
Phototoxicity of the Ethanolic Extract of Skeletonema marinoi for the Dermocosmetic Improvement of Acne
 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Phototoxicity of the Skeletonema sp. Ethanol Extracts in Planktonic Bacteria
2.2. Phototoxicity of the Skeletonema marinoi Ethanol Extract in Planktonic S. aureus Depends on Illumination Energy

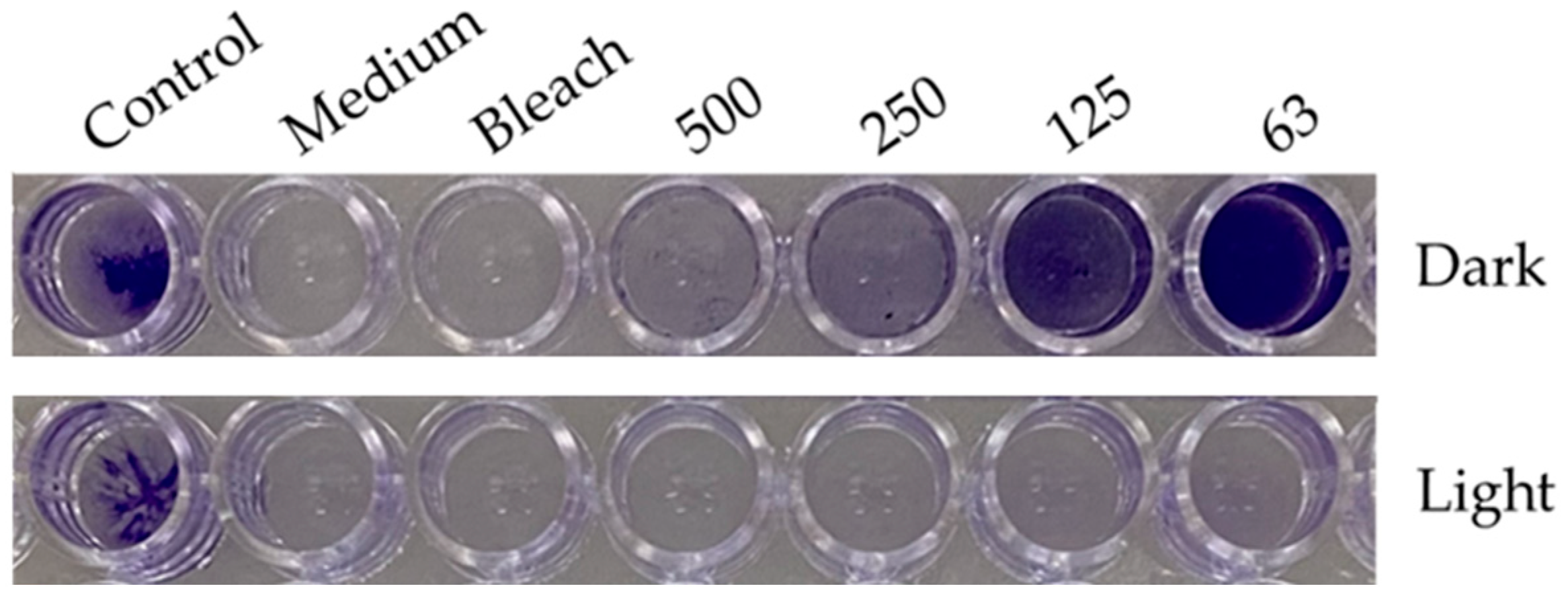
2.3. Phototoactivation of the Skeletonema marinoi Ethanol Extract Inhibits the Formation of S. aureus Biofilms and Destroys Established Biofilms
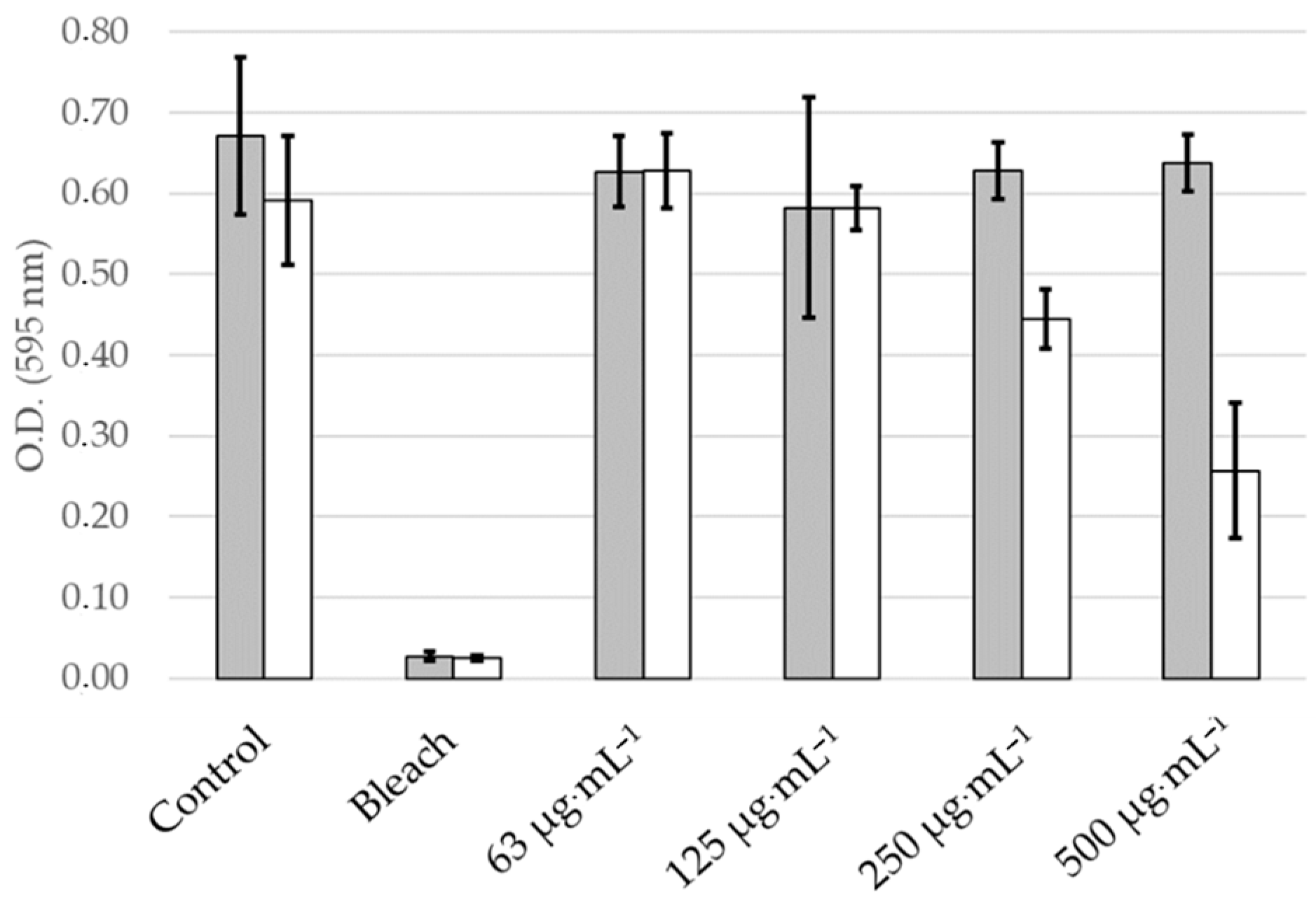
2.4. Toxicity and Phototoxicity of the Skeletonema marinoi Ethanol Extract in Primary Normal Human Epidermal Keratinocytes (NHEK)
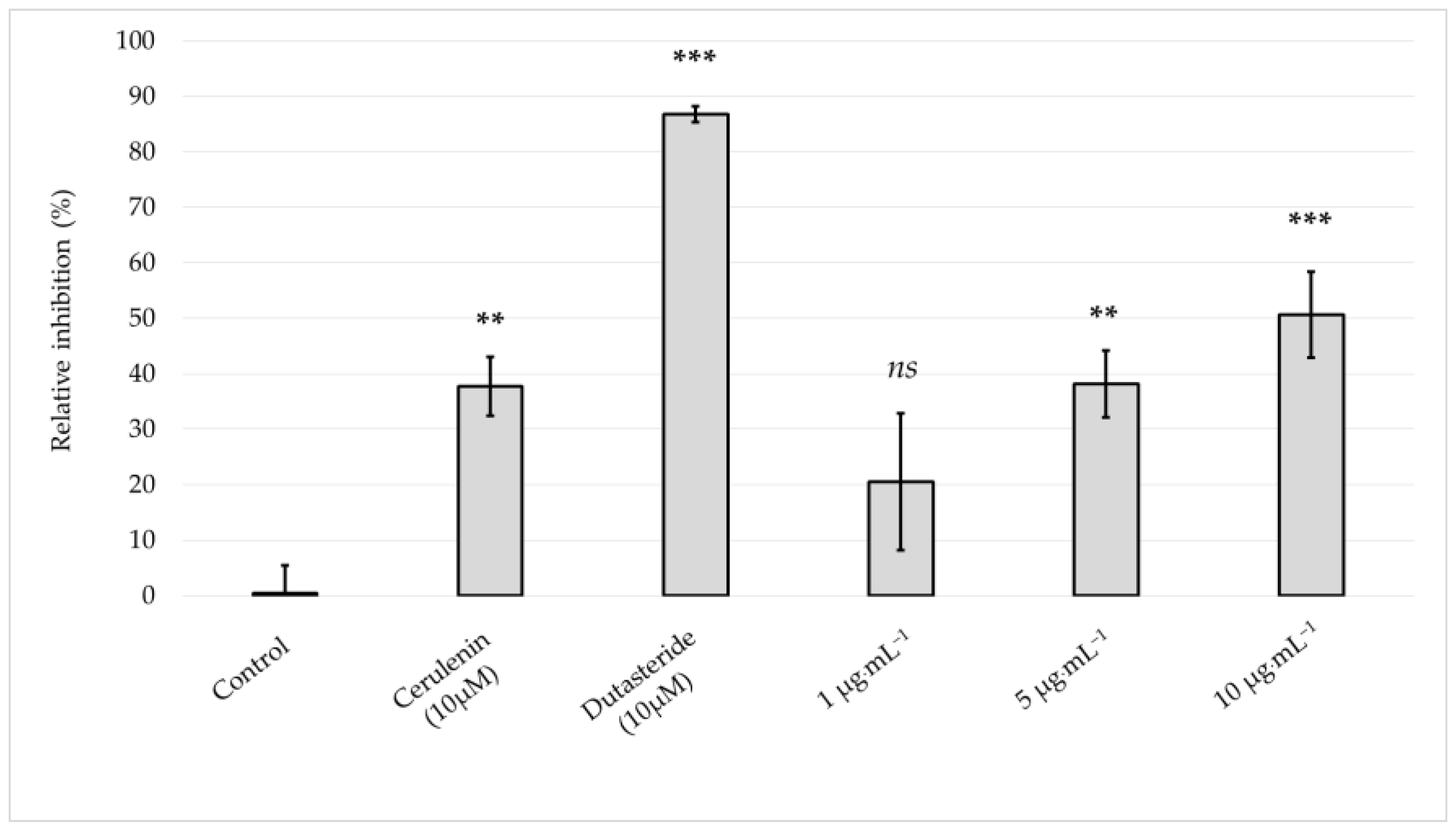
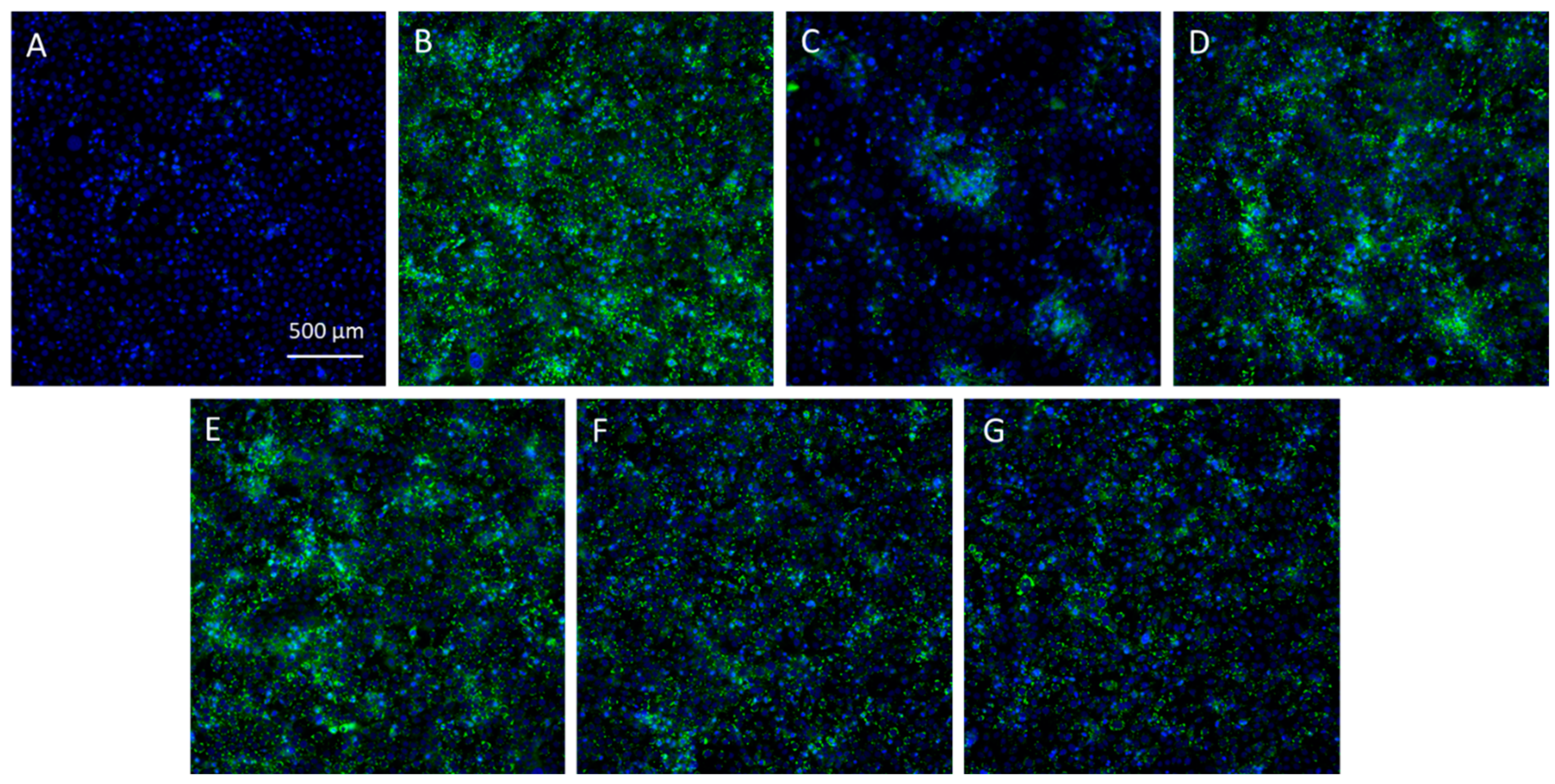
2.5. Antilipogenic Activity of the Skeletonema marinoi Ethanol Extract in the Human Sebocyte Cell Line SEBO662AR
2.6. Analysis of the Pigment Composition of the Skeletonema marinoi Ethanol Extract
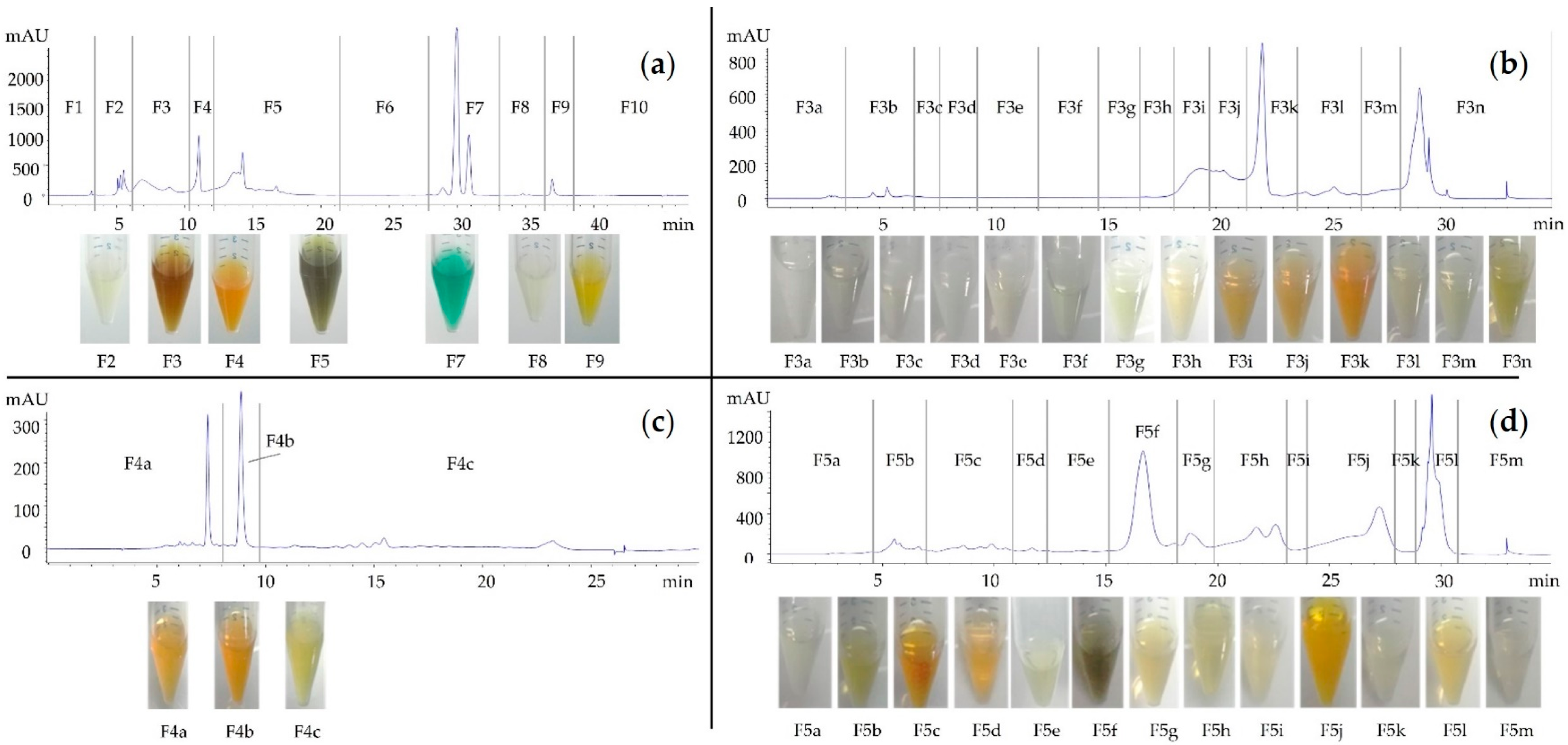
2.7. Bioguided Fractionation of the Skeletonema marinoi Ethanol Extract
2.8. Correlation of Phototoxicity and Pheophorbide a Content through Principal Component Analysis in Various Species of Skeletonema and Phenotyping of the Pigment Contents among Different Strains of Skeletonema marinoi
3. Conclusive Remarks
4. Materials and Methods
4.1. Microalgae Collection and Cultures
4.2. Molecular Identification of Microalgae
4.3. Pigment Extraction and Analysis
4.3.1. Extraction of Pigments for Phototoxicity Screenings
4.3.2. Extraction for Pigment Diversity Analysis
4.3.3. HPLC Analysis
4.3.4. Semi-Preparative HPLC for the Crude-Extract Fractionation
4.3.5. LC-HRMS Analysis of Phototoxic Pigment Extracts
4.4. Evaluation of the Cytotoxicity at Obscurity and Phototoxicity after Illumination against Planktonic and Biofilm Bacteria (MIC/MBC Assays)
4.4.1. Bacterial Strains and Growth
4.4.2. Determination of MIC and MBC in Dark Condition (Cytotoxicity) and after Light Illumination (Phototoxicity) in Planktonic Bacteria
4.4.3. Biofilm Inhibition Assay in Dark Condition and after Light Illumination
4.4.4. Biofilm Eradication Assay in Dark Condition and after Light Illumination
4.5. Cytotoxicity and Phototoxicity in Keratinocytes
