
Avellanin A Has an Antiproliferative Effect on TP-Induced RWPE-1 Cells via the PI3K-Akt Signalling Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
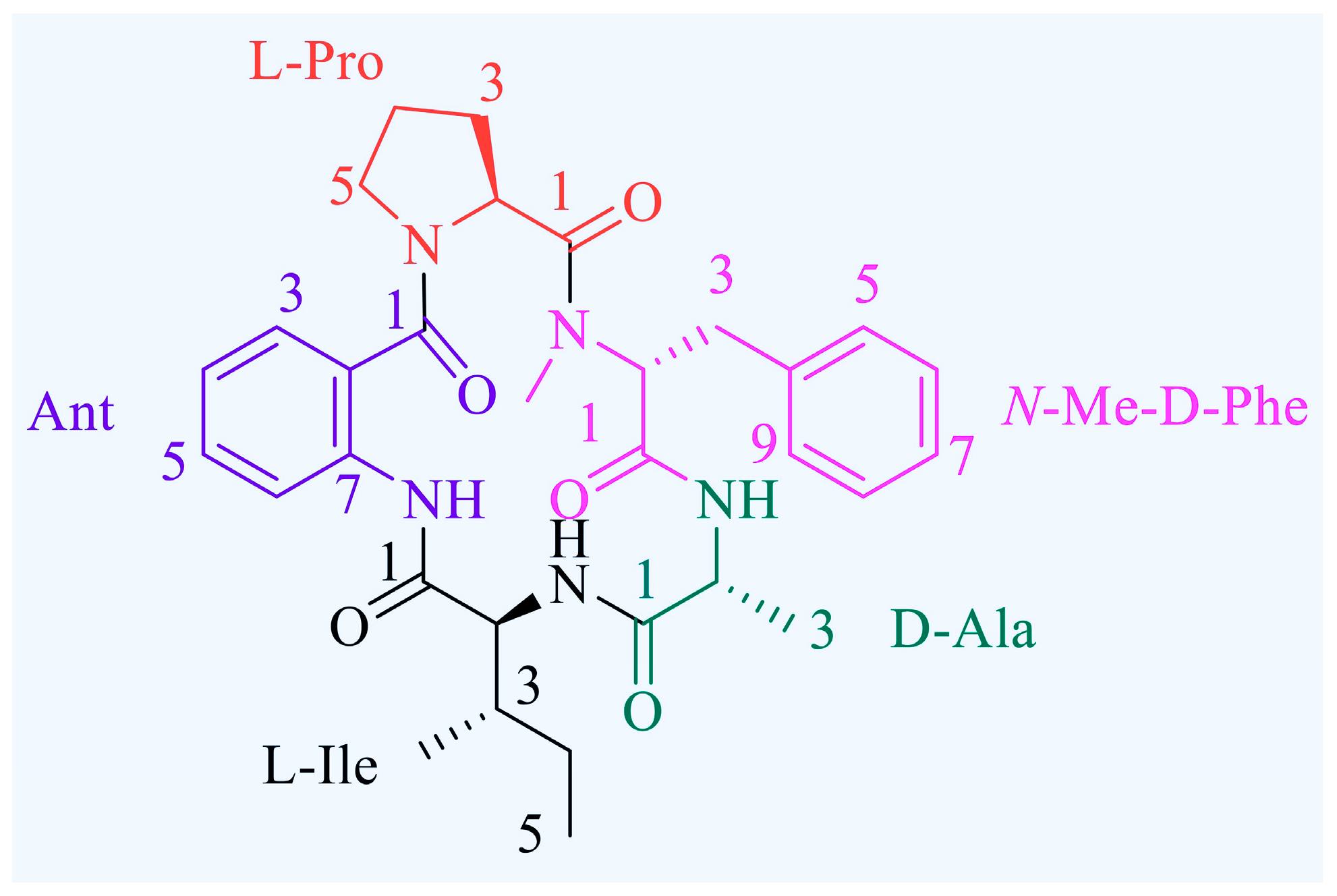
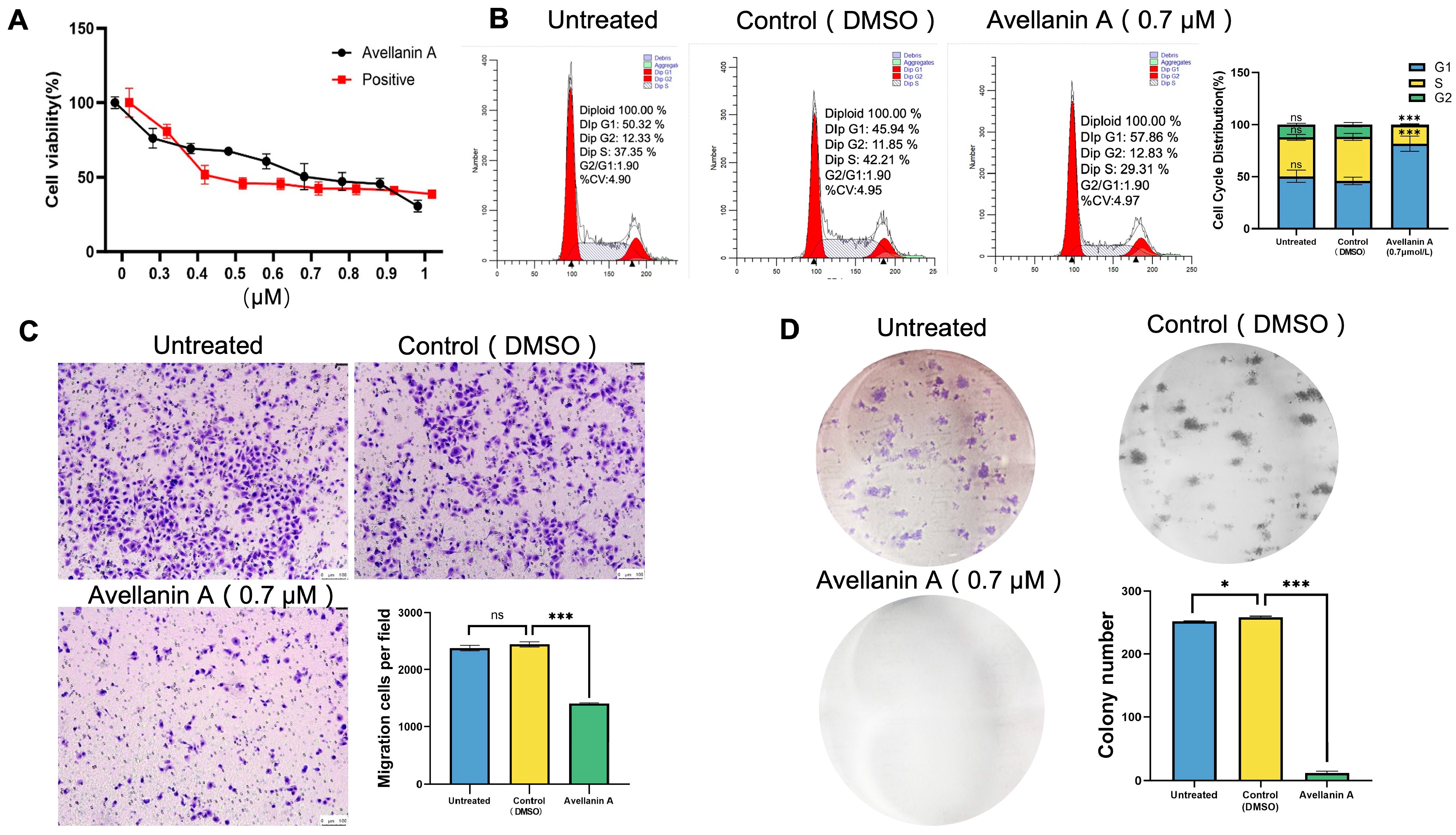
2.1. Antiproliferative Efficacy of Avellanin A on RWPE-1 Cells
2.2. Transcriptome Analysis of RWPE-1 Cells Treated with Avellanin A
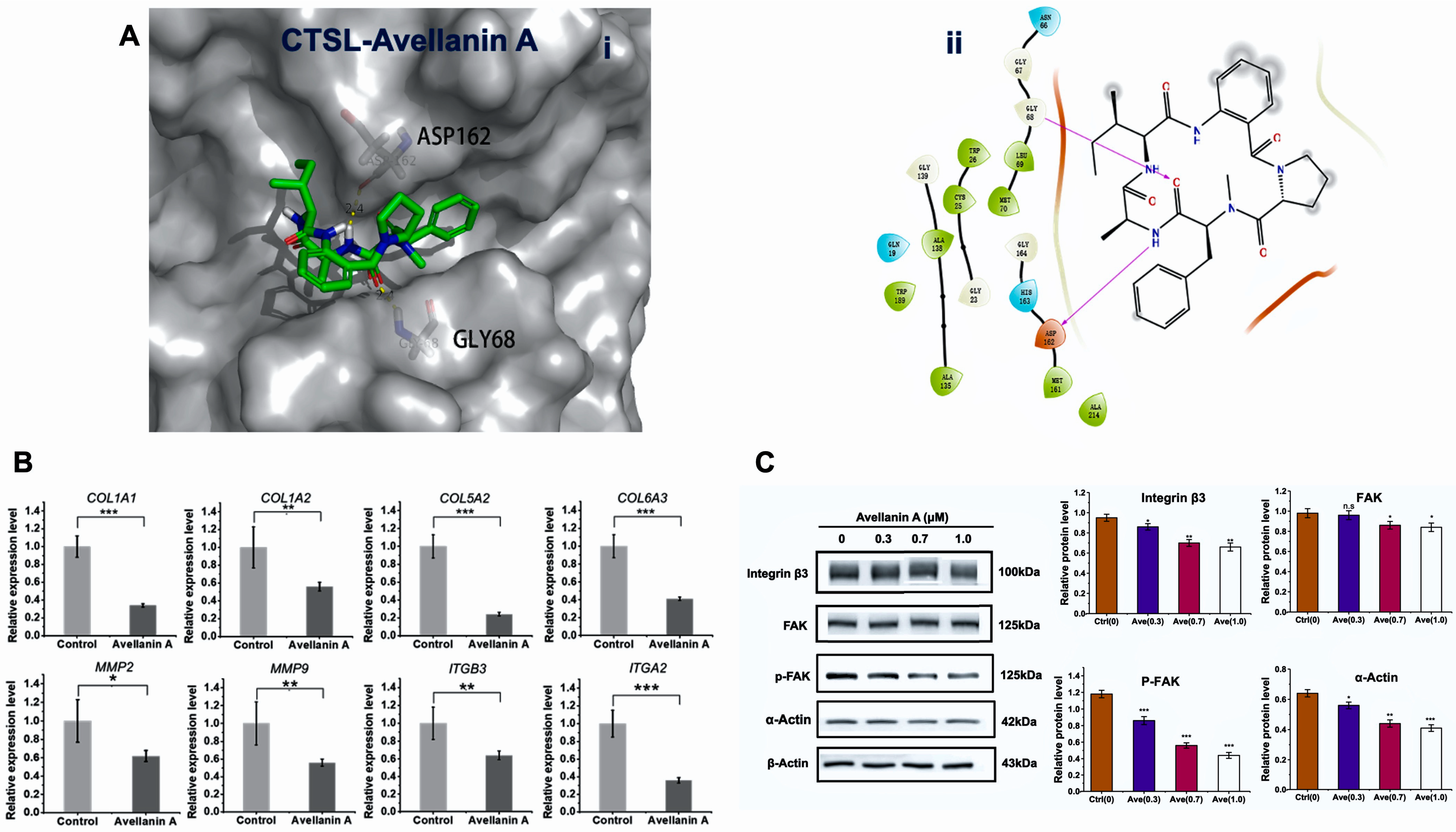
2.3. Interaction of Avellanin A with the Cathepsin L
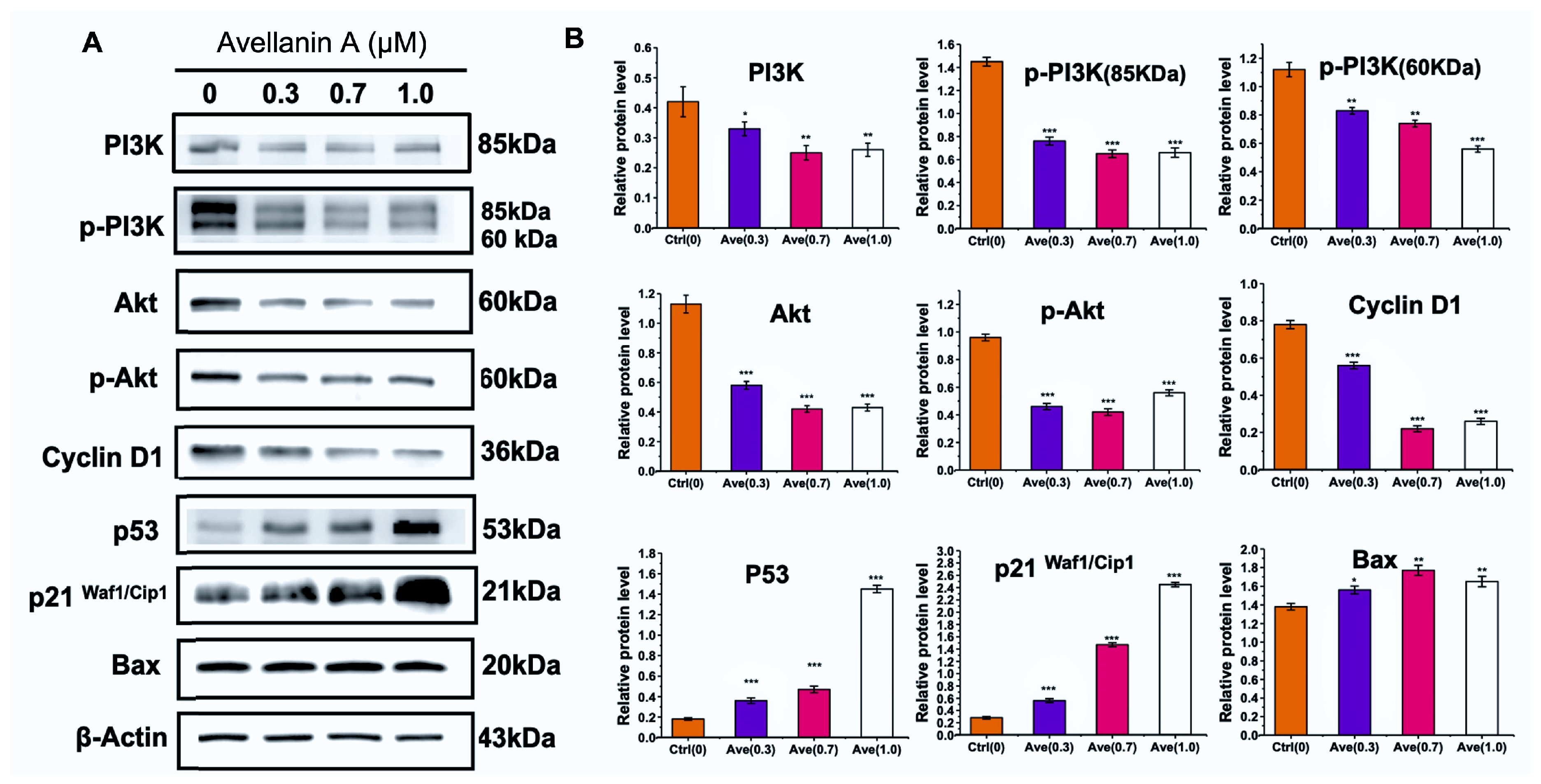
2.4. Avellanin A Suppressed the PI3K-Akt Pathway in RWPE-1 Cells
3. Discussion
4. Materials and Methods
4.1. Fungal Material
4.2. Isolation and Purification
4.3. Cell Culture and Reagents
4.4. Cell Viability Assays
4.5. Cell Cycle Assay
4.6. Cell Migration Assays
4.7. Colony Formation Assay
4.8. Transcriptomic Expression Profiling and Bioinformatics Analysis
4.9. Molecular Docking
4.10. Fluorescence-Based Quantitative PCR
4.11. Western Blotting
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, S.W.H.; Chan, E.M.C.; Lai, Y.K. The global burden of lower urinary tract symptoms suggestive of benign prostatic hyperplasia: A systematic review and meta-analysis. Sci. Rep. 2017, 7, 7984. [Google Scholar] [CrossRef]
- Shah, A.; Shah, A.A.; Nandakumar, K.; Lobo, R. Mechanistic targets for BPH and prostate cancer-a review. Rev. Environ. Health 2021, 36, 261–270. [Google Scholar] [CrossRef]
- Lokeshwar, S.D.; Harper, B.T.; Webb, E.; Jordan, A.; Dykes, T.A.; Neal, D.E.; Terris, M.K.; Klaassen, Z. Epidemiology and treatment modalities for the management of benign prostatic hyperplasia. Transl. Androl. Urol. 2019, 8, 529–539. [Google Scholar] [CrossRef]
