

Marine Bacteriocins: An Evolutionary Gold Mine to Payoff Antibiotic Resistance


Abstract
:1. Introduction
2. Diverse Bacteriocins from Marine Bacteria
2.1. Leaderless Bacteriocins
2.1.1. Piscicocin
2.1.2. Mundticin KS
2.1.3. Divercin V41
2.1.4. Divergicin M35
2.1.5. BaCf3
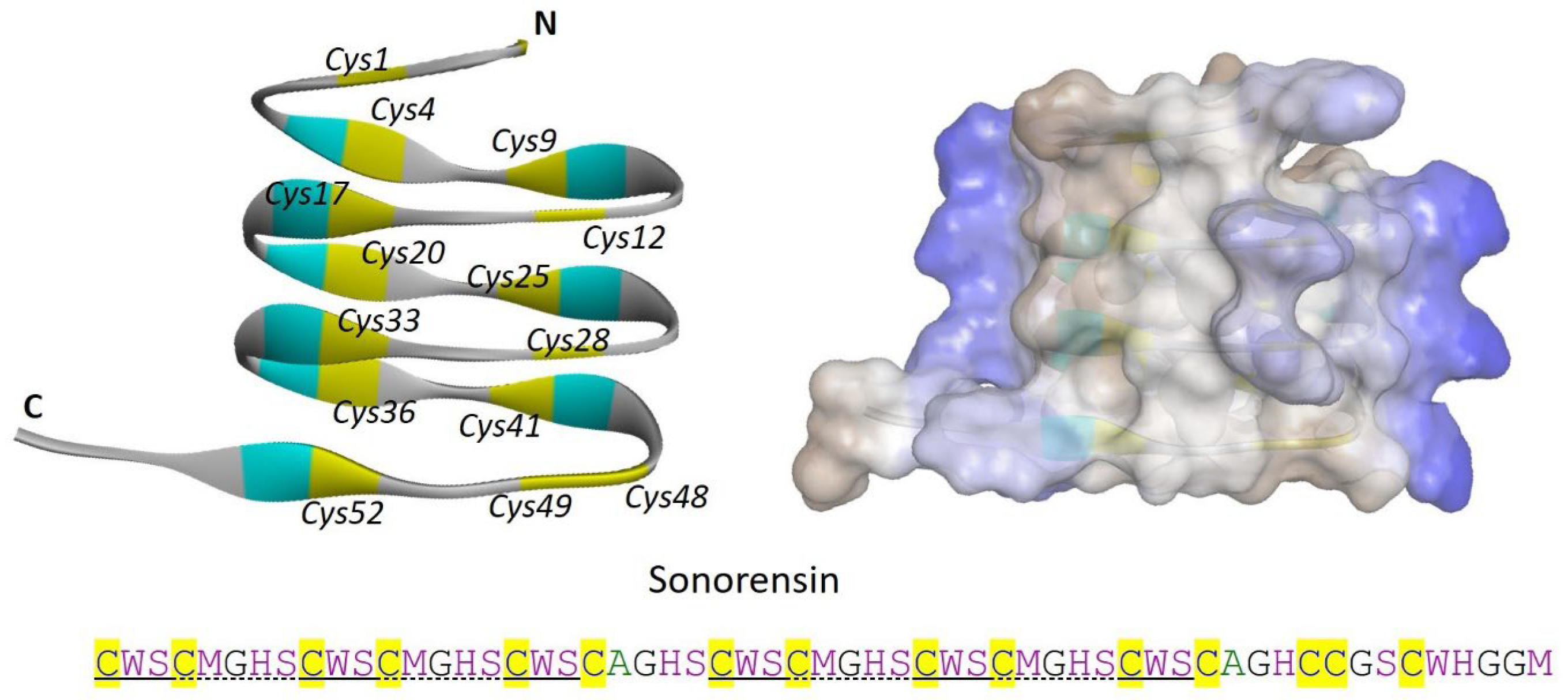
2.1.6. Sonorensin
2.1.7. CAMT6
2.2. Lanthipeptides
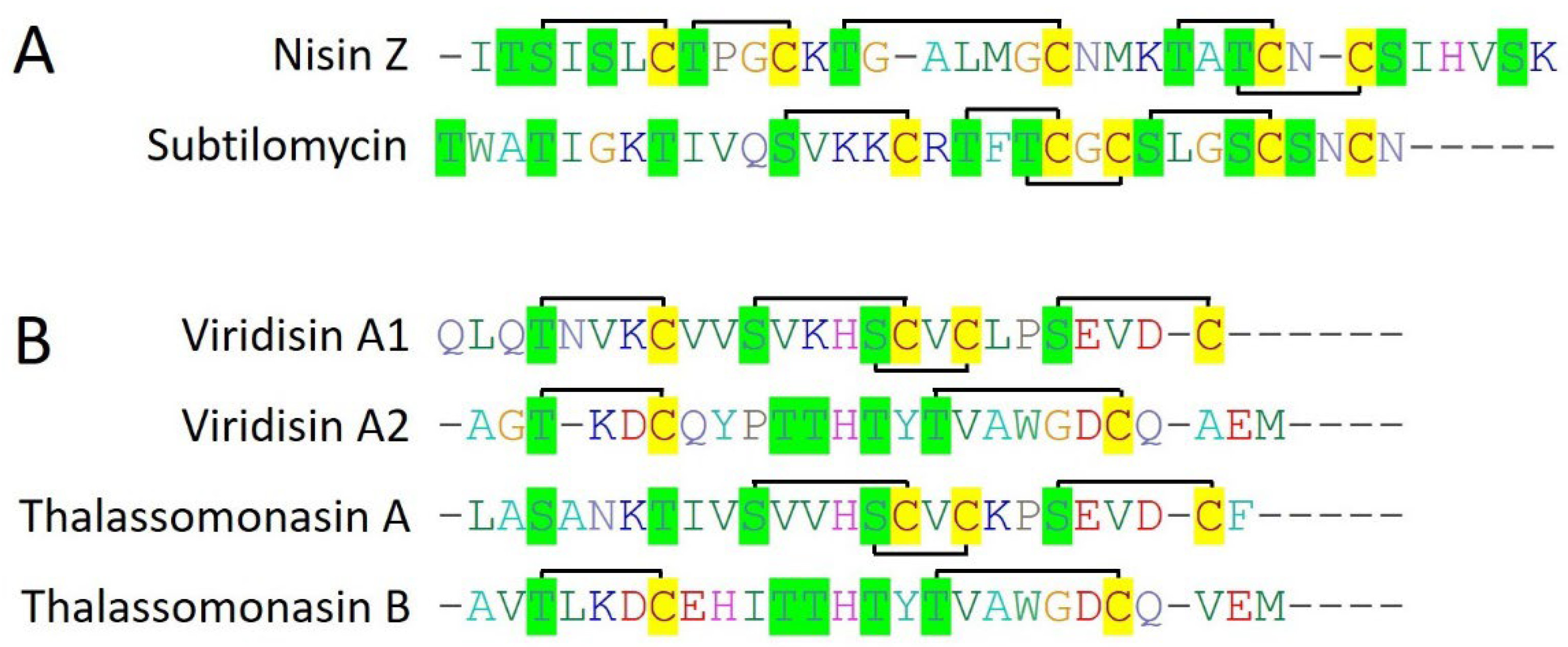
2.2.1. Nisin Z
2.2.2. Subtilomycin
2.2.3. Viridisin
2.2.4. Thalassomonasin
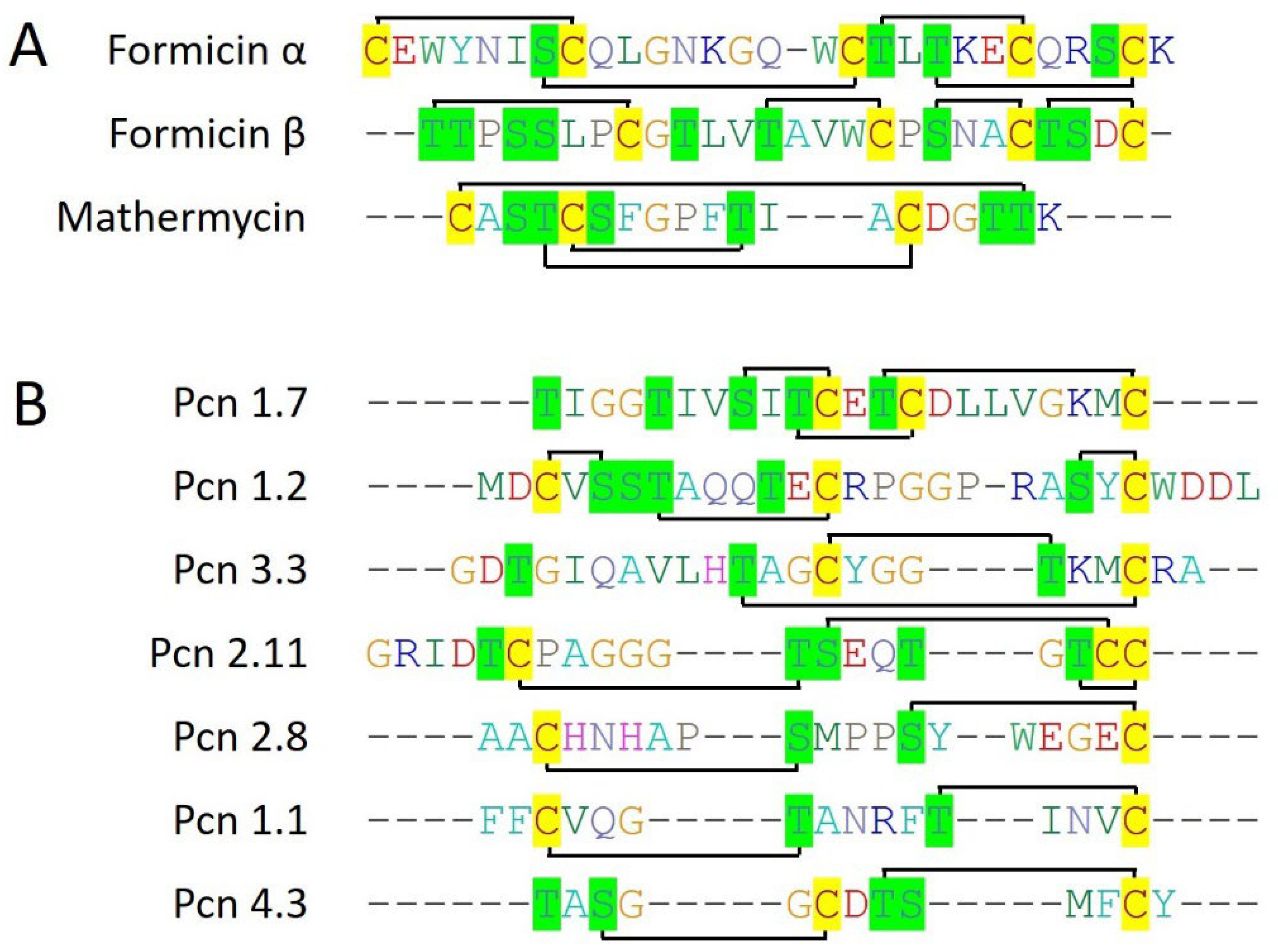
2.2.5. Formicin
2.2.6. Mathermycin
2.2.7. Prochlorosins
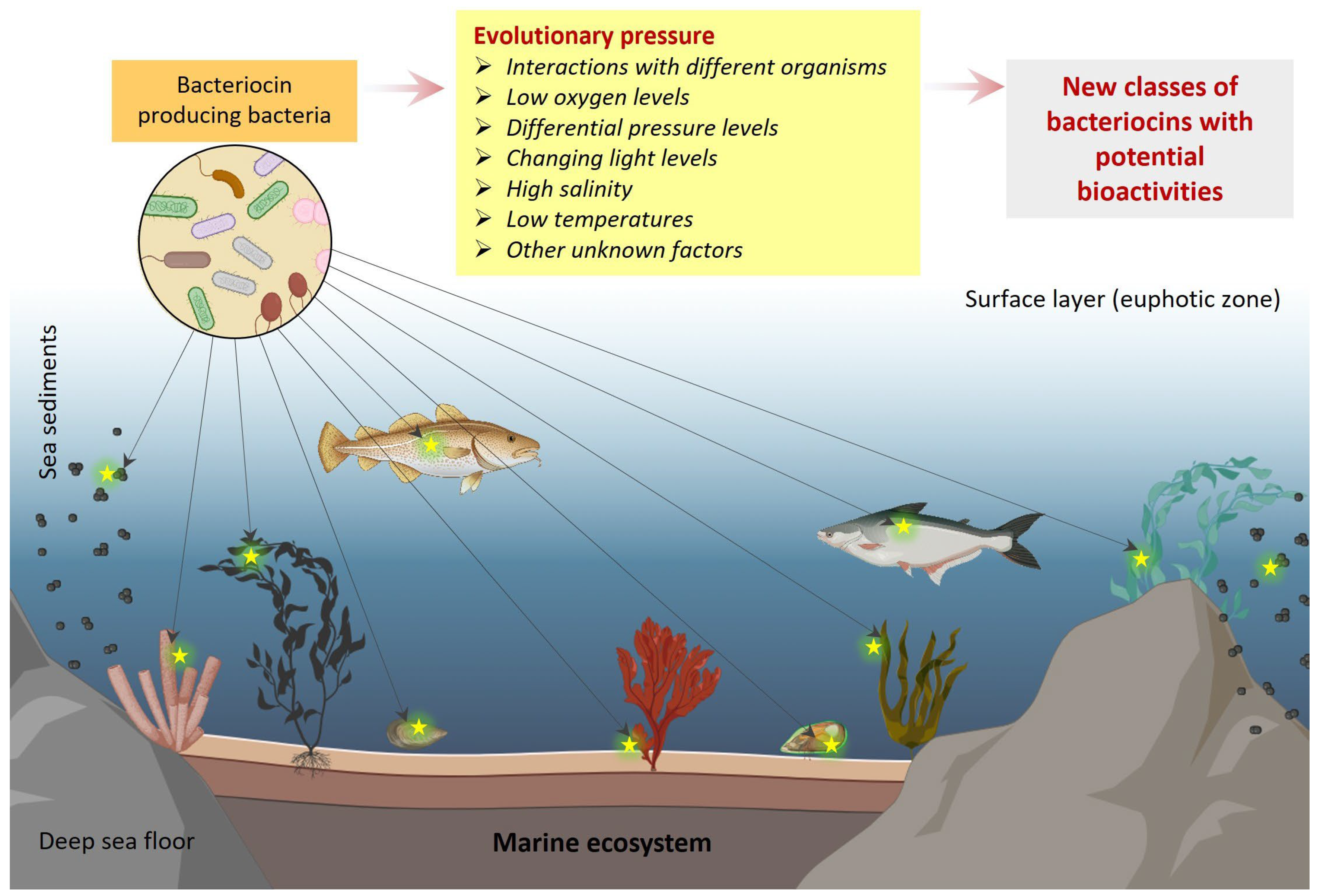
3. Evolutionary Diversity of Marine Bacteriocins
4. Challenges and Future Directions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walsh, T.R.; Gales, A.C.; Laxminarayan, R.; Dodd, P.C. Antimicrobial Resistance: Addressing a Global Threat to Humanity. PLoS Med. 2023, 20, e1004264. [Google Scholar] [CrossRef]
- Traxler, M.F.; Kolter, R. Natural products in soil microbe interactions and evolution. Nat. Prod. Rep. 2015, 32, 956–970. [Google Scholar] [CrossRef] [PubMed]
- Baindara, P.; Chaudhry, V.; Mittal, G.; Liao, L.M.; Matos, C.O.; Khatri, N.; Franco, O.L.; Patil, P.B.; Korpole, S. Characterization of the antimicrobial peptide penisin, a class Ia novel lantibiotic from Paenibacillus sp. Strain A3. Antimicrob. Agents Chemother. 2016, 60, 580–591. [Google Scholar] [CrossRef]
- Donia, M.S.; Cimermancic, P.; Schulze, C.J.; Wieland Brown, L.C.; Martin, J.; Mitreva, M.; Clardy, J.; Linington, R.G.; Fischbach, M.A. A systematic analysis of biosynthetic gene clusters in the human microbiome reveals a common family of antibiotics. Cell 2014, 158, 1402–1414. [Google Scholar] [CrossRef]
- Ziemert, N.; Lechner, A.; Wietz, M.; Millań-Aguiñaga, N.; Chavarria, K.L.; Jensen, P.R. Diversity and evolution of secondary metabolism in the marine actinomycete genus Salinispora. Proc. Natl. Acad. Sci. USA 2014, 111, E1130–E1139. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Ghosh, C.; Sarkar, P.; Issa, R.; Haldar, J. Alternatives to Conventional Antibiotics in the Era of Antimicrobial Resistance. Trends Microbiol. 2019, 27, 323–338. [Google Scholar] [CrossRef]