4.6. Cytotoxicity and Lipogenesis Inhibition in Sebocytes
4.7. Interspecies Comparison and Correlation between Phototoxicity and Pheophorbide a and Derivatives Content
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tan, J.K.L.; Bhate, K. A Global Perspective on the Epidemiology of Acne. Br. J. Dermatol. 2015, 172 (Suppl. 1), 3–12. [Google Scholar] [CrossRef]
- Dréno, B.; Pécastaings, S.; Corvec, S.; Veraldi, S.; Khammari, A.; Roques, C. Cutibacterium Acnes (Propionibacterium Acnes) and Acne Vulgaris: A Brief Look at the Latest Updates. J. Eur. Acad. Dermatol. Venereol. 2018, 32 (Suppl. 2), 5–14. [Google Scholar] [CrossRef]
- Dréno, B.; Dagnelie, M.A.; Khammari, A.; Corvec, S. The Skin Microbiome: A New Actor in Inflammatory Acne. Am. J. Clin. Dermatol. 2020, 21, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Totté, J.E.E.; van der Feltz, W.T.; Bode, L.G.M.; van Belkum, A.; van Zuuren, E.J.; Pasmans, S.G.M.A. A Systematic Review and Meta-Analysis on Staphylococcus Aureus Carriage in Psoriasis, Acne and Rosacea. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus Aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial Biofilms: A Common Cause of Persistent Infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Li, S.; Liu, Y.; Xu, B.; Liu, S.; Zhang, J.; Zhou, D.; Liu, H. Advances in Phototherapy for Infectious Diseases. Nano Today 2024, 57, 102327. [Google Scholar] [CrossRef]
- Elkihel, A.; Christie, C.; Vernisse, C.; Ouk, T.-S.; Lucas, R.; Chaleix, V.; Sol, V. Xylan-Based Cross-Linked Hydrogel for Photodynamic Antimicrobial Chemotherapy. ACS Appl. Bio Mater. 2021, 4, 7204–7212. [Google Scholar] [CrossRef]
- Ringot, C.; Saad, N.; Brégier, F.; Bressollier, P.; Poli, E.; Chaleix, V.; Ouk, T.S.; Sol, V. Antibacterial Activity of a Photosensitive Hybrid Cellulose Fabric. Photochem. Photobiol. Sci. 2018, 17, 1780–1786. [Google Scholar] [CrossRef]
- Cieplik, F.; Deng, D.; Crielaard, W.; Buchalla, W.; Hellwig, E.; Al-Ahmad, A.; Maisch, T. Antimicrobial Photodynamic Therapy—What We Know and What We Don’t. Crit. Rev. Microbiol. 2018, 44, 571–589. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, F.; Khan, A.U.; Misba, L.; Akhtar, K.; Ali, A. Antimicrobial and Antibiofilm Photodynamic Therapy against Vancomycin Resistant Staphylococcus Aureus (VRSA) Induced Infection in Vitro and in Vivo. Eur. J. Pharm. Biopharm. 