- Dhanabalan, A.K.; Kumar, P.; Vasudevan, S.; Chworos, A.; Velmurugan, D. Identification of a novel drug molecule for neurodegenerative disease from marine algae through in-silico analysis. J. Biomol. Struct. Dyn. 2024, 1–10. [Google Scholar] [CrossRef]
- Xu, M.W.; Bai, Z.F.; Xie, B.C.; Peng, R.; Du, Z.W.; Liu, Y.; Zhang, G.S.; Yan, S.; Xiao, X.H.; Qin, S.L. Marine-Derived Bisindoles for Potent Selective Cancer Drug Discovery and Development. Molecules 2024, 29, 933. [Google Scholar] [CrossRef]
- Kallifidas, D.; Dhakal, D.; Chen, M.Y.; Chen, Q.Y.; Kokkaliari, S.; Rosa, N.A.C.; Ratnayake, R.; Bruner, S.D.; Paul, V.J.; Ding, Y.S.; et al. Biosynthesis of Dolastatin 10 in Marine Cyanobacteria, a Prototype for Multiple Approved Cancer Drugs. Org. Lett. 2024, 26, 1321–1325. [Google Scholar] [CrossRef]
- Marunganathan, V.; Kumar, M.S.K.; Kari, Z.A.; Giri, J.; Shaik, M.R.; Shaik, B.; Guru, A. Marine-derived κ-carrageenan-coated zinc oxide nanoparticles for targeted drug delivery and apoptosis induction in oral cancer. Mol. Biol. Rep. 2024, 51, 89. [Google Scholar] [CrossRef]
- Akram, W.; Rihan, M.; Ahmed, S.; Arora, S.; Ahmad, S.; Vashishth, R. Marine-Derived Compounds Applied in Cardiovascular Diseases: Submerged Medicinal Industry. Mar. Drugs 2023, 21, 193. [Google Scholar] [CrossRef]
- Wang, J.M.; Qin, Y.N.; Lin, M.P.; Song, Y.Y.; Lu, H.M.; Xu, X.Y.; Liu, Y.H.; Zhou, X.F.; Gao, C.H.; Luo, X.W. Marine Natural Products from the Beibu Gulf: Sources, Chemistry, and Bioactivities. Mar. Drugs 2023, 21, 63. [Google Scholar] [CrossRef]