- Zimina, M.; Babich, O.; Prosekov, A.; Sukhikh, S.; Ivanova, S.; Shevchenko, M.; Noskova, S. Overview of global trends in classification, methods of preparation and application of bacteriocins. Antibiotics 2020, 9, 553. [Google Scholar] [CrossRef]
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160. [Google Scholar] [CrossRef] [PubMed]
- Willey, J.M.; van der Donk, W. A Lantibiotics: Peptides of diverse structure and function. Annu. Rev. Microbiol. 2007, 61, 477–501. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, T.; Doi, H.; Uramoto, G.I.; Wörmer, L.; Adhikari, R.R.; Xiao, N.; Morono, Y.; D’Hondt, S.; Hinrichs, K.U.; Inagaki, F. Global diversity of microbial communities in marine sediment. Proc. Natl. Acad. Sci. USA 2020, 117, 27587–27597. [Google Scholar] [CrossRef]
- Pilet, M.F.; Xavier, D.; Rachel, B.; Georges, N.; Michel, D.; Piard, J.C. Evidence for two bacteriocins produced by Carnobacterium piscicola and Carnobacterium divergens isolated from fish and active against Listeria monocytogenes. J. Food Prot. 1995, 58, 256–262. [Google Scholar] [CrossRef]
- Bhugaloo-Vial, P.; Dousset, X.; Metivier, A.; Sorokine, O.; Anglade, P.; Boyaval, P.; Marion, D. Purification and amino acid sequences of piscicocins V1a and V1b, two class IIa bacteriocins secreted by Carnobacterium piscicola VI that display significantly different levels of specific inhibitory activity. Appl. Environ. Microbiol. 1996, 62, 4410–4416. [Google Scholar] [CrossRef]
- Yamazaki, K.; Suzuki, M.; Kawai, Y.; Inoue, N.; Montville, T.J. Purification and characterization of a novel class IIa bacteriocin, piscicocin CS526, from surimi-associated Carnobacterium piscicola CS526. Appl. Environ. Microbiol. 2005, 71, 554–557. [Google Scholar] [CrossRef]