2021, 160, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Yan, E.; Kwek, G.; Qing, N.S.; Lingesh, S.; Xing, B. Antimicrobial Photodynamic Therapy for the Remote Eradication of Bacteria. ChemPlusChem 2023, 88, e202300009. [Google Scholar] [CrossRef] [PubMed]
- Mang, T.; Rogers, S.; Keinan, D.; Honma, K.; Baier, R. Antimicrobial Photodynamic Therapy (aPDT) Induction of Biofilm Matrix Architectural and Bioadhesive Modifications. Photodiagnosis Photodyn. Ther. 2016, 13, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Alves-Silva, E.G.; Arruda-Vasconcelos, R.; Louzada, L.M.; de-Jesus-Soares, A.; Ferraz, C.C.R.; Almeida, J.F.A.; Marciano, M.A.; Steiner-Oliveira, C.; Santos, J.M.M.; Gomes, B.P. Effect of Antimicrobial Photodynamic Therapy on the Reduction of Bacteria and Virulence Factors in Teeth with Primary Endodontic Infection. Photodiagnosis Photodyn. Ther. 2023, 41, 103292. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, H.; Pourhajibagher, M.; Alikhani, M.Y.; Bahador, A. The Effect of Antimicrobial Photodynamic Therapy on the Expression of Biofilm Associated Genes in Staphylococcus Aureus Strains Isolated from Wound Infections in Burn Patients. Photodiagnosis Photodyn. Ther. 2019, 25, 406–413. [Google Scholar] [CrossRef]
- Luo, O.D.; Bose, R.; Bawazir, M.A.; Thuraisingam, T.; Ghazawi, F.M. A Review of the Dermatologic Clinical Applications of Topical Photodynamic Therapy. J. Cutan. Med. Surg. 2024, 28, NP1–NP18. [Google Scholar] [CrossRef]
- Hongcharu, W.; Taylor, C.R.; Chang, Y.; Aghassi, D.; Suthamjariya, K.; Anderson, R.R. Topical ALA-Photodynamic Therapy for the Treatment of Acne Vulgaris. J. Invest. Dermatol. 2000, 115, 183–192. [Google Scholar] [CrossRef]
- Manivasagan, P.; Bharathiraja, S.; Santha Moorthy, M.; Mondal, S.; Seo, H.; Dae Lee, K.; Oh, J. Marine Natural Pigments as Potential Sources for Therapeutic Applications. Crit. Rev. Biotechnol. 2018, 38, 745–761. [Google Scholar] [CrossRef]
- Juin, C.; Bonnet, A.; Nicolau, E.; Bérard, J.-B.; Devillers, R.; Thiéry, V.; Cadoret, J.-P.; Picot, L. UPLC-MSE Profiling of Phytoplankton Metabolites: Application to the Identification of Pigments and Structural Analysis of Metabolites in Porphyridium Purpureum. Mar. Drugs 2015, 13, 2541–2558. [Google Scholar] [CrossRef] [PubMed]