- Wang, Z.M.; Qader, M.; Wang, Y.F.; Kong, F.D.; Wang, Q.; Wang, C. Progress in the discovery of new bioactive substances from deep-sea associated fungi during 2020–2022. Front. Mar. Sci. 2023, 10, 1232891. [Google Scholar] [CrossRef]
- Wang, S.C.; Fan, L.M.; Pan, H.Y.; Li, Y.Y.; Qiu, Y.; Lu, Y.M. Antimicrobial peptides from marine animals: Sources, structures, mechanisms and the potential for drug development. Front. Mar. Sci. 2023, 9, 1112595. [Google Scholar] [CrossRef]
- Ghoran, S.H.; Taktaz, F.; Sousa, E.; Fernandes, C.; Kijjoa, A. Peptides from Marine-Derived Fungi: Chemistry and Biological Activities. Mar. Drugs 2023, 21, 510. [Google Scholar] [CrossRef]
- Yan, J.X.; Wu, Q.H.; Maity, M.; Braun, D.R.; Alas, I.; Wang, X.; Yin, X.; Zhu, Y.L.; Bell, B.A.; Rajski, S.R.; et al. Rapid Unambiguous Structure Elucidation of Streptnatamide A, a New Cyclic Peptide Isolated from A Marine-derived Streptomyces sp. Chem.—A Eur. J. 2023, 29, e202301813. [Google Scholar] [CrossRef]
- Yamazaki, M.; Horie, Y.; Bae, K.; Maebayashi, Y.; Jisai, Y.; Fujimoto, H. New fungal metabolites avellanins a and B from Hamigers avellanea with pressor effect. Chem. Pharm. Bull. 1987, 35, 2122–2124. [Google Scholar] [CrossRef]
- Honda, M.; Inagaki, M.; Masuda, Y. Total synthesis of the cyclic pentapeptides PF1171B, D, E, and avellanins A, B, C with inhibitory activity against apolipoprotein B production. Tetrahedron Lett. 2021, 81, 153340. [Google Scholar] [CrossRef]
