Abstract
A new compound maleimide-5-oxime was isolated, together with 3,4-dihydroxybenzoic acid, tetillapyrone, from the ethyl acetate extract of the marine sponge Haliclona baeri while tetillapyrone, nortetillapyrone, p-hydroxybenzaldehyde and phenylacetic acid were isolated from the ethyl acetate extract of Haliclona cymaeformis, collected from the Gulf of Thailand. The structures of tetillapyrone and nortetillapyrone were re-examined using HMBC correlations. Maleimide-5-oxime, tetillapyrone and nortetillapyrone were found to be inactive against three human tumor cell lines (the estrogen-dependent ER(+) MCF-7, the estrogen-independent ER(−) MDA-MB-231 and NCI-H460. Maleimide-5-oxime, p-hydroxybenzaldehyde, phenylacetic acid, tetillapyrone and nortetillapyrone were evaluated for their growth inhibitory effect against seven yeasts and eight filamentous fungi. Only nortetillapyrone showed antifungal activity, with a preponderance on the dermatophytic filamentous fungi.
1. Introduction
The frequency of invasive fungal infection has risen substantially with the increasing numbers of immunocompromised patients, such as those infected with HIV, receiving cancer chemotherapy, immunosuppressive therapy, or treatment with broad-spectrum antibiotics [1]. The most frequently encountered infections are caused by the yeast Candida albicans and by species of filamentous fungus Aspergillus. Other fungal pathogens that have emerged in recent years include yeast species such as Candida glabata, C. krusei, C. tropicalis, Cryptococcus and Trichosporon and filamentous fungi such as Fusarium, Rhizopus and Rhizomucor [2]. On the other hand, dermatophytes, which invade and multiply in keratinized tissues [3] and cause infection, have a worldwide distribution with geographical differences in the incidence and prevalence of different dermatophyte species [4] and the most common agents belong to three genera: Epidermophyton, Trichophyton and Microsporum. Majority of clinically used antifungals suffer from various drawbacks in terms of toxicity, efficacy and cost, and their frequent use has led to the emergence of resistant strains. Hence, there is a great demand for novel antifungal agents belonging to the wide range of structural classes, selectively acting on novel targets with fewer side effects [5].