- Kawamoto, S.; Shima, J.; Sato, R.; Eguchi, T.; Ohmomo, S.; Shibato, J.; Horikoshi, N.; Takeshita, K.; Sameshima, T. Biochemical and genetic characterization of mundticin KS, an antilisterial peptide produced by Enterococcus mundtii NFRI 7393. Appl. Environ. Microbiol. 2002, 68, 3830–3840. [Google Scholar] [CrossRef]
- Schelegueda, L.I.; Vallejo, M.; Gliemmo, M.F.; Marguet, E.R.; Campos, C.A. Synergistic antimicrobial action and potential application for fish preservation of a bacteriocin produced by Enterococcus mundtii isolated from Odontesthes platensis. Lebensm. Wiss. Technol. 2015, 64, 794–801. [Google Scholar] [CrossRef]
- Métivier, A.; Pilet, M.F.; Dousset, X.; Sorokine, O.; Anglade, P.; Zagorec, M.; Piard, J.C.; Marion, D.; Cenatiempo, Y.; Fremaux, C. Divercin V41, a new bacteriocin with two disulphide bonds produced by Carnobacterium divergens V41: Primary structure and genomic organization. Microbiology 1998, 144, 2837–2844. [Google Scholar] [CrossRef] [PubMed]
- Tahiri, I.; Desbiens, M.; Benech, R.; Kheadr, E.; Lacroix, C.; Thibault, S.; Ouellet, D.; Fliss, I. Purification, characterization and amino acid sequencing of divergicin M35: A novel class IIa bacteriocin produced by Carnobacterium divergens M35. Int. J. Food Microbiol. 2004, 97, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Bindiya, E.S.; Tina, K.J.; Sasidharan, R.S.; Bhat, S.G. BaCf3: Highly thermostable bacteriocin from Bacillus amyloliquefaciens BTSS3 antagonistic on food-borne pathogens. 3 Biotech 2019, 9, 136. [Google Scholar] [CrossRef]
- Chopra, L.; Singh, G.; Choudhary, V.; Sahoo, D.K. Sonorensin: An antimicrobial peptide, belonging to the heterocycloanthracin subfamily of bacteriocins, from a new marine isolate, Bacillus sonorensis MT93. Appl. Environ. Microbiol. 2014, 80, 2981–2990. [Google Scholar] [CrossRef]
- Li, Q.; Chen, Q.; Wu, Y.; Chen, Z.; Liu, Y.; Fang, Z.; Deng, Q. Purification, characterization and structural identification of a novel bacteriocin produced by marine original Enterococcus durans YQ-6, and its inhibition of Listeria monocytogenes. Lebensm. Wiss. Technol. 2023, 173, 114329. [Google Scholar] [CrossRef]