- Ismail, E.T.; El-Son, M.A.M.; El-Gohary, F.A.; Zahran, E. Prevalence, genetic diversity, and antimicrobial susceptibility of Vibrio spp. infected gilthead sea breams from coastal farms at Damietta, Egypt. BMC Vet. Res. 2024, 20, 129. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Grkovic, T.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2024, 41, 162–207. [Google Scholar] [CrossRef]
- Poreba, M.; Rut, W.; Vizovisek, M.; Groborz, K.; Kasperkiewicz, P.; Finlay, D.; Vuori, K.; Turk, D.; Turk, B.; Salvesen, G.S.; et al. Selective imaging of cathepsin L in breast cancer by fluorescent activity-based probes. Chem. Sci. 2018, 9, 2113–2129. [Google Scholar] [CrossRef]
- Ueki, N.; Wang, W.; Swenson, C.; McNaughton, C.; Sampson, N.S.; Hayman, M.J. Synthesis and Preclinical Evaluation of a Highly Improved Anticancer Prodrug Activated by Histone Deacetylases and Cathepsin L. Theranostics 2016, 6, 808–816. [Google Scholar] [CrossRef]
- Sudhan, D.R.; Siemann, D.W. Cathepsin L targeting in cancer treatment. Pharmacol. Therapeut 2015, 155, 105–116. [Google Scholar] [CrossRef]
- Qian, F.; Xu, H.Y.; Zhang, Y.K.; Li, L.F.; Yu, R.T. Methionine deprivation inhibits glioma growth through downregulation of CTSL. Am. J. Cancer Res. 2022, 12, 5004–5018. [Google Scholar] [PubMed]
- Zhang, L.M.; Zhao, Y.; Yang, J.; Zhu, Y.N.; Li, T.; Liu, X.Y.; Zhang, P.F.; Cheng, J.L.; Sun, S.; Wei, C.L.; et al. CTSL, a prognostic marker of breast cancer, that promotes proliferation, migration, and invasion in cells in triple-negative breast cancer. Front. Oncol. 2023, 13, 1158087. [Google Scholar] [CrossRef]