There is mounting evidence that fungi display highly specific adaptations in the marine environment that include the production of unique secondary metabolites antibiotics [1]. The fact that marine organisms contain secondary metabolites different from their terrestrial counterparts in structure and biological activity has led to the hypothesis that marine organisms may contain efficient antifungal compounds, with different modes of action and selective antifungal activity compared with human cells [6].
Sponges in the genus Haliclona are well-known for producing a variety of secondary metabolites, most commonly bioactive alkaloids [7] and many of which are cytotoxic [8]. Antifungal compounds isolated from Haliclona include haliclonadiamine, an alkaloid isolated from Haliclona sp. from Palau [9] and alkyl amino alcohols, halaminols A, B and C from the tropical marine sponge Haliclona sp., collected on the Great Barrier Reef [10].
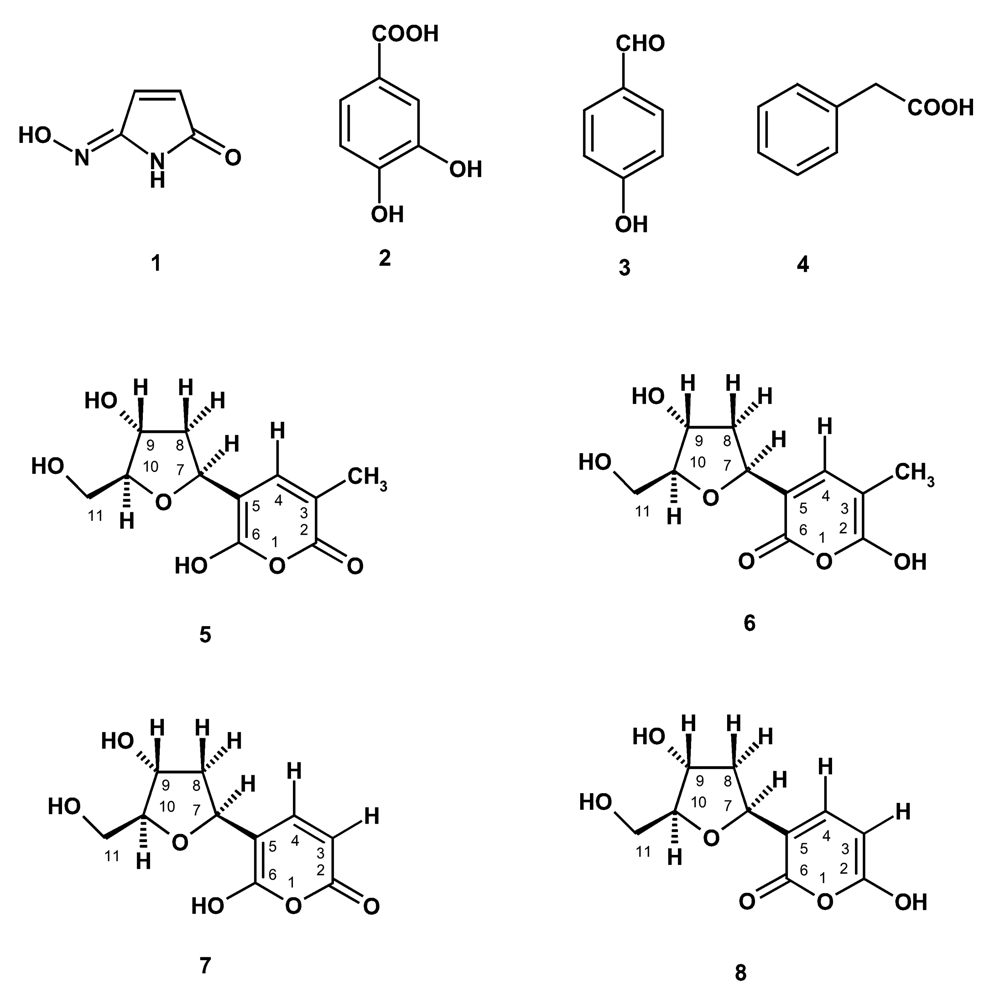
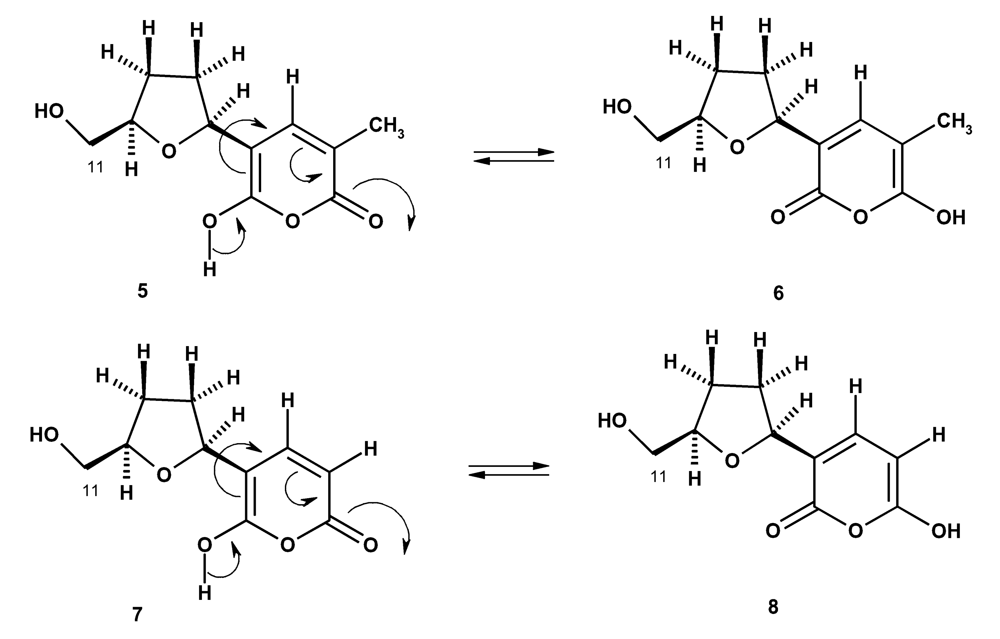
In the course of our investigation on bioactive compounds from the marine sponges from the Gulf of Thailand, we have investigated the chemical constituents of the two marine sponges Haliclona baeri and Haliclona cymaeformis, collected from the coastal area of Chonburi Province and from Koh Chang, Trad Province, Eastern Thailand, respectively. Isolated from Haliclona baeri were a new maleimide-5-oxime (1), 3,4-dihydroxybenzoic acid (2), tetillapyrone (5 and 6) while Haliclona cymaeformis furnished, besides tetillapyrone (5 and 6), p-hydroxybenzaldehyde (3), phenylacetic acid (4) and nortetillapyrone (7 and 8) (Figure 1). We have also re-examined the structures of tetillapyrone and nortetillapyrone, previously reported by our group from Tetilla japonica [11], as well as investigated the antifungal activity of the isolated compounds.