- Sequeiros, C.; Garcés, M.E.; Vallejo, M.; Marguet, E.R.; Olivera, N.L. Potential aquaculture probiont Lactococcus lactis TW34 produces nisin Z and inhibits the fish pathogen Lactococcus garvieae. Arch. Microbiol. 2015, 197, 449–458. [Google Scholar] [CrossRef]
- Heo, W.S.; Kim, E.Y.; Kim, Y.R.; Hossain, M.T.; Kong, I.S. Salt effect of nisin Z isolated from a marine fish on the growth inhibition of Streptococcus iniae, a pathogen of streptococcosis. Biotechnol. Lett. 2012, 34, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Phelan, R.W.; Barret, M.; Cotter, P.D.; Connor, P.M.O.; Chen, R.; Morrissey, J.P.; Dobson, A.D.W.; Gara, F.O.; Barbosa, T.M. Subtilomycin: A New Lantibiotic from Bacillus subtilis Strain MMA7 Isolated from the Marine Sponge Haliclona simulans. Mar. Drugs 2013, 11, 1878–1898. [Google Scholar] [CrossRef]
- Vermeulen, R.; Van Staden, A.D.P.; Van Zyl, L.J.; Dicks, L.M.T.; Trindade, M. Unusual Class I Lanthipeptides from the Marine Bacteria Thalassomonas viridans. ACS Synth. Biol. 2022, 11, 3608–3616. [Google Scholar] [CrossRef] [PubMed]
- Thetsana, C.; Ijichi, S.; Kaweewan, I.; Nakagawa, H.; Kodani, S. Heterologous expression of a cryptic gene cluster from a marine proteobacterium Thalassomonas actiniarum affords new lanthipeptides thalassomonasins A and B. J. Appl. Microbiol. 2022, 132, 3629–3639. [Google Scholar] [CrossRef] [PubMed]
- Collins, F.W.J.; O’Connor, P.M.; O’Sullivan, O.; Rea, M.C.; Hill, C.; Ross, R.P. Formicin—A novel broad-spectrum twocomponent lantibiotic produced by Bacillus paralicheniformis APC 1576. Microbiology 2016, 162, 1662–1671. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.; Chen, Q.; Chen, S.; Xu, B.; Ju, J.; Wang, H. Mathermycin, a lantibiotic from the marine actinomycete Marinactinospora thermotolerans SCSIO 00652. Appl. Environ. Microbiol. 2017, 83, e00926-17. [Google Scholar] [CrossRef] [PubMed]
