (°C) | 90 | 95 | 100 |
| B-hydrolysis time (h) | 3.5 | 4 | 4.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; Geng, L.; Wang, J.; Wei, Y.; Yan, C.; Wu, N.; Yue, Y.; Zhang, Q. Optimization of the Preparation Process of Glucuronomannan Oligosaccharides and Their Effects on the Gut Microbiota in MPTP-Induced PD Model Mice. Mar. Drugs 2024, 22, 193. https://doi.org/10.3390/md22050193
Wang B, Geng L, Wang J, Wei Y, Yan C, Wu N, Yue Y, Zhang Q. Optimization of the Preparation Process of Glucuronomannan Oligosaccharides and Their Effects on the Gut Microbiota in MPTP-Induced PD Model Mice. Marine Drugs. 2024; 22(5):193. https://doi.org/10.3390/md22050193
Chicago/Turabian StyleWang, Baoxiang, Lihua Geng, Jing Wang, Yuxi Wei, Changhui Yan, Ning Wu, Yang Yue, and Quanbin Zhang. 2024. "Optimization of the Preparation Process of Glucuronomannan Oligosaccharides and Their Effects on the Gut Microbiota in MPTP-Induced PD Model Mice" Marine Drugs 22, no. 5: 193. https://doi.org/10.3390/md22050193
APA StyleWang, B., Geng, L., Wang, J., Wei, Y., Yan, C., Wu, N., Yue, Y., & Zhang, Q. (2024). Optimization of the Preparation Process of Glucuronomannan Oligosaccharides and Their Effects on the Gut Microbiota in MPTP-Induced PD Model Mice. Marine Drugs, 22(5), 193. https://doi.org/10.3390/md22050193