- Goulet, B.; Sansregret, L.; Leduy, L.; Bogyo, M.; Weber, E.; Chauhan, S.S.; Nepveu, A. Increased expression and activity of nuclear cathepsin L in cancer cells suggests a novel mechanism of cell transformation. Mol. Cancer Res. 2007, 5, 899–907. [Google Scholar] [CrossRef]
- Zhang, C.P.; Yu, Z.Y.; Yang, S.S.; Liu, Y.T.; Song, J.N.; Mao, J.; Li, M.H.; Zhao, Y. ZNF460-mediated circRPPH1 promotes TNBC progression through ITGA5-induced FAK/PI3K/AKT activation in a ceRNA manner. Mol. Cancer 2024, 23, 33. [Google Scholar] [CrossRef] [PubMed]
- Ke, Z.B.; Cai, H.; Wu, Y.P.; Lin, Y.Z.; Li, X.D.; Huang, J.B.; Sun, X.L.; Zheng, Q.S.; Xue, X.Y.; Wei, Y.; et al. Identification of key genes and pathways in benign prostatic hyperplasia. J. Cell Physiol. 2019, 234, 19942–19950. [Google Scholar] [CrossRef]
- El-Shafei, N.H.; Zaafan, M.A.; Kandil, E.A.; Sayed, R.H. Simvastatin ameliorates testosterone-induced prostatic hyperplasia in rats via modulating IGF-1/PI3K/AKT/FOXO signaling. Eur. J. Pharmacol. 2023, 950, 175762. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.J.; Zhang, M.Y.; Hu, S.; Zhang, C.Y.; Zhou, Y.; Han, M.; Li, J.J.; Li, F.L.; Ni, H.M.; Fang, S.Q.; et al. MiRNA-766-3p inhibits gastric cancer via targeting COL1A1 and regulating PI3K/AKT signaling pathway. J. Cancer 2024, 15, 990–998. [Google Scholar] [CrossRef]
- Li, A.A.; Wang, S.J.; Nie, J.B.; Xiao, S.N.; Xie, X.S.; Zhang, Y.; Tong, W.L.; Yao, G.L.; Liu, N.; Dan, F.; et al. USP3 promotes osteosarcoma progression via deubiquitinating EPHA2 and activating the PI3K/AKT signaling pathway. Cell Death Dis. 2024, 15, 235. [Google Scholar] [CrossRef]