- Cubillos-Ruiz, A.; Berta-Thompson, J.W.; Becker, J.W.; Van Der Donk, W.A.; Chisholm, S.W. Evolutionary radiation of lanthipeptides in marine cyanobacteria. Proc. Natl. Acad. Sci. USA 2017, 114, E5424–E5433. [Google Scholar] [CrossRef]
- Dinata, R.; Baindara, P. Laterosporulin25: A probiotically produced, novel defensin-like bacteriocin and its immunogenic properties. Int. Immunopharmacol. 2023, 121, 110500. [Google Scholar] [CrossRef]
- Baindara, P.; Gautam, A.; Raghava, G.P.S.; Korpole, S. Anticancer properties of a defensin like class IId bacteriocin Laterosporulin10. Sci. Rep. 2017, 7, srep46541. [Google Scholar] [CrossRef] [PubMed]
- Mulders, J.W.M.; Boerrigter, I.J.; Rollema, H.S.; Siezen, R.J.; de Vos, W.M. Identification and characterization of the lantibiotic nisin Z, a natural nisin variant. Eur. J. Biochem. 1991, 201, 581–584. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Solanki, V.; Sharma, S.; Thakur, K.G.; Krishnan, B.; Korpole, S. The intramolecular disulfide-stapled structure of laterosporulin, a class IId bacteriocin, conceals a human defensin-like structural module. FEBS J. 2015, 282, 203–214. [Google Scholar] [CrossRef]
- Deming, J.W.; Baross, J.A. Survival, Dormancy, and Nonculturable Cells in Extreme Deep-Sea Environments. In Nonculturable Microorganisms in the Environment; Springer: Berlin/Heidelberg, Germany, 2000; pp. 147–197. [Google Scholar]
- Costello, M.J.; Chaudhary, C. Marine Biodiversity, Biogeography, Deep-Sea Gradients, and Conservation. Curr. Biol. 2017, 27, R511–R527. [Google Scholar] [CrossRef]
- Zeng, X.; Alain, K.; Shao, Z. Microorganisms from deep-sea hydrothermal vents. Mar. Life Sci. Technol. 2021, 3, 204–230. [Google Scholar] [CrossRef]
- Yadav, S.; Koenen, M.; Bale, N.J.; Reitsma, W.; Engelmann, J.C.; Stefanova, K.; Damsté, J.S.S.; Villanueva, L. Organic matter degradation in the deep, sulfidic waters of the Black Sea: Insights into the ecophysiology of novel anaerobic bacteria. Microbiome 2024, 12, 98. [Google Scholar] [CrossRef]
