Mycosporine-Like Amino Acids and Marine Toxins - The Common and the Different

Abstract
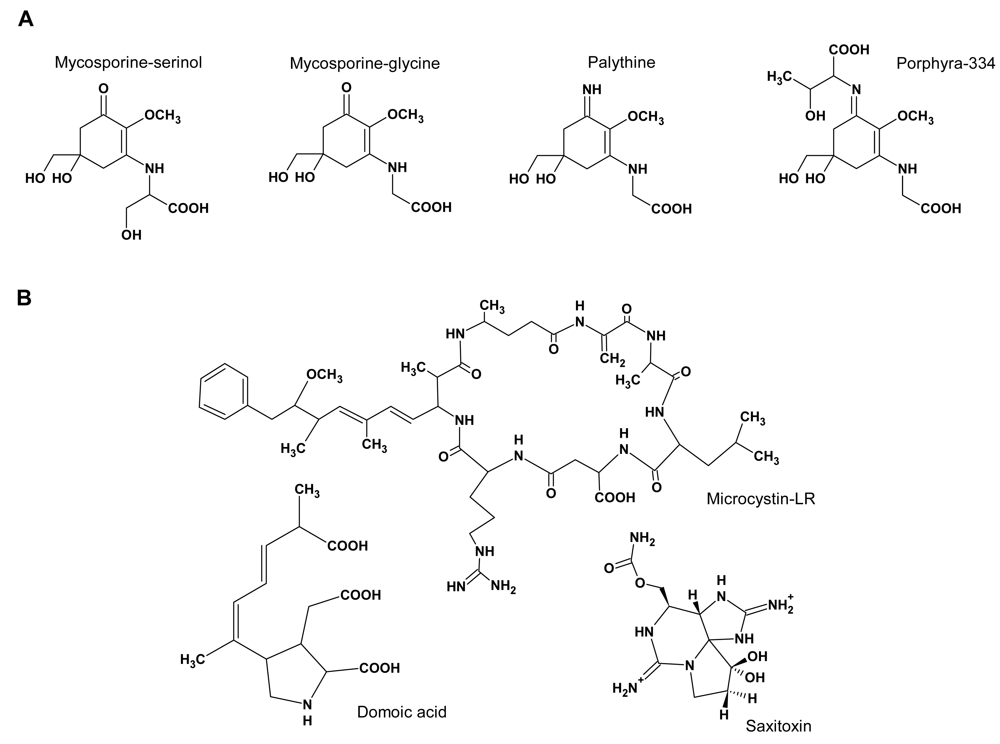
:Introduction
Functions of MAAs and toxins
Co-occurrence of MAAs and toxins in cyanobacteria and eukaryotic algae
Cyanobacteria
Dinoflagellates
Other algae
Influence of environmental factors on toxin and MAA production
Effects of radiation
Effects of nutrients
Other factors
Distribution of MAAs and algal toxins in food webs
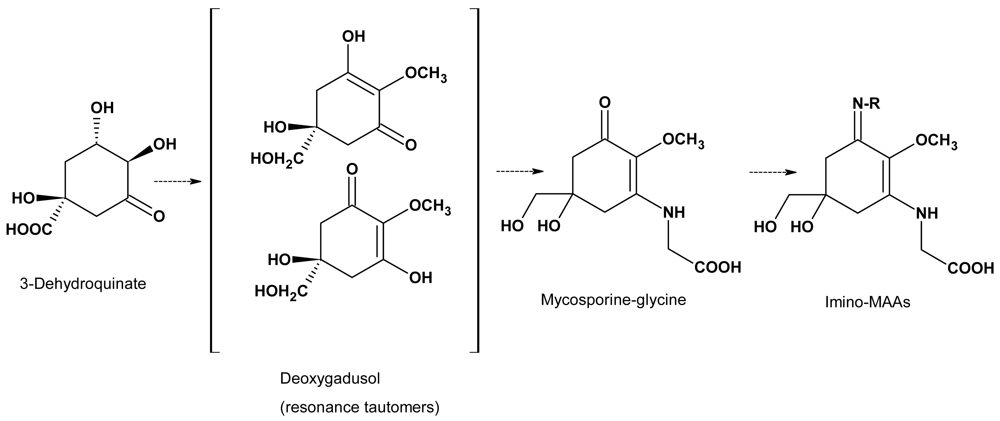
Parallels in the biosynthesis of MAAs and marine toxins
Conclusive remarks
- Samples Availability: Available from the authors.