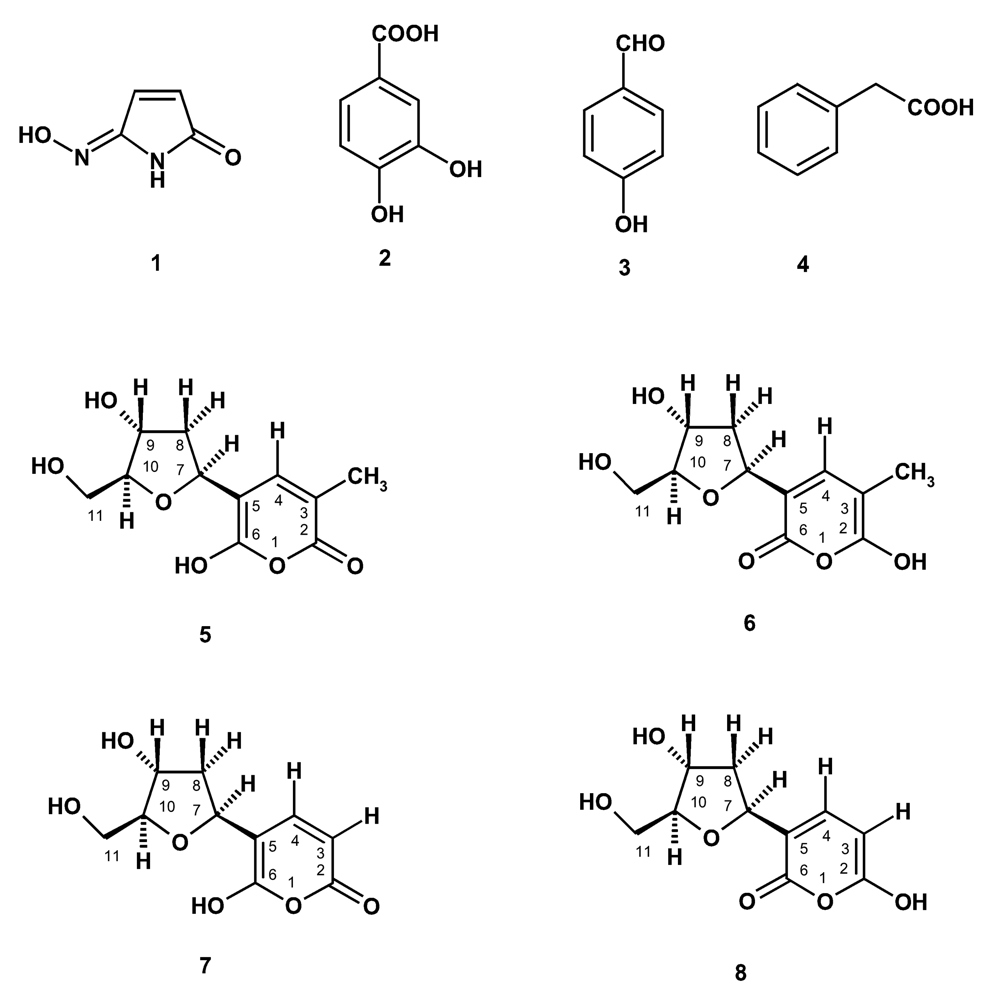
Figure 1.
Compounds isolated from Haliclona baeri and H. cymaeformis
2. Results and Discussion
We have previously reported isolation of tetillapyrone (5) and nortetillapyrone (8) from the marine sponge Tetilla japonica, collected from the Gulf of Thailand [11]. Though the structures of both compounds were established by 1H, 13CNMR, COSY, NOESY, HETCOR, HMBC and HRMS as well as by X-ray analysis in the case of tetillapyrone (5), there were still some uncertainties with respect to the pyrone moiety. The correlations observed in the HMBC spectrum for both compounds raised some doubts about the correct position of the hydroxyl and carbonyl groups of the pyrone ring. In addition, the data obtained from the X-ray analysis did not provide any evidence if a hydrogen atom was attached to the oxygen atom on C-2 or on C-6. Besides, the positions of the hydroxyl and carbonyl groups of the pyrone ring in the structure of nortetillapyrone (8) were opposite to those observed in the pryrone ring of tetillapyrone (5). All of these evidences, combined with the new isolation of more quantities of tetillapyrone and nortetillapyrone from the marine sponges H. cymaeformis and H. baeri, have led us to re-examine their structures by careful analysis of the HMBC spectra of both compounds.