- Srinivasan, R.; Kannappan, A.; Shi, C.; Lin, X. Marine bacterial secondary metabolites: A treasure house for structurally unique and effective antimicrobial compounds. Mar. Drugs 2021, 19, 530. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.-E.; Kellermann, M.Y.; Schupp, P.J. Secondary Metabolites of Marine Microbes: From Natural Products Chemistry to Chemical Ecology. In YOUMARES 9—The Oceans: Our Research, Our Future; Springer: Berlin/Heidelberg, Germany, 2020; pp. 159–180. [Google Scholar]
- Baindara, P.; Mandal, S.M. Gut-Antimicrobial Peptides: Synergistic Co-Evolution with Antibiotics to Combat Multi-Antibiotic Resistance. Antibiotics 2023, 12, 1732. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Li, B.; Claesen, J.; Shi, Y.; Bibb, M.J.; Donk, W.A. Van Der Discovery of Unique Lanthionine Synthetases Reveals New Mechanistic and Evolutionary Insights. PLoS Biol. 2010, 8, 4–13. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, F.X.; Zeng, Z.; Xu, M.; Sun, F.; Yang, L.; Bi, X.; Lin, Y.; Gao, Y.J.; Hao, H.X.; et al. Advances in Metagenomics and Its Application in Environmental Microorganisms. Front. Microbiol. 2021, 12, 766364. [Google Scholar] [CrossRef]
- Van Heel, A.J.; De Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Blin, K.; Cimermancic, P.; De Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. AntiSMASH: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, 339–346. [Google Scholar] [CrossRef]
- Tietz, J.I.; Schwalen, C.J.; Patel, P.S.; Maxson, T.; Blair, P.M.; Tai, H.C.; Zakai, U.I.; Mitchell, D.A. A new genome-mining tool redefines the lasso peptide biosynthetic landscape. Nat. Chem. Biol. 2017, 13, 470–478. [Google Scholar] [CrossRef]
- Merwin, N.J.; Mousa, W.K.; Dejong, C.A.; Skinnider, M.A.; Cannon, M.J.; Li, H.; Dial, K.; Gunabalasingam, M.; Johnston, C.; Magarvey, N.A. DeepRiPP integrates multiomics data to automate discovery of novel ribosomally synthesized natural products. Proc. Natl. Acad. Sci. USA 2020, 117, 371–380. [Google Scholar] [CrossRef]