References
- Lebar, MD; Heimbegner, JL; Baker, BJ. Cold-water marine natural products. Nat Prod Rep 2007, 24, 774–97. [Google Scholar]
- Blunt, JW; Copp, BR; Hu, W-P; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat Prod Rep 2007, 24, 31–86. [Google Scholar]
- Cardozo, KHM; Guaratini, T; Barros, MP; Falcão, VR; Tonon, AP; Lopes, NP; Campos, S; Torres, MA; Souza, AO; Colepicolo, P; Pinto, E. Metabolites from algae with economical impact. Comp Biochem Physiol C Toxicol Pharmacol 2007, 146, 60–78. [Google Scholar]
- Takano, S; Nakanishi, A; Uemura, D; Hirata, Y. Isolation and structure of a 334 nm UV-absorbing substance, porphyra-334 from the red alga Porphyra tenera Kjellman. Chem Lett 1979, 8, 419–20. [Google Scholar]
- Nakamura, H; Kobayashi, J; Hirata, Y. Separation of mycosporine-like amino acids in marine organisms using reverse-phase high performance liquid chromatography. J Chromatogr 1982, 250, 113–18. [Google Scholar]
- Bandaranayake, WM. Mycosporines: are they nature’s sunscreens? Nat Prod Rep 1998, 15, 159–72. [Google Scholar]
- Favre-Bonvin, J; Bernillon, J; Salin, N; Arpin, N. Biosynthesis of mycosporines: mycosporine glutaminol in Trichothecium roseum. Phytochemistry 1987, 26, 2509–14. [Google Scholar]
- Shick, JM; Romaine-Lioud, S; Ferrier-Pagès, C; Gattuso, J-P. Ultraviolet-B radiation stimulates shikimate pathway-dependent accumulation of mycosporine-like amino acids in the coral Stylophora pistillata despite decrease in its population of symbiotic dinoflagellates. Limnol Oceanogr 1999, 44, 1667–82. [Google Scholar]
- Arai, T; Nishijima, M; Adachi, K; Sano, H. Isolation and structure of a UV absorbing substance from the marine bacterium Micrococcus sp. AK-334; Marine Biotechnology Institute: Tokyo, Japan, 1992; pp. 88–94. [Google Scholar]
- Dunlap, WC; Shick, JM. Ultraviolet radiation-absorbing mycosporine-like amino acids in coral reef organisms: a biochemical and environmental perspective. J Phycol 1998, 34, 418–30. [Google Scholar]
- Shick, JM; Dunlap, WC. Mycosporine-like amino acids and related gadusols: biosynthesis, accumulation, and UV-protective functions in aquatic organisms. Annu Rev Physiol 2002, 64, 223–62. [Google Scholar]
- Starcevic, A; Akthar, S; Dunlap, WC; Shick, JM; Hranueli, D; Cullum, J. Enzymes of the shikimic acid pathway encoded in the genome of a basal metazoan, Nematostella vectensis, have microbial origins. PNAS 2008, 105, 2533–37. [Google Scholar]
- Klisch, M; Sinha, RP; Richter, PR; Häder, D-P. Mycosporine-like amino acids (MAAs) protect against UV-B-induced damage in Gyrodinium dorsum Kofoid. J Plant Physiol 2001, 158, 1449–54. [Google Scholar]
- Adams, NL; Shick, JM. Mycosporine-like amino acids provide protection against ultraviolet radiation in eggs of the green sea urchin Strongylocentrotus droebachiensis. Photochem Photobiol 1996, 64, 149–58. [Google Scholar]
- Neale, PJ; Banaszak, AT; Jarriel, CR. Ultraviolet sunscreens in Gymnodinium sanguineum (Dinophyceae): mycosporine-like amino acids protect against inhibition of photosynthesis. J Phycol 1998, 34, 928–38. [Google Scholar]
- Oren, A. Mycosporine-like amino acids as osmotic solutes in a community of halophilic cyanobacteria. Geomicrobiol J 1997, 14, 231–40. [Google Scholar]
- Dunlap, WC; Yamamoto, Y. Small-molecule antioxidants in marine organisms: antioxidant activity of mycosporine-glycine. Comp Biochem Physiol B Biochem Mol Biol 1995, 112, 105–14. [Google Scholar]