The structure of tetillapyrone (5), previously proposed by us [11], was based on the fact that one carbon signal was missing in the 13C NMR spectrum and thus the carbon signal at δ109.45 was attributed to C-3 and C-5 of the pyrone ring. As the protons of the methyl group (δ1.76s) on C-3 of the pyrone ring and the hydroxyl proton at δ11.28s, showed crosspeaks with the carbon at δ109.45, we have concluded that the hydroxyl group was on C-6. In fact, it is improbable that C-3 and C-5 could have the same chemical shift value as both carbons have completely different environment. Carefully examining the HMBC at 500 MHz spectrum of tetillapyrone (Table 1), we have found that the methyl protons (δ1.78s) gave strong correlations to, besides C-4 (δ136.21), the carbons at δ109.43 (C-3), 150 54 (C-6) and 163.85 (C-2). This was very strange since the methyl protons were five bonds away from C-5. On the other hand, the hydroxyl proton at δ11.30 brs (OH-6) showed also correlation with the carbon at δ109.43 (C-3), which is also of five bonds distance. However, the only chance for this to happen is that tetillapyrone does not exist as only one compound (5) but instead as two compounds (5 and 6). Thus the correlations observed in the HMBC spectrum were due to the methyl protons (δ1.78s) with the carbons at δ150.54 (C-2 of compound 6) and δ163.85 (C-2 of compound 5), respectively. This also explains the correlation observed between the hydroxyl proton at δ11.30 brs (OH-2) and the carbon at δ109.43 (C-3) of compound 6.

Table 1.
1H and 13C NMR data (500 MHz, DMSO) of tetillapyrone (5 and 6).
It is interesting to note that in the structure previously proposed for nortetillapyrone (8), the positions of the hydroxyl and carbonyl groups of the pyrone moiety were opposite to those found in the structure 5 of tetillapyrone. Though, the HMBC spectrum showed strong correlation between H-3 (δ5.62) and the carbon at δ163.18, like that in structure 5 of tetillapyrone, the correlation between H-3 and the hydroxyl proton at δ 11.23 brs observed in the COSY spectrum, has led to the conclusion that nortetillapyrone should have the structure 8. In this structure, the chemical shift values of the carbonyl carbon (δ150.48) and the enolic carbon (δ163.18) were opposite to those found in tetillapyrone (5). However, there is no satisfactory explanation for this incongruence. The only possibility is, like in the case of tetillapyrone, nortetillapyrone also exists, in solution, as two compounds (7 and 8). In this circumstance, the correlation observed in HMBC spectrum (Table 2) between H-4 (δ 5.52d) and the carbon at δ163.18 (C-2) was due to structure 7, while the correlation between H-4 (δ 5.52d) and the hydroxyl group at δ11.23 brs (OH-2), observed in the COSY spectrum, was due to structure 8.
Table 2.
1H and 13C NMR data (500 MHz, DMSO) of nortetillapyrone (7 and 8)
Since structures 5 and 6 of tetillapyrone are tautomers, they can exist in equilibrium in the solution. In the same way, structures 7 and 8 are also tautomers of nortetllapyrone and thus can also exist in equilibrium in the solution (Figure 2).
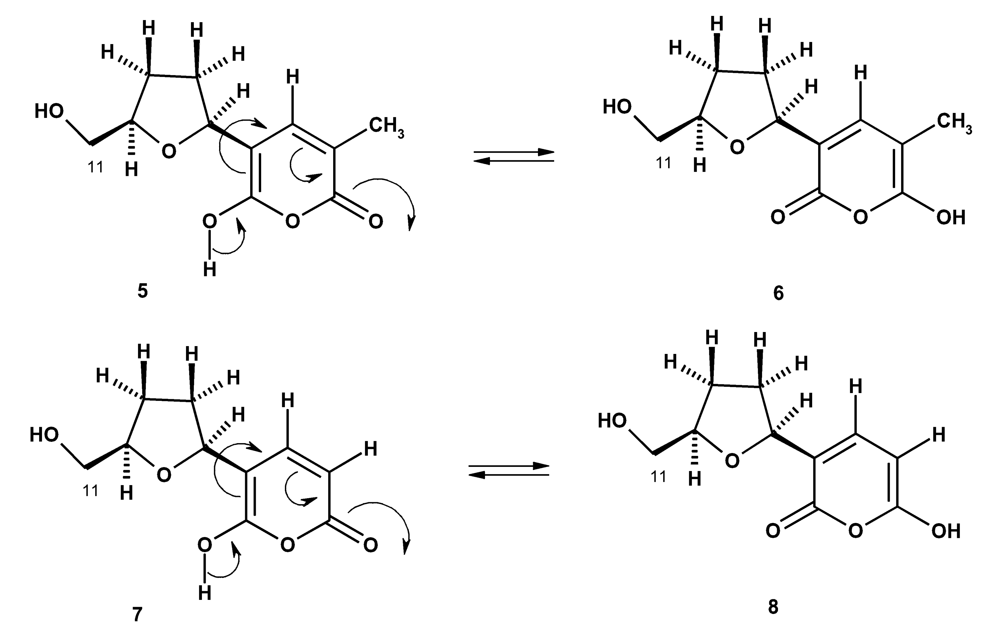
Figure 2.