- Choi, Y.J.; Fan, M.; Wedamulla, N.E.; Tang, Y.J.; Kim, E.K. Alleviatory effect of isoquercetin on benign prostatic hyperplasia via IGF-1/PI3K/Akt/mTOR pathway. Food Sci. Hum. Well 2024, 13, 1698–1710. [Google Scholar] [CrossRef]
- Yu, H.C.; Jeon, Y.G.; Na, A.Y.; Han, C.Y.; Lee, M.R.; Yang, J.D.; Yu, H.C.; Son, J.B.; Kim, N.D.; Kim, J.B.; et al. p21-activated kinase 4 counteracts PKA-dependent lipolysis by phosphorylating FABP4 and HSL. Nat. Metab. 2024, 6, 94–112. [Google Scholar] [CrossRef]
- Ben-Oz, B.M.; Machour, F.E.; Nicola, M.; Argoetti, A.; Polyak, G.; Hanna, R.; Kleifeld, O.; Mandel-Gutfreund, Y.; Ayoub, N. A dual role of RBM42 in modulating splicing and translation of CDKN1A/p21 during DNA damage response. Nat. Commun. 2023, 14, 7628. [Google Scholar] [CrossRef]
- Ma, C.Y.; Wang, D.D.; Tian, Z.F.; Gao, W.R.; Zang, Y.C.; Qian, L.L.; Xu, X.; Jia, J.H.; Liu, Z.F. USP13 deubiquitinates and stabilizes cyclin D1 to promote gastric cancer cell cycle progression and cell proliferation. Oncogene 2023, 42, 2249–2262. [Google Scholar] [CrossRef]
- Wang, J.; Su, W.; Zhang, T.T.; Zhang, S.S.; Lei, H.W.; Ma, F.D.; Shi, M.N.; Shi, W.J.; Xie, X.D.; Di, C.X. Aberrant Cyclin D1 splicing in cancer: From molecular mechanism to therapeutic modulation. Cell Death Dis. 2023, 14, 244. [Google Scholar] [CrossRef]
- Chen, J.; Wei, J.Q.; Hong, M.N.; Zhang, Z.; Zhou, H.D.; Lu, Y.Y.; Zhang, J.; Guo, Y.T.; Chen, X.; Wang, J.G.; et al. Mitogen-Activated Protein Kinases Mediate Adventitial Fibroblast Activation and Neointima Formation via GATA4/Cyclin D1 Axis. Cardiovasc. Drug Ther. 2024, 38, 215–222. [Google Scholar] [CrossRef]