- Zhong, Z.; He, B.; Li, J.; Li, Y.X. Challenges and advances in genome mining of ribosomally synthesized and post-translationally modified peptides (RiPPs). Synth. Syst. Biotechnol. 2020, 5, 155–172. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.H.; Truman, A.W. Genome mining strategies for ribosomally synthesised and post-translationally modified peptides. Comput. Struct. Biotechnol. J. 2020, 18, 1838–1851. [Google Scholar] [CrossRef]
- Seyedsayamdost, M.R. High-Throughput platform for the discovery of elicitors of silent bacterial gene clusters. Proc. Natl. Acad. Sci. USA 2014, 111, 7266–7271. [Google Scholar] [CrossRef] [PubMed]
- Baindara, P.; Nayudu, N.; Korpole, S. Whole genome mining reveals a diverse repertoire of lanthionine synthetases and lanthipeptides among the genus Paenibacillus. J. Appl. Microbiol. 2020, 128, 473–490. [Google Scholar] [CrossRef]
- Jain, P.M.; Nellikka, A.; Kammara, R. Understanding bacteriocin heterologous expression: A review. Int. J. Biol. Macromol. 2024, 277, 133916. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AMP | Class | Producer Bacteria | Source | Activity Spectrum | References |
|---|---|---|---|---|---|
| Leaderless | |||||
| Piscicocins V1a Piscicocin CS526 | Class IIa | C. piscicola V1 C. piscicola CS526 | Fresh fish, smoked and marinated fish, fish intestinal tract | L. monocytogenes L. sake L. curvatus L. plantarum L. mesenteroıdes P. acidilactici E. faecalis | [12,14] |
| Mundticin KS | Class IIa | E. mundtii NFRI 7393 | Silverside (Odontesthes platensis) | E. faecium L. plantarum L. lactis L. curvatus L. monocytogenes P. pentosaceus S. thermophiles S. pyogenes P. aeruginosa S. putrefaciens | [15,16] |
| Diversin V41 | Class IIa | C. divergens V41 | Fish viscera | Gram-positive bacteria | [12,17] |
| Divergicin M35 | Class IIa | C. divergens M35 | Frozen smoked mussels | L. monocytogenes | [18] |
| BaCf3 | Not assigned | B. amyloliquefaciens BTSS3 | Deep-sea shark (Centroscyllium fabricii) | Bacillus sp. S. warnie M. luteus G. stearothermophilus S. typhimurium C. perfringens E. faecalis | [19] |