Tautomerization of tetillapyrone (5, 6) and nortetillapyrone (6, 7).
The structure of compound 1 was identified as maleimide-5-oxime by spectroscopic and HRMS data. EIHRMS of compound 1 gave the molecular ion peak at 122.02723, indicating its molecular formula as C4H4N2O2 (cald.122.02728). The 1H and 13C NMR spectra of compound 1 were very simple, showing the proton and chemical shift values very similar to those observed for 3-methylmaleimide-5-oxime, previously isolated from the marine sponge Pseudoceratina purpurea by our group [12]. However, the 1HNMR of compound 1 showed two doublets of two coupled protons at δ 7.40d (J=7.6 Hz) and 5.45d (J=7.6 Hz) instead of the signals of the vinylic methyl protons (δ 1.73d, J=0.9Hz) and a vinylic proton (δ 7.2.6d, J=0.9Hz).
In order to investigate the biological activity of the constituents of the two marine sponges, maleimide-5-oxime (1), p-hydroxybenzaldehyde (3), phenylacetic acid (4), tetillapyrone (5 and 6), and nortetillapyrone (7 and 8) were evaluated for their growth inhibitory effect against seven yeasts (Candida albicans, C. glabata, C. dubliniensis, C. tropicalis, C. krusei, C. parapsilosis, Cryptococcus neoformans), three non dermatophyte filamentous fungi (Aspergillus fumigatus, A. flavus, A. niger) and five dermatophyte filamentous fungi (Microsporum gypseum, M. canis, Trichophyton mentagrophytes, T. rubrum and Epidermophyton floccosum). Maleimide-5-oxime (1), p-hydroxybenzaldehyde (3), phenylacetic acid (4) and tetillapyrone (5 and 6) were found to be inactive against all the test organisms (MIC > 250 μg/mL). Among the compounds tested, only nortetillapyrone (7, 8) showed growth inhibitory effect against tested fungal strains. Interestingly, some selectivity was observed for this compound. It showed no activity against Candida albicans, C. krusei and C. parapsilosis even at maximum concentration tested (250 μg/mL). However, nortetillapyrone showed some activity against C. tropicalis (250 μg/mL) and good activity against C. glabrata (62.5–31.25 μg/mL), C. dubliniensis (62.5 μg/mL) and Cryptococcus neoformans (31.25 μg/mL). In Addition, this compound exhibited significant antifungal activity against dermatophytes with the MIC values ranging from 31.25 to 125 μg/mL. It was more active against Microsporum canis (31.25 μg/mL) and less active against Trichophyton rubrum (62.5–125 μg/mL). Nortetillapyrone showed no activity against Aspergillus species even at maximum concentration tested (250 μg/mL).
Maleimide-5-oxime (1), tetillapyrone (5 and 6), and nortetillapyrone (7 and 8) were also evaluated for their capacity to inhibit the in vitro growth of three human tumor cell lines, namely the estrogen-dependent ER(+) MCF-7 (human breast adenocarcinoma), the estrogen-independent ER(−) MDA-MB-231 (human breast adenocarcinoma) and NCI-H460 (non-small cell lung cancer). Maleimide-5-oxime (1) and tetillapyrone (5 and 6) were found to be inactive against all the cell lines tested (GI50 > 200 μM) while nortetillapyrone (7 and 8) only weakly inhibited the growth of estrogen-independent ER (−) MDA-MB-231 cell line (GI50 = 155.7 μM) but was inactive against the estrogen-dependent ER (+) MCF-7 and NCI-H460 cell lines (GI50 > 200 μM).