- Grivard, A.; Goubet, I.; Duarte Filho, L.M.d.S.; Thiéry, V.; Chevalier, S.; de Oliveira-Junior, R.G.; El Aouad, N.; Guedes da Silva Almeida, J.R.; Sitarek, P.; Quintans-Junior, L.J.; et al. Archaea Carotenoids: Natural Pigments with Unexplored Innovative Potential. Mar. Drugs 2022, 20, 524. [Google Scholar] [CrossRef] [PubMed]
- Serive, B.; Nicolau, E.; Bérard, J.-B.; Kaas, R.; Pasquet, V.; Picot, L.; Cadoret, J.-P. Community Analysis of Pigment Patterns from 37 Microalgae Strains Reveals New Carotenoids and Porphyrins Characteristic of Distinct Strains and Taxonomic Groups. PLoS ONE 2017, 12, e0171872. [Google Scholar] [CrossRef]
- Baudelet, P.-H.; Gagez, A.-L.; Bérard, J.-B.; Juin, C.; Bridiau, N.; Kaas, R.; Thiéry, V.; Cadoret, J.-P.; Picot, L. Antiproliferative Activity of Cyanophora Paradoxa Pigments in Melanoma, Breast and Lung Cancer Cells. Mar. Drugs 2013, 11, 4390–4406. [Google Scholar] [CrossRef]
- Villanova, V.; Engelbrektsson, J.; Strömberg, N.; Ekendahl, S.; Spetea, C. Unveiling the Ecological Resilience and Industrial Potential of Skeletonema Marinoi through Mixotrophic Cultivation in Nordic Winter Condition. Physiol. Plant. 2024, 176, e14308. [Google Scholar] [CrossRef] [PubMed]
- Volpe, C.; Nymark, M.; Andersen, T.; Winge, P.; Lavaud, J.; Vadstein, O. Skeletonema Marinoi Ecotypes Show Specific Habitat-Related Responses to Fluctuating Light Supporting High Potential for Growth under Photobioreactor Light Regime. New Phytol. 2024, 243, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Smerilli, A.; Orefice, I.; Corato, F.; Gavalás Olea, A.; Ruban, A.V.; Brunet, C. Photoprotective and Antioxidant Responses to Light Spectrum and Intensity Variations in the Coastal Diatom Skeletonema Marinoi. Environ. Microbiol. 2017, 19, 611–627. [Google Scholar] [CrossRef]
- Zulfiqar, M.; Stettin, D.; Schmidt, S.; Nikitashina, V.; Pohnert, G.; Steinbeck, C.; Peters, K.; Sorokina, M. Untargeted Metabolomics to Expand the Chemical Space of the Marine Diatom Skeletonema Marinoi. Front. Microbiol. 2023, 14, 1295994. [Google Scholar] [CrossRef]
- Cotterill, J.A. Benzoyl Peroxide. Acta Derm. Venereol. Suppl. 1980, Suppl. 89, 57–63. [Google Scholar] [CrossRef]