| Sonorensin | Not assigned | B. sonorensis MT93 | Marine | L. monocytogenes V. vulnificus B. subtilis S. aureus P. aeruginosa E. coli | [20] |
| CAMT6 | Not assigned | E. durans YQ-6 | Marine fish (Larimichthys polyactis) | S. aureus B. subtilis B. equi B. cereus S. haemolyticus P. acnes S.paratyphi V. parahaemolyticus P. foulis E. aerogenes L. monocytogenes | [21] |
| Lanthipeptides | |||||
| Nisin Z | Type I lantibiotic | L. lactic NIZO 22186 | Marine fish (Olive flounder) | S. iniae L. garvieae | [22,23] |
| Subtilomycin | Type I lantibiotic | B. subtilis MMA7 | Marine sponge (Haliclona simulans) | Bacillus sp. Clostridium L. monocytogenes S. aureus P. aeruginosa VISA MRSA Candida sp. | [24] |
| Viridisin | Type I lantibiotic | T. viridans XOM25 | Marine | Not determined | [25] |
| Thalassomonasin A and B | Type I lantibiotic | T. actiniarum NBRC 104231 | Marine | B. subtilis S. aureus M. luteus E. coli P. aeruginosa | [26] |
| Formicin | Type II lantibiotic | B. paralicheniformis APC 1576 | Atlantic mackerel (Scomber scombrus) | L. monocytogenes S. aureus S. mutans C. difficile Clostridia Enterococcus sp. | [27] |
| Mathermycin | Type II lantibiotic | Marinactinospora thermotolerans SCSIO 00652 | Marine sediments | B. subtilis | [28] |
| Prochlorosins | Type II lantibiotic | Prochlorococcus Synechococcus | Marine | Not determined | [29] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baindara, P.; Dinata, R.; Mandal, S.M. Marine Bacteriocins: An Evolutionary Gold Mine to Payoff Antibiotic Resistance. Mar. Drugs 2024, 22, 388. https://doi.org/10.3390/md22090388
Baindara P, Dinata R, Mandal SM. Marine Bacteriocins: An Evolutionary Gold Mine to Payoff Antibiotic Resistance. Marine Drugs. 2024; 22(9):388. https://doi.org/10.3390/md22090388
Chicago/Turabian StyleBaindara, Piyush, Roy Dinata, and Santi M. Mandal. 2024. "Marine Bacteriocins: An Evolutionary Gold Mine to Payoff Antibiotic Resistance" Marine Drugs 22, no. 9: 388. https://doi.org/10.3390/md22090388