3. Conclusion
As most antifungal compounds from marine origin are cytotoxic, consequently they have not generally been considered promising antifungal agents for clinical application [6]. Though many secondary metabolites from marine source were reported for their antifungal activity, their cytotoxic effect has not been measured. From this point of view, nortetillapyrone showed a very interesting biological activity profile. This compound not only selectively inhibits the growth of dermatophytes but also possesses no cytotoxicity against the tumor cell lines. Consequently, the structural feature of nortetillapyrone could serve as a model for development of a potential antifungal agent with therapeutic value.
4. Experimental
4.1. General
1H and 13C NMR spectra were recorded at ambient temperature in CDCl3 and DMSO on a Bruker AMC instrument operating at 300.12 and 75.47 MHz, respectively or a Bruker DRX instrument operating at 500, respectively 125 MHz. EI mass spectra were measured on a Hitachi Perkin-Elmer RMV-6M instrument. HRMS spectra were measured on a Kratos concept II 2 sector mass spectrometer. The accelerating voltage was 8 kV. Si gel for chromatography was silica gel 60 (0.2–0.5 mm Merck) for analytical work and for preparative work TLC silica gel 60 GF 254 Merck.
4.2. Animal material
Haliclona cymaeformis (Esper, 1794) order Haplosclerida, family Chalinidae, was collected by Scuba diving in the Gulf of Thailand near Koh Chang (2 meter depth), Trad Province in March 2004. Haliclona baeri (Wilson, 1952) was also collected by Scuba diving in the in the coastal area of Chonburi Province (3 meter depth). The sponge materials were identified by Dr. Sumaitt Putchakarn and the vouchers of the specimens, registered as BIMS-I 953 and BIMS-I 1766, respectively were deposited at Bangsaen Institute of Marine Science (BIMS), Burapha University, Bangsaen, Chonburi, Thailand. The materials were frozen immediately at −20ºC prior to extraction.
4.3. Extraction, isolation and characterization of the constituents of Haliclona baeri
The sample (1.3 kg net weight) was thawed, homogenized with EtOH (3 L), allowed to stand for 24 h in a dark chamber and filtered. The residue on the filter paper was again extracted with EtOH (2 × 3L), the aqueous alcoholic extracts were combined, evaporated at reduced pressure to ca. 500 ml and extracted with EtOAc (3 × 500mL). The EtOAc extracts were combined and concentrated at reduced pressure to give the crude EtOAc extract. This (10 g) was chromatographed over a Si gel column (120g) and eluted with petrol-CHCl3 and CHCl3-Me2O, 350 ml fractions being eluted as follows. Frs 1–62 (petrol-CHCl3, 1:1), 63–78 (petrol-CHCl3, 3:7), 79–95 (petrol-CHCl3, 1:9), 96–182 (CHCl3-Me2O, 4:1), 183–206 (CHCl3- Me2O, 3:2). Frs 101–114 were combined (107.8 mg) and purified by TLC (Si gel, CHCl3-Me2O–HCO2H, 5:5:0.1) to give 8.6 mg of maleimide 5-oxime (1). Frs 125–198 were combined (185.6 mg) and purified by TLC (Si gel, CHCl3-MeOH-HCO2H, 93:7:1) to give 13.1 mg 3,4-dihydroxybenzoic acid (2) and 8.0 mg of tetillapyrone (5and 6).
4.4. Extraction, isolation and characterization of the constituents of Haliclona cymaeformis
The sample of Haliclona cymaeformis (7.6 kg net weight) was thawed, homogenized with EtOH (10L), allowed to stand for 24 h in the dark chamber and filtered. The residue on the filter paper was again extracted with EtOH (2 × 10L), the aqueous alcoholic extracts were combined, evaporated at reduced pressure to give ca. 700 ml and extracted with EtOAc (3 × 1L). The EtOAc extracts were combined and concentrated at reduced pressure to give a crude EtOAc extract (21g). This extract was applied to flash chromatography column of Si Gel (150 g) and eluted with CHCl3 (frs. 1–5; 5 × 200 mL), CHCl3-MeOH,, 49:1 (frs 6–15; 10 × 200 mL), CHCl3-MeOH,, 19:1 (frs16–20; 5 × 200 mL), CHCl3-MeOH,, 9:1 (frs 21–22; 2 × 500 mL), EtOAc (frs 23–24; 2 × 500mL) and MeOH (frs 25–26; 2 × 500mL). Frs 18–24 were combined (3.1g), applied on the flash chromatography column of ODS-A gel (35g) and eluted with mixture of MeOH-H2O; 7:3 (sfr 1; 200mL), MeOH-H2O; 4:1 (sfr 2; 200mL), MeOH-H2O; 9:1 (sfr 3; 2 × 100mL), MeOH (sfr 4; 400mL), CHCl3-MeOH-H2O; 14:6:1 (sfr 5, 400mL). Sfr 1 (156 mg) was purified by TLC Si gel, CHCl3-MeOH-HCO2H, 19:1:0.1) to give 3.5 mg of p-hydroxybenzaldehyde (3) and 15 mg of tetillapyrone (5 and 6). Sfr 5 (103 mg) was purified by TLC (Si gel, CHCl3-MeOH-HCO2H, 19:1:0.1) to give 7 mg of phenylacetic acid (4) and 17 mg of nortetillapyrone (7 and 8).