- Schmid, D; Schürch, C; Zülli, F. UV-A sunscreen from red algae for protection against premature skin aging. Cosmet Toilet Manufact Worldwide 2004, Yearbook 2004. 139–43. [Google Scholar]
- Torres, A; Enk, CD; Hochberg, M; Srebnik, M. Porphyra-334, a potential natural source for UVA protective sunscreens. Photochem Photobiol Sci 2006, 5, 432–35. [Google Scholar]
- Bultel-Poncé, V; Felix-Theodore, F; Sarthon, C; Ponge, J-F; Bodo, B. New pigments from the terrestrial cyanobacterium Scytonema sp. collected on the Mitaraka Inselberg, French Guyana. J Nat Prod 2004, 67, 678–81. [Google Scholar]
- Büdel, B; Karsten, U; Garcia-Pichel, F. Ultraviolet-absorbing scytonemin and mycosporine-like amino acids derivatives in exposed rock-inhabiting cyanobacterial lichens. Oecologia 1997, 112, 165–72. [Google Scholar]
- Bjerke, JW; Lerfall, K; Elvebakk, A. Effects of ultraviolet radiation and PAR on the content of usnic and divaricatic acids in two arctic-alpine lichens. Photochem Photobiol Sci 2002, 1, 678–85. [Google Scholar]
- Proteau, PJ; Gerwick, WH; Garcia-Pichel, F; Castenholz, R. The structure of scytonemin, an ultraviolet sunscreen pigment from the sheaths of cyanobacteria. Experientia 1993, 49, 825–29. [Google Scholar]
- Cocchietto, M; Skert, N; Nimis, PL; Sava, G. A review on usnic acid, an interesting natural compound. Naturwissenschaften 2002, 89, 137–46. [Google Scholar]
- Garcia-Pichel, F; Sherry, ND; Castenholz, RW. Evidence for an ultraviolet sunscreen role of the extracellular pigment scytonemin in the terrestrial cyanobacterium Chlorogloeopsis sp. Photochem Photobiol 1992, 56, 17–23. [Google Scholar]
- Anon. Marine Biotoxins; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004. [Google Scholar]
- Daugbjerg, N; Hansen, G; Larsen, J; Moestrup, O. Phylogeny of some of the major genera of dinoflagellates based on ultrastructure and partial LSU rDNA sequence data, including the erection of three new genera of unarmoured dinoflagellates. Phycologia 2000, 39, 302–17. [Google Scholar]
- Van Dolah, FM. Marine algal toxins: origins, health effects, and their increased occurrence. Environ Health Perspect 2000, 108, 133–41. [Google Scholar]
- Chorus, I; Bartram, J. Toxic cyanobacteria in water, a guide to their public health consequences, monitoring and management; E & FN Spon: London, UK, 1999. [Google Scholar]
- Vasconcelos, V. Cyanobacteria toxins: diversity and ecological effects. Limnetica 2001, 20, 45–58. [Google Scholar]
- Yuan, YV; Walsh, NA. Antioxidant and antiproliferative activities of extracts from a variety of edible seaweeds. Food Chem Toxicol 2006, 44, 1044–50. [Google Scholar]
- Gao, K. Chinese studies on the edible blue-green alga, Nostoc flagelliforme: a review. J Appl Phycol 1998, 10, 37–49. [Google Scholar]
- Chioccara, F; Miscuraca, G; Novellino, E; Prota, G. Occurrence of two new mycosporine-like amino acids, mytilins a and b in the edible mussel, Mytilus galloprovincialis. Tetrahedron Lett 1979, 20, 3181–82. [Google Scholar]
- Yakovleva, I; Takemura, A; Bhagooli, R; Hidaka, M. Differential susceptibility to oxidative stress of two scleractinian corals: antioxidant functioning of mycosporine-glycine. Comp Biochem Physiol B 2004, 139, 721–30. [Google Scholar]
- Suh, H-J; Lee, H-W; Jung, J. Mycosporine glycine protects biological systems against photodynamic damage by quenching singlet oxygen with a high efficiency. Photochem Photobiol 2003, 78, 109–13. [Google Scholar]