- Zhao, J.L.; Wu, Y.; Xiao, T.; Cheng, C.; Zhang, T.; Gao, Z.Y.; Hu, S.Y.; Ren, Z.; Yu, X.Z.; Yang, F.; et al. A specific anti-cyclin D1 intrabody represses breast cancer cell proliferation by interrupting the cyclin D1-CDK4 interaction. Breast Cancer Res. Tr. 2023, 198, 555–568. [Google Scholar] [CrossRef]
- Fang, M.; Wu, H.K.; Pei, Y.M.; Zhang, Y.; Gao, X.Y.; He, Y.Y.; Chen, G.J.; Lv, F.X.; Jiang, P.; Li, Y.M.; et al. E3 ligase MG53 suppresses tumor growth by degrading cyclin D1. Signal Transduct. Tar. 2023, 8, 263. [Google Scholar] [CrossRef]
- Rahimi, E.; Asefi, F.; Afzalinia, A.; Khezri, S.; Zare-Zardini, H.; Ghorani-Azam, A.; Es-haghi, A.; Yazdi, M.E.T. Chitosan coated copper/silver oxide nanoparticles as carriers of breast anticancer drug: Cyclin D1/P53 expressions and cytotoxicity studies. Inorg. Chem. Commun. 2023, 158, 111581. [Google Scholar] [CrossRef]
- Li, S.Z.; Xue, J.C.; Zhang, H.; Shang, G.N. ARHGAP44-mediated regulation of the p53/C-myc/Cyclin D1 pathway in modulating the malignant biological behavior of osteosarcoma cells. J. Orthop. Surg. Res. 2023, 18, 972. [Google Scholar] [CrossRef]
- Gutu, N.; Binish, N.; Keilholz, U.; Herzel, H.; Granada, A.E. p53 and p21 dynamics encode single-cell DNA damage levels, fine-tuning proliferation and shaping population heterogeneity. Commun. Biol. 2023, 6, 1196. [Google Scholar] [CrossRef]