- de Annunzio, S.R.; de Freitas, L.M.; Blanco, A.L.; da Costa, M.M.; Carmona-Vargas, C.C.; de Oliveira, K.T.; Fontana, C.R. Susceptibility of Enterococcus Faecalis and Propionibacterium Acnes to Antimicrobial Photodynamic Therapy. J. Photochem. Photobiol. B 2018, 178, 545–550. [Google Scholar] [CrossRef]
- Dharmaraja, A.T. Role of Reactive Oxygen Species (ROS) in Therapeutics and Drug Resistance in Cancer and Bacteria. J. Med. Chem. 2017, 60, 3221–3240. [Google Scholar] [CrossRef] [PubMed]
- Staerck, C.; Gastebois, A.; Vandeputte, P.; Calenda, A.; Larcher, G.; Gillmann, L.; Papon, N.; Bouchara, J.-P.; Fleury, M.J.J. Microbial Antioxidant Defense Enzymes. Microb. Pathog. 2017, 110, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjya, R.; Kiran Marella, T.; Tiwari, A.; Saxena, A.; Kumar Singh, P.; Mishra, B. Bioprospecting of Marine Diatoms Thalassiosira, Skeletonema and Chaetoceros for Lipids and Other Value-Added Products. Bioresour. Technol. 2020, 318, 124073. [Google Scholar] [CrossRef] [PubMed]
- Circu, M.L.; Aw, T.Y. Reactive Oxygen Species, Cellular Redox Systems, and Apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of Apoptosis Signalling Pathways by Reactive Oxygen Species. Biochim. Biophys. Acta-Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, I.; Danby, F.W.; Ju, Q.; Wang, X.; Xiang, L.F.; Xia, L.; Chen, W.; Nagy, I.; Picardo, M.; Suh, D.H.; et al. New Developments in Our Understanding of Acne Pathogenesis and Treatment. Exp. Dermatol. 2009, 18, 821–832. [Google Scholar] [CrossRef]
- Van Heukelem, L.; Thomas, C.S. Computer-Assisted High-Performance Liquid Chromatography Method Development with Applications to the Isolation and Analysis of Phytoplankton Pigments. J. Chromatogr. A 2001, 910, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, E.; Bishayee, A.; Fimognari, C. On a Beam of Light: Photoprotective Activities of the Marine Carotenoids Astaxanthin and Fucoxanthin in Suppression of Inflammation and Cancer. Mar. Drugs 2020, 18, 544. [Google Scholar] [CrossRef]
- Kuczynska, P.; Jemiola-Rzeminska, M.; Strzalka, K. Photosynthetic Pigments in Diatoms. Mar. Drugs 2015, 13, 5847–5881. [Google Scholar] [CrossRef]