- Oren, A; Gunde-Cimerman, N. Mycosporines and mycosporine-like amino acids: UV protectants or multipurpose secondary metabolites? FEMS MicrobiolLett 2007, 269, 1–10. [Google Scholar]
- Jokiel, PL; York, HR. Solar ultraviolet photobiology of the reef coral Pocillopora damicornis and symbiotic zooxanthellae. Bull Mar Sci 1982, 32, 301–15. [Google Scholar]
- Shick, JM; Lesser, MP; Dunlap, WC; Stochaj, WR; Chalker, BE; Wu Won, J. Depth-dependent responses to solar ultraviolet radiation and oxidative stress in the zooxanthellate coral Acropora microphthalma. Mar Biol 1995, 122, 41–51. [Google Scholar]
- Tartarotti, B; Laurion, I; Sommaruga, R. Large variability in the concentration of mycosporine-like amino acids among zooplankton from lakes located across an altitude gradient. Limnol Oceanogr 2001, 46, 1546–52. [Google Scholar]
- Karsten, U; Sawall, T; Hanelt, D; Bischof, K; Figueroa, FL; Flores-Moya, A; Wiencke, C. An inventory of UV-absorbing mycosporine-like amino acids in macroalgae from polar to warm-temperate regions. Bot Mar 1998, 41, 443–53. [Google Scholar]
- Conde, FR; Churio, MS; Previtali, CM. The deactivation pathways of the excited-states of the mycosporine-like amino acids shinorine and porphyra-334 in aqueous solution. Photochem Photobiol Sci 2004, 3, 960–67. [Google Scholar]
- Misonou, T; Saitoh, J; Oshiba, S; Tokitomo, Y; Maegawa, M; Inoue, Y; Hori, H; Sakurai, T. UV-absorbing substance in the red alga Porphyra yezoensis (Bangiales, Rhodophyta) block thymine photodimer production. Mar Biotechnol 2003, 5, 194–200. [Google Scholar]
- Suh, H-J; Lee, H-W; Jung, J. Singlet oxygen quenching by deoxygadusol and related mycosporine-like amino acids from phytoplankton Prorocentrum micans. J Photosci 2004, 11, 77–81. [Google Scholar]
- Macías, FA; Galindo, JLG; García-Díaz, M; Galindo, JCG. Allelopathic agents from aquatic ecosystems: potential biopesticides models. Phytochem Rev 2008, 7, 155–78. [Google Scholar]
- Ajuzie, CC. Palatability and fatality of the dinoflagellate Prorocentrum lima to Artemia salina. J Appl Phycol 2007, 19, 513–19. [Google Scholar]
- Pflugmacher, S. Possible allelopathic effects of cyanotoxins, with reference to microcystin-LR, in aquatic ecosystems. Environ Toxicol 2002, 17, 407–13. [Google Scholar]
- Hu, Z-Q; Liu, Y-D; Li, D-H. Physiological and biochemical analyses of microcystin-RR toxicity to the cyanobacterium Synechococcus elongatus. Environ Toxicol 2004, 19, 571–77. [Google Scholar]
- LeBlanc, S; Pick, FR; Aranda-Rodriguez, R. Allelopathic effects of the toxic cyanobacterium Microcystis aeruginosa on duckweed, Lemna gibba L. Environ Toxicol 2005, 20, 67–73. [Google Scholar]
- Selander, E; Thor, P; Toth, G; Pavia, H. Copepods induce paralytic shellfish toxin production in marine dinoflagellates. Proc R Soc B Biol Sci 2006, 273, 1673–80. [Google Scholar]
- Maldonado, MT; Hughes, MP; Rue, EL. The effect of Fe and Cu on growth and domoic acid production by Pseudo-nitzschia multiseries and Pseudo-nitzschia australis. Limnol Oceanogr 2002, 47, 515–26. [Google Scholar]
- Carreto, JI; Carignan, MO; Montoya, NG. Comparative studies on mycosporine-like amino acids, paralytic shellfish toxins and pigment profiles of the toxic dinoflagellates Alexandrium tamarense, A. catenella and A. minutum. Mar Ecol Prog Ser 2001, 223, 49–60. [Google Scholar]
- Gröniger, A; Sinha, RP; Klisch, M; Häder, D-P. Photoprotective compounds in cyanobacteria, phytoplankton and macroalgae - a database. J Photochem Photobiol B Biol 2000, 58, 115–22. [Google Scholar]