Maleimide-5-oxime (1): yellow viscous mass; EI HRMS 122.02723 (M+),cald for C4H4N2O2, 122.02728.1HNMR (300 MHz, DMSO): δ11.03 brs (−NH), 10.86 brs (=N-OH), 7.40d (J=7.6 Hz, H-4), 5.45d (J=7.6 Hz, H-3), 13CNMR (75.47 MHz, DMSO): δ164.37 (CO-2), 151.54 (C-5), 142.22 (CH-4), 100.24 (CH-3).
4.5. Biological Activity
5.5.1. Cell Growth Assay
The effect of compounds on the growth of human tumor cell lines were evaluated according to the procedure adopted by National Cancer Institute (NCI, USA) for the in vitro anticancer drug discovery screen which uses the protein-binding dye sulforhodamine B (SRB) to assess cell growth inhibition [13, 14] and according to the method described by Kijjoa et al. [15].
5.5.2. Antifungal Activity Determination
Antifungal activity of maleimide-5-oxime (1), p-hydroxybenzaldehyde (3), phenylacetic acid (4), tetillapyrone (5 and 6) and nortetillapyrone (7 and 8) was evaluated against Candida, Aspergillus and dermatophyte strains: three yeasts clinical strains, one of Cryptococcus neoformans PH1, one of Candida dubliniensis PH10 and one of C. glabrata D10R, as well as, four American Type Culture Collection (ATCC) type strains (C. albicans ATTC 10231, C. tropicalis ATCC 13803, C. krusei ATCC 6258 and C. parapsilosis ATCC 90018); one Aspergillus clinical strain (A. flavus F44), as well as, two ATCC type strains (A. niger ATCC 16404, and A. fumigatus ATCC 46645); five dermatophyte clinical strains (Microsporum canis FF1, M. gypseum FF3, Trichophyton rubrum FF5, T. mentagrophytes FF7 and Epidermophyton floccosum FF9).
The fungal isolates were identified by standard microbiology methods and stored on Sabouraud Dextrose broth with glycerol at −70ºC and subcultured in Sabouraud-Chloramphenicol agar. Minimal Inhibitory Concentration (MIC) was determined by a microdilution method according to the CLSI reference method M27-A2 for yeasts [16], and M38-A for filamentous fungi [17]. The test was performed in sterile 96 flat-bottom well microtitration plates and the medium used was MOPS-buffered RPMI 1640. Briefly, MICs were determined for all yeasts strains with the adjusted final inoculum of 1–4 × 104 CFU/mL, prepared from a 24–48 hours culture. With filamentous fungi (except for Epidermophyton floccosum), the final inoculum used was 4–8 × 104CFU/mL, prepared from a sporulated culture. For Epidermophyton floccosum, the final inoculum was 8 × 104 to 1.6 × 105 CFU/mL. Two fold serial dilutions of compounds in DMSO, ranging from 250 to 15.125 μg/mL, were tested. Fungal growth controls, with and without DMSO, were included. The final DMSO concentration was 2%, and did not affect the fungal growth. In addition, reference antifungal compounds, fluconazole (Pfizer, United Kingdom) for yeasts and dermatophytes or amphotericin B (Sigma) for Aspergillus, were used as the standard antifungal drugs. Twofold serial dilutions ranging from 128 to 0.25 μg/mL for fluconazole and 16 to 0.063 μg/mL for amphotericin B were used. Plates were incubated at 30ºC for 48 hours for Candida species, 72 hours for Cryptococcus neoformans, 4–5 days for Aspergillus species and 8 days for dermatophytes. Quality control determinations of the MIC of fluconazole were ensured by testing C. parapsilosis ATCC 90018. The results were within the recommended limits. All determinations were performed in duplicate and three independent experiments lead to concordant results.