- Pajot, A.; Hao Huynh, G.; Picot, L.; Marchal, L.; Nicolau, E. Fucoxanthin from Algae to Human, an Extraordinary Bioresource: Insights and Advances in up and Downstream Processes. Mar. Drugs 2022, 20, 222. [Google Scholar] [CrossRef]
- Wainwright, M. Photodynamic Therapy: The Development of New Photosensitisers. Anti-Cancer Agents Med. Chem. 2008, 8, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Kandagatla, S.K.; Uhl, R.T.; Graf, T.N.; Oberlies, N.H.; Raner, G.M. Pheophorbide Derivatives Isolated from Açaí Berries (Euterpea oleracea) Activate an Antioxidant Response Element In Vitro. Nat. Prod. Commun. 2019, 14, 1934578X19852443. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.K.; Shiau, F.-Y.; Sumlin, A.B.; Dougherty, T.J.; Smith, K.M. Structure/Activity Relationships among Photosensitizers Related to Pheophorbides and Bacteriopheophorbides. Bioorganic Med. Chem. Lett. 1992, 2, 491–496. [Google Scholar] [CrossRef]
- Saide, A.; Lauritano, C.; Ianora, A. Pheophorbide a: State of the Art. Mar. Drugs 2020, 18, 257. [Google Scholar] [CrossRef] [PubMed]
- Xodo, L.E.; Rapozzi, V.; Zacchigna, M.; Drioli, S.; Zorzet, S. The Chlorophyll Catabolite Pheophorbide a as a Photosensitizer for the Photodynamic Therapy. Curr. Med. Chem. 2012, 19, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.; Jeffrey, S.W. A Note on the Occurrence of Chlorophyllase in Marine Algae. J. Exp. Mar. Biol. Ecol. 1971, 7, 255–262. [Google Scholar] [CrossRef]
- Jeffrey, S.W.; Hallegraeff, G.M. Chlorophyllase Distribution in Ten Classes of Phytoplankton: A Problem for Chlorophyll Analysis. Mar. Ecol. Progress Ser. 1987, 35, 293–304. [Google Scholar] [CrossRef]
- Karpiński, T.M.; Adamczak, A. Fucoxanthin—An Antibacterial Carotenoid. Antioxidants 2019, 8, 239. [Google Scholar] [CrossRef]
- Stoermer, E.F.; Julius, M.L. 15—Centric diatoms. In Freshwater Algae of North America; Wehr, J.D., Sheath, R.G., Eds.; Aquatic Ecology; Academic Press: Burlington, MA, USA, 2003; pp. 559–594. ISBN 978-0-12-741550-5. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. marinoi | S. grethae | S. menzelii | S. subsalsum | |
|---|---|---|---|---|
| C. acnes | ||||
| Dark (control) | >1000 | >1000 | >1000 | >1000 |
| White light | 6 | 6 | 31 | 16 |
| Red light | 125 | 500 | >1000 | 1000 |
| S. aureus | ||||
| Dark (control) | 250 | 250 | >1000 | >1000 |