- Jeffrey, SW; MacTavish, HS; Dunlap, WC; Vesk, M; Groenewoud, K. Occurrence of UVA- and UVB-absorbing compounds in 152 species (206 strains) of marine microalgae. Mar Ecol Prog Ser 1999, 189, 35–51. [Google Scholar]
- Liu, Z; Häder, DP; Sommaruga, R. Occurrence of mycosporine-like amino acids (MAAs) in the bloom-forming cyanobacterium Microcystis aeruginosa. J Plankton Res 2004, 26, 963–66. [Google Scholar]
- Kurmayer, R; Christiansen, G; Chorus, I. The abundance of microcystin-producing genotypes correlates positively with colony size in Microcystis sp. and determines its microcystin net production in Lake Wannsee. Appl Environ Microbiol 2003, 69, 787–95. [Google Scholar]
- Lyra, C; Laamanen, M; Lehtimäki, JM; Surakka, A; Sivonen, K. Benthic cyanobacteria of the genus Nodularia are non-toxic, without gas vacuoles, able to glide and genetically more diverse than planktonic Nodularia. Int J Syst Evol Microbiol 2005, 55, 555–68. [Google Scholar]
- Koskenniemi, K; Lyra, C; Rajaniemi-Wacklin, P; Jokela, J; Sivonen, K. Quantitative real-time PCR detection of toxic Nodularia cyanobacteria in the Baltic Sea. Appl Environ Microbiol 2007, 73, 2173–79. [Google Scholar]
- Sinha, RP; Ambasht, NK; Sinha, JP; Klisch, M; Häder, D-P. UV-B-induced synthesis of mycosporine-like amino acids in three strains of Nodularia (cyanobacteria). J Photochem Photobiol B Biol 2003, 71, 51–58. [Google Scholar]
- Pereira, P; Onodera, H; Andrinolo, D; Franca, S; Araújo, F; Lagos, N; Oshima, Y. Paralytic shellfish toxins in the freshwater cyanobacterium Aphanizomenon flosaquae, isolated from Montargil reservoir, Portugal. Toxicon 2008, 38, 1689–702. [Google Scholar]
- Smayda, TJ. Harmful algal blooms: their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol Oceanogr 1997, 42, 1137–57. [Google Scholar]
- Babin, M; Cullen, JJ; Roesler, CS; Donaghay, PL; Doucette, GJ; Kahru, M; Scholin, CA; Sieracki, ME; Sosik, HM. New approaches and technologies for observing harmful algal blooms. Oceanography 2005, 18, 210–27. [Google Scholar]
- Bates, SS. Domoic-acid-producing diatoms: another genus added! JPhycol 2000, 36, 978–85. [Google Scholar]
- Carreto, JI; Carignan, MO; Montoya, NG. A high-resolution reverse-phase liquid chromatography method for the analysis of mycosporine-like amino acids (MAAs) in marine organisms. Mar Biol 2005, 146, 237–52. [Google Scholar]
- Helbling, EW; Chalker, BE; Dunlap, WC; Holm-Hansen, O; Villafañe, VE. Photoacclimation of Antarctic marine diatoms to solar ultraviolet radiation. J Exp Mar Biol Ecol 1996, 204, 85–101. [Google Scholar]
- Karsten, U; Sawall, T; Wiencke, C. A survey of the distribution of UV-absorbing substances in tropical macroalgae. Phycol Res 1998, 46, 271–79. [Google Scholar]
- Klisch, M; Häder, D-P. Wavelength dependence of mycosporine-like amino acid synthesis in Gyrodinium dorsum. J Photochem Photobiol B Biol 2002, 66, 60–66. [Google Scholar]
- Kräbs, G; Watanabe, M; Wiencke, C. A monochromatic action spectrum for the photoinduction of the UV-absorbing mycosporine-like amino acid shinorine in the red alga Chondrus crispus. Photochem Photobiol 2004, 79, 515–19. [Google Scholar]
- Portwich, A; Garcia-Pichel, F. A novel prokaryotic UVB photoreceptor in the cyanobacterium Chlorogloeopsis PCC 6912. Photochem Photobiol 2000, 71, 493–98. [Google Scholar]