Acknowledgements
This work was supported by FCT-Fundação para a Ciência e Tecnologia (Project POCI/MAR/58114/2004) and Unidade de ID 226/94; FEDER and POCI, and GRICES. Work in Thailand was supported by the Thailand Research Fund (No. TRG 4780003). We thank Dr. Sumaitt Puchakarn, BIMS, Burapha University, Thailand, for collection and identification of the sponge materials and Dr. Graham Eaton, Department of Chemistry, University of Leicester, UK for HRMS.
- Sample Availability: Available from the author.
References
- Li, HY; Matsunaga, S; Fusetani, N. Antifungal metabolites from marine sponges. Curr Org Chem 1998, 2, 649–682. [Google Scholar]
- Richardson, MD. Changing patterns and trends in systematic fungal infections. J Antimicrobial Chemother 2005, 56, i5–i11. [Google Scholar]
- Gupta, AK; Tu, LQ. Dermatophytes: Diagnosis and treatment. J Am Acad Dermatol 2006, 54, 1050–1055. [Google Scholar]
- Johnson, L. Dermatophytes-the skin eaters. Mycologist 2003, 17, 147–149. [Google Scholar]
- Sundriyal, S; Sharma, RK; Jain, R. Current advances in antifungal targets and drug development. Curr Med Chem 2006, 13, 1321–1335. [Google Scholar]
- Donia, M; Hartmann, M. Marine natural products and their potential applications as anti-infective agents. THE LANCET Infectious Disease 2003, 3, 338–348. [Google Scholar]
- Faulkner, DJ. Marine Natural Products. Nat Prod Rep 1993, 10, 497–539. [Google Scholar]
- Rashid, MA; Gustafson, KR; Boswell, JL; Boyd, MR. Haligramides A and B, two new cytotoxic hexapeptides from the marine sponge Haliclona nigra. J Nat Prod 2000, 63, 956–959. [Google Scholar]
- Fahy, E; Molinski, TF; Harper, MK; Sulliwan, BW; Faulkner, DJ. Haliclonadiamine: an antimicrobial alkaloid from the sponge Haliclona sp. Tetrahedron Lett 1988, 29, 3427–3428. [Google Scholar]
- Clark, RJ; Garson, MJ; Hooper, JNA. Halaminols A-C, three new alkyl amino alcohol from the sponge Haliclona sp. J Nat Prod 2001, 64, 1568–1571. [Google Scholar]
- Watanadilok, R; Sonchaeng, P; Kijjoa, A; Damas, AM; Gales, L; Silva, AMS; Herz, W. Tetillapyrone and nortetillapyrone, two unusual hydroxypyran-2-ones from the marine sponge Tetilla japonica. J Nat Prod 2001, 64, 1056–1058. [Google Scholar]
- Kijjoa, A; Bessa, J; Wattanadilok, R; Sawangwong, P; Nascimento, MSJ; Pedro, M; Silva, AMS; Eaton, G; van Soest, R; Herz, W. Dibromotyrosine derivatives, a maleimide, aplysamine -2 and other constituents of the marine sponge Pseudoceratina purpurea. Zeitschrift fur Naturforschung 2005, 60b, 904–908. [Google Scholar]
- Monks, A; Scudiero, D; Skehan, P; Shoemaker, R; Paul, K; Vistica, D; Hose, C; Langley, J; Cronise, P; Vaigro-Wolff, A; Gray-Goodrich, M; Campbell, H; Mayo, J; Boyd, M. Feasibility of a high-flux anticancer drug screen using a diverse panel of cultured human tumor cell lines. J Natl Cancer Inst 1991, 83, 757–776. [Google Scholar]
- Skehan, P; Storeng, R; Scudiero, D; Monks, A; McMahon, J; Vistica, D; Warren, JT; Bokesch, H; Kenny, S; Boyd, MR. New calorimetric cytotoxic assay for anticancer drug screening. J Natl Cancer Inst 1990, 82, 1107–1112. [Google Scholar]
- Kijjoa, A; Wattanadilok, R; Campos, N; Nascimento, MSJ; Pinto, M; Herz, W. Anticancer activity Evaluation of Kuanoniamines A and C isolated from the marine sponge Oceanapia sagittaria:collected from the Gulf of Thailand. Mar Drugs 2007, 5, 6–22. [Google Scholar]
- National Committee for Clinical Laboratory Standards, 2002; Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts: approved standard Document M27-A2; Wayne, PA; National Committee for Clinical Laboratory Standards.
- National Committee for Clinical Laboratory Standards, 2002; Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi: approved standard Document M38-A; Wayne, PA; National Committee for Clinical Laboratory Standards; Sample Availability: Available from the author.