| White light | 63 | 31 | 250 | 125 |
| Red light | 125 | 125 | >1000 | >1000 |
| S. epidermidis | ||||
| Dark (control) | 250 | 250 | 250 | 250 |
| White light | 16 | 63 | 63 | 16 |
| Red light | 63 | 250 | 250 | 125 |
| MBC White Light (µg·mL−1) | MBC Dark (µg·mL−1) | |
|---|---|---|
| C. acnes | 6 | >1000 |
| S. aureus | 63 | >1000 |
| S. epidermidis | 63 | >1000 |
| Molecule | Formula | Ion | Theoretical Monoisotopic Mass (m/z) | Peak Area (×106) | RT (min) | Score (%) | Experimental Monoisotopic Mass (m/z) | Error (ppm) |
|---|---|---|---|---|---|---|---|---|
| Pheophorbide a | C35H37O5N4 | [M + H]+ | 593.2758 | 303 | 11.18 | 98.5 | 593.2775 | 2.79 |
| [M + K]+ | 631.2312 | 631.2343 | 4.91 | |||||
| [M + Na]+ | 615.2578 | 615.2579 | 0.16 | |||||
| Pheophorbide a isomer | C35H37O5N4 | [M + H]+ | 593.2758 | 127 | 11.43 | 97.3 | 593.2760 | 0.26 |
| [M + K]+ | 631.2312 | 631.2292 | −3.17 | |||||
| [M + Na]+ | 615.2578 | ND | ||||||
| all-trans-Fucoxanthin | C42H58O6 | [M + H]+ | 659.4306 | 184 | 10.88 | 97.6 | 659.4307 | 0.13 |
| [M + H − H2O]+ | 641.4201 | 641.4201 | 0.08 | |||||
| [M + Na]+ | 681.4126 | 681.4134 | 1.23 | |||||
| cis-Fucoxanthin | C42H58O6 | [M + H]+ | 659.4306 | 359 | 11.52 | 95.8 | 659.4324 | 2.71 |
| [M + H − H2O]+ | 641.4201 | 641.4184 | −2.57 | |||||
| [M + Na]+ | 681.4126 | 681.4147 | 3.14 | |||||
| 13-OH pheophorbide a | C35H36N4O6 | [M + H]+ | 609.2708 | 465 | 11.01 | 99.4 | 609.2709 | 0.16 |
| [M + K]+ | 647.2266 | 647.2386 | 18.54 | |||||
| [M + Na]+ | 631.2527 | 631.2530 | 0.48 |
| Sub-Fraction | S.aureus Growth Inhibition under White Light | S.aureus Growth Inhibition under Red Light | S.aureus Growth Inhibition at Dark (Control) |
|---|---|---|---|
| F3a to F3i | - | - | - |
| F3j | + | + | + |
| F3k to F3n | - | - | - |
| F4a | - | - | - |
| F4b | + | + | + |
| F4c | - | - | - |
| F5a to F5e | - | - | - |
| F5f | + | + | - |
| F5g | + | + | - |
| F5h to F5n | - | - | - |
| S. marinoi IFR-855 | S. grethae CCAP 1077/3 | S. subsalsum SAG 8.94 | S. menzelii CCMP 787 | |
|---|---|---|---|---|
| Chlorophyll c2 | 0.36 | 0.20 | 0.05 | 0.56 |
| Chlorophyll c1 | 0.17 | 0.12 | 0.03 | Undetected |
| Chlorophyll a | 2.49 | 3.22 | 6.36 | 5.05 |
| Pheophorbide a | 5.94 | 0.50 | Undetected | Undetected |