- Mchenry, M.W.; Shi, P.W.; Camara, C.M.; Cohen, D.T.; Rettenmaier, T.J.; Adhikary, U.; Gygi, M.A.; Yang, K.; Gygi, S.P.; Wales, T.E.; et al. Covalent inhibition of pro-apoptotic BAX. Nat. Chem. Biol. 2024, 1–11. [Google Scholar] [CrossRef]
- Shen, L.H.; Fan, L.; Luo, H.; Li, W.Y.; Cao, S.Z.; Yu, S.M. Cow placenta extract ameliorates d-galactose-induced liver damage by regulating BAX/CASP3 and p53/p21/p16 pathways. J. Ethnopharmacol. 2024, 323, 117685. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, M.; Osbron, C.A.; Koehler, H.S.; Goodman, A.G. STING dependent BAX-IRF3 signaling results in apoptosis during late-stage infection. Cell Death Dis. 2024, 15, 195. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Cao, G.; Zhang, H.; Bai, M.; Yi, X.; Qu, X. Avellanin A Has an Antiproliferative Effect on TP-Induced RWPE-1 Cells via the PI3K-Akt Signalling Pathway. Mar. Drugs 2024, 22, 275. https://doi.org/10.3390/md22060275
Xu C, Cao G, Zhang H, Bai M, Yi X, Qu X. Avellanin A Has an Antiproliferative Effect on TP-Induced RWPE-1 Cells via the PI3K-Akt Signalling Pathway. Marine Drugs. 2024; 22(6):275. https://doi.org/10.3390/md22060275
Chicago/Turabian StyleXu, Chang, Guangping Cao, Hong Zhang, Meng Bai, Xiangxi Yi, and Xinjian Qu. 2024. "Avellanin A Has an Antiproliferative Effect on TP-Induced RWPE-1 Cells via the PI3K-Akt Signalling Pathway" Marine Drugs 22, no. 6: 275. https://doi.org/10.3390/md22060275
APA StyleXu, C., Cao, G., Zhang, H., Bai, M., Yi, X., & Qu, X. (2024). Avellanin A Has an Antiproliferative Effect on TP-Induced RWPE-1 Cells via the PI3K-Akt Signalling Pathway. Marine Drugs, 22(6), 275. https://doi.org/10.3390/md22060275