- Sinha, RP; Sinha, JP; Gröniger, A; Häder, D-P. Polychromatic action spectrum for the induction of a mycosporine-like amino acid in a rice-field cyanobacterium, Anabaena sp. J Photochem Photobiol B Biol 2002, 66, 47–53. [Google Scholar]
- Sinha, RP; Klisch, M; Helbling, EW; Häder, D-P. Induction of mycosporine-like amino acids (MAAs) in cyanobacteria by solar ultraviolet-B radiation. J Photochem Photobiol B Biol 2001, 60, 129–35. [Google Scholar]
- Carreto, JI; Lutz, VA; De Marco, SG; Carignan, MO. Graneli, E, Edler, L, Sundström, B, Anderson, DM, Eds.; Toxic Marine Phytoplankton; Elsevier: New York, NY, USA, 1990. [Google Scholar]
- Leach, CM. Ultraviolet-absorbing substances associated with light-induced sporulation in fungi. Canad J Bot 1965, 43, 185–200. [Google Scholar]
- Kaebernik, M; Brett, AN; Börner, T; Dittmann, E. Light and the transcriptional response of the microcystin biosynthesis gene cluster. Appl Environ Microbiol 2000, 66, 3387–92. [Google Scholar]
- Litchman, E; Neale, PJ; Banaszak, AT. Increased sensitivity to ultraviolet radiation in nitrogen-limited dinoflagellates: photoprotection and repair. Limnol Oceanogr 2002, 47, 86–94. [Google Scholar]
- Peinado, NK; Díaz, RTA; Figueroa, FL; Helbling, EW. Ammonium and UV radiation stimulate the accumulation of mycosporine-like amino acids in Porphyra columbina (Rhodophyta) from Patagonia, Argentina. J Phycol 2004, 40, 248–59. [Google Scholar]
- Touzet, N; Franco, JM; Raine, R. Influence of inorganic nutrition on growth and PSP toxin production of Alexandrium minutum (Dinophyceae) from Cork Harbour, Ireland. Toxicon 2007, 50, 106–19. [Google Scholar]
- Leong, SCY; Murata, A; Nagashima, Y; Taguchi, S. Variability in toxicity of the dinoflagellate Alexandrium tamarense in response to different nitrogen sources and concentrations. Toxicon 2004, 43, 407–15. [Google Scholar]
- Pan, Y; Subba Rao, DV; Mann, KH; Li, WKW; Harrison, WG. Effects of silicate limitation on production of domoic acid, a neurotoxin, by the diatom Pseudo-nitzschia multiseries. II. Continuous culture studies. Mar Ecol Prog Ser 1996, 273, 235–43. [Google Scholar]
- Pan, Y; Subba Rao, DV; Mann, KH; Brown, RG; Pocklington, R. Effects of silicate limitation on production of domoic acid, a neurotoxin, by the diatom Pseudo-nitzschia multiseries. I. Batch culture studies. Mar Ecol Prog Ser 1996, 131, 225–33. [Google Scholar]
- Pan, Y; Subba Rao, DV; Mann, KH. Changes in domoic acid production and cellular chemical composition of the toxigenic diatom Pseudonitzschia multiseries under phosphate limitation. J Phycol 1996, 32, 371–81. [Google Scholar]
- Lundholm, N; Hansen, PJ; Kotaki, Y. Effect of pH on growth and domoic acid production by potentially toxic diatoms of the genera Pseudonitzschia and Nitzschia. Mar Ecol Prog Ser 2004, 273, 1–15. [Google Scholar]
- Teegarden, GJ; Cemballa, AG. Grazing of toxic dinoflagellates, Alexandrium spp., by adult copepods of coastal Maine: implications for the fate of paralytic shellfish toxins in marine food webs. J Exp Mar Biol Ecol 1996, 196, 145–76. [Google Scholar]
- Karentz, D; McEuen, FS; Land, MC; Dunlap, WC. A survey of mycosporine-like amino acid compounds in Antarctic marine organisms: potential protection from ultraviolet exposure. Mar Biol 1991, 108, 157–66. [Google Scholar]
- Carroll, AK; Shick, JM. Dietary accumulation of UV-absorbing mycosporine-like amino acids (MAAs) by the green sea urchin (Strongylocentrotus droebachiensis). Mar Biol 1996, 124, 561–69. [Google Scholar]