| cis-Fucoxanthin | 0.06 | Undetected | 0.05 | 0.12 |
| All Trans-Fucoxanthin | 1.98 | 1.52 | 1.73 | 4.64 |
| Diadinoxanthin | 0.07 | 0.36 | 0.73 | 0.63 |
| Diatoxanthin | 1.18 | 0.18 | 0.49 | 0.49 |
| Violaxanthin | Undetected | Undetected | 0.02 | Undetected |
| Zeaxanthin | 0.10 | 0.04 | 0.04 | Undetected |
| β-Carotene | 0.33 | 0.16 | 0.20 | 0.15 |
| S. marinoi IFR-855 | S. marinoi AC-714 | S. marinoi CCAP 1077/5 | S. marinoi SAG 19.99 | |
|---|---|---|---|---|
| Chlorophyll c2 | 0.143 ± 0.003 | 0.207 ± 0.003 | 0.336 ± 0.077 | 0.510 ± 0.115 |
| Chlorophyll c1 | 0.118 ± 0.002 | 0.200 ± 0.004 | 0.386 ± 0.049 | 0.394 ± 0.066 |
| Chlorophyll a | 0.135 ± 0.018 | 0.493 ± 0.126 | 0.302 ± 0.065 | 0.416 ± 0.042 |
| Pheophorbide a | 1.968 ± 0.081 | 1.971 ± 0.180 | 1.483 ± 0.101 | 1.222 ± 0.213 |
| Fucoxanthin | 0.085 ± 0.021 | 1.615 ± 0.068 | 0.215 ± 0.102 | 0.532 ± 0.147 |
| Diadinoxanthin | 0.376 ± 0.009 | 0.237 ± 0.005 | 0.086 ± 0.030 | 0.367 ± 0.084 |
| Diatoxanthin | 0.064 ± 0.002 | 0.167 ± 0.008 | 0.055 ± 0.017 | 0.079 ± 0.020 |
| Zeaxanthin | 0.023 ± 0.006 | 0.006 ± 0.001 | 0.025 ± 0.002 | 0.000 ± 0.000 |
| β,β-Carotene | 0.021 ± 0.002 | 0.025 ± 0.001 | 0.031 ± 0.008 | 0.071 ± 0.017 |
| Ratio Pheo a/Chl a | 14.578 | 3.998 | 4.911 | 2.938 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bérard, J.-B.; Ouk, T.-S.; Sol, V.; Lefoulon, L.; Landolt, C.; Grenier, K.; Saad, N.; Robert, E.; Sibat, M.; Schreiber, N.; et al. Phototoxicity of the Ethanolic Extract of Skeletonema marinoi for the Dermocosmetic Improvement of Acne. Mar. Drugs 2024, 22, 343. https://doi.org/10.3390/md22080343
Bérard J-B, Ouk T-S, Sol V, Lefoulon L, Landolt C, Grenier K, Saad N, Robert E, Sibat M, Schreiber N, et al. Phototoxicity of the Ethanolic Extract of Skeletonema marinoi for the Dermocosmetic Improvement of Acne. Marine Drugs. 2024; 22(8):343. https://doi.org/10.3390/md22080343
Chicago/Turabian StyleBérard, Jean-Baptiste, Tan-Sothea Ouk, Vincent Sol, Louise Lefoulon, Cornelia Landolt, Karine Grenier, Naima Saad, Elise Robert, Manoëlla Sibat, Nathalie Schreiber, and et al. 2024. "Phototoxicity of the Ethanolic Extract of Skeletonema marinoi for the Dermocosmetic Improvement of Acne" Marine Drugs 22, no. 8: 343. https://doi.org/10.3390/md22080343