- Helbling, EW; Menchi, CF; Villafañe, VE. Bioaccumulation and role of UV-absorbing compounds in two marine crustacean species from Patagonia, Argentina. Photochem Photobiol Sci 2002, 1, 820–25. [Google Scholar]
- Mason, DS; Schafer, F; Shick, JM; Dunlap, WC. Ultraviolet radiation-absorbing mycosporine-like amino acids (MAAs) are acquired from their diet by medaka fish (Oryzias latipes) but not by SKH-1 hairless mice. Comp Biochem Physiol A Mol Integr Physiol 1998, 120, 587–98. [Google Scholar]
- Sieiro, C; Poza, M; de Miguel, T; Villa, T. Genetic basis of microbial carotenogenesis. Int Microbiol 2003, 6, 11–16. [Google Scholar]
- Lane, MP; Nakashima, TT; Vederas, JC. Biosynthetic source of oxygens in griseofulvin. Spin-echo resolution of 18O isotope shifts in 13C NMR spectroscopy. J Am Chem Soc 1982, 104, 913–15. [Google Scholar]
- Portwich, A; Garcia-Pichel, F. Biosynthetic pathway of mycosporines (mycosporine-like amino acids) in the cyanobacterium Chlorogloeopsis sp. strain PCC 6912. Phycologia 2003, 42, 384–92. [Google Scholar]
- Dittmann, E; Neilan, BA; Erhard, M; von Döhren, H; Börner, T. Insertional mutagenesis of a peptide synthetase gene that is responsible for hepatotoxin production in Microcystis aeruginosa PCC 7806. Mol Microbiol 1997, 26, 779–84. [Google Scholar]
- Tillet, D; Dittman, E; Erhard, M; von, Döhren H; Börner, T; Neilan, BA. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: an integrated peptide-polyketide synthetase system. Chem Biol 2000, 7, 753–64. [Google Scholar]
- Nishizawa, T; Ueda, A; Asayama, M; Fujii, K; Harada, K; Ochi, K; Shirai, M. Polyketide synthase gene coupled to the peptide synthetase module involved in the biosynthesis of the cyclic heptapeptide microcystin. J Biochem 2000, 127, 779–89. [Google Scholar]
- Finking, R; Marahiel, M. Biosynthesis of nonribosomal peptides. Annu Rev Microbiol 2004, 58, 453–88. [Google Scholar]



{kind=link}
{kind=link}
| Toxin group1 | Primary target organ in mammals | Cyanobacterial genera2 |
|---|---|---|
| Cyclic peptides | ||
| Microcystins | Liver | Microcystis, Anabaena, Oscillatoria (Planktothrix), Nostoc, Hapalosiphon, Anabaenopsis |
| Nodularin | Liver | Nodularia |
| Alkaloids | ||
| Anatoxin-a | Nerve synapse | Anabaena, Oscillatoria (Planktothrix), Aphanizomenon |
| Anatoxin-a(S) | Nerve synapse | Anabaena |
| Saxitoxins | Nerve axons | Anabaena, Aphanizomenon, Lyngbya, Cylindrospermopsis |
| Cylindrospermopsins | Liver3 | Cylindrospermopsis, Aphanizomenon, Umezakia |
| Lyngbyatoxin-a | Skin, gastro-intestinal tract | Lyngbya |
| Aplysiatoxins | Skin | Lyngbya, Schizothrix, Oscillatoria (Planktothrix) |
| Lipopolysaccharides (LPS) | Potential irritant; affects any exposed tissue | All |
Share and Cite
Klisch, M.; Häder, D.P. Mycosporine-Like Amino Acids and Marine Toxins - The Common and the Different. Mar. Drugs 2008, 6, 147-163. https://doi.org/10.3390/md6020147
Klisch M, Häder DP. Mycosporine-Like Amino Acids and Marine Toxins - The Common and the Different. Marine Drugs. 2008; 6(2):147-163. https://doi.org/10.3390/md6020147
Chicago/Turabian StyleKlisch, Manfred, and Donat P. Häder. 2008. "Mycosporine-Like Amino Acids and Marine Toxins - The Common and the Different" Marine Drugs 6, no. 2: 147-163. https://doi.org/10.3390/md6020147
APA StyleKlisch, M., & Häder, D. P. (2008). Mycosporine-Like Amino Acids and Marine Toxins - The Common and the Different. Marine Drugs, 6(2), 147-163. https://doi.org/10.3390/md6020147