Effects of in vitro Brevetoxin Exposure on Apoptosis and Cellular Metabolism in a Leukemic T Cell Line (Jurkat)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
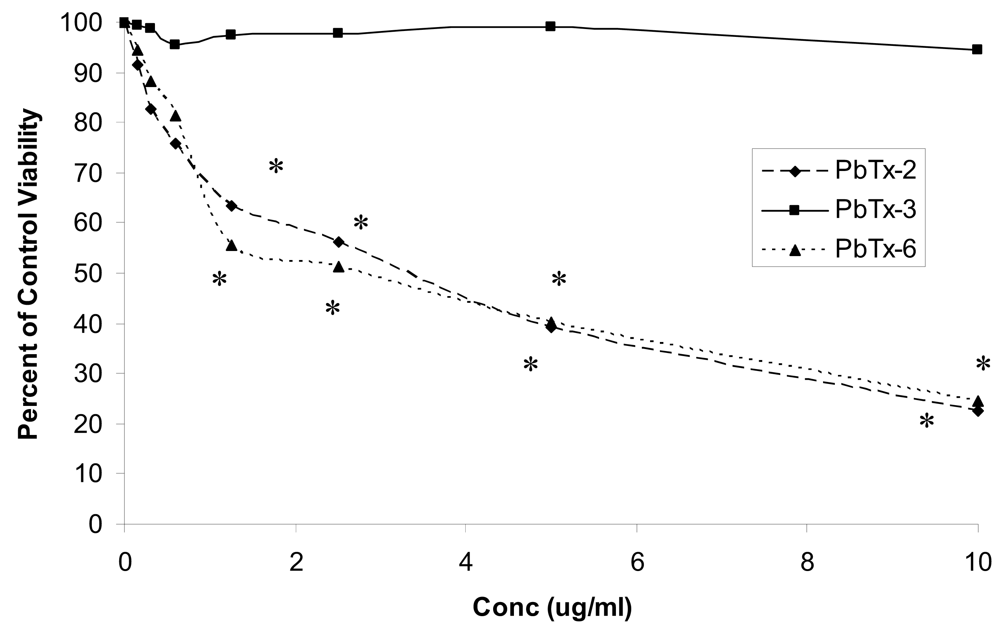
2.1 Effects of toxin on cell viability
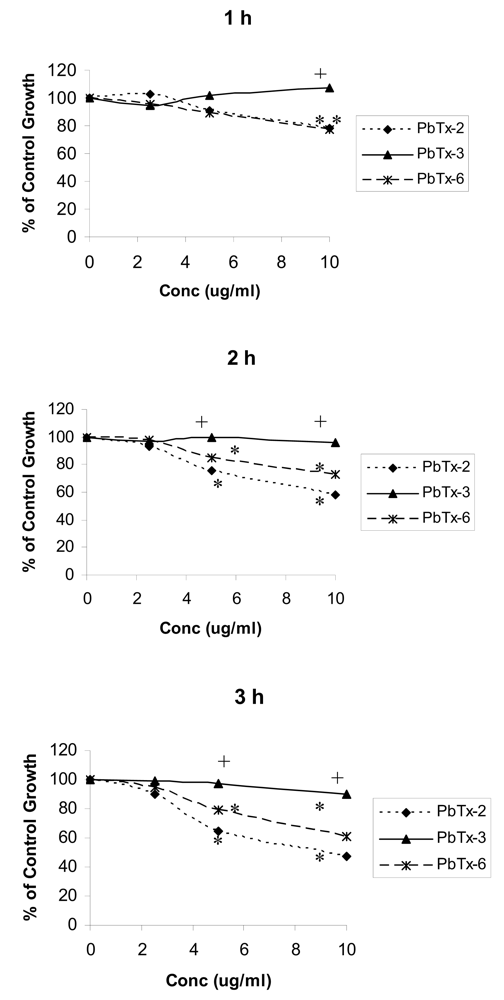
2.2 Effects of brevetoxins on cellular metabolism measured using MTT
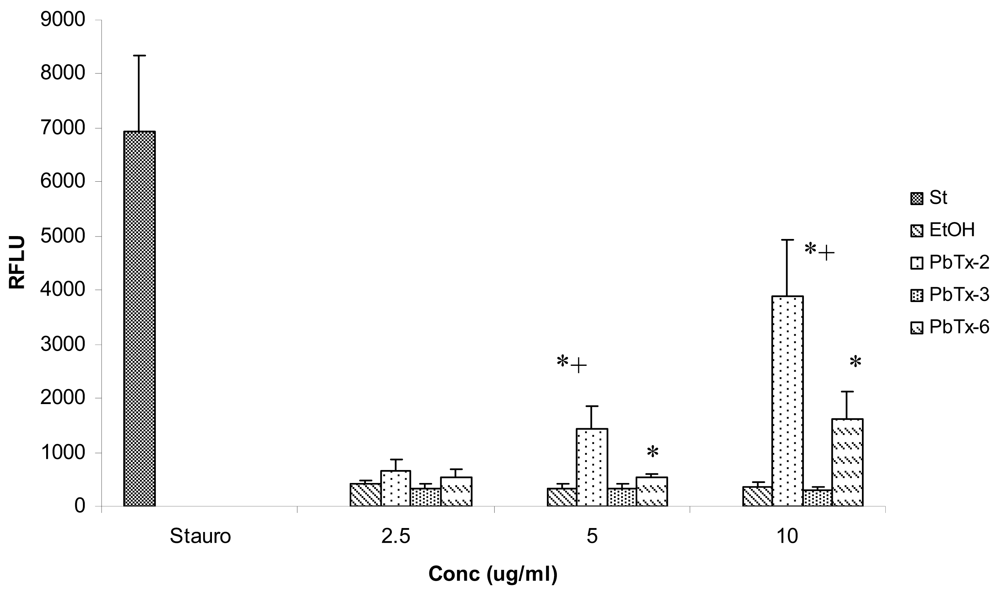
2.3 Effects of in vitro brevetoxin exposure on apoptosis in Jurkat cells
3. Experimental Section
3.1 Cells
3.2 Toxins
3.3 Cell Viability
3.4 MTT Assay
3.5 Caspase-3 activity
3.6 Statistical techniques
Acknowledgements
References
- Van Dolah, FM. Marine Algal Toxins: Origins, Health Effects, and Their Increased Occurrence. Environ Health Perspect 2000, 108, 133–141. [Google Scholar]
- Knap, A; Dwailly, E; Furgal, C; Galvin, J; Baden, D; Bowen, RE; Depledge, M; Duguay, L; Fleming, LE; Ford, T; Moser, F; Owen, R; Suk, WA; Unluata, U. Indicators of Ocean Health and Human Health: Developing a Research and Monitoring Framework. Environ Health Perspect 2002, 110, 839–845. [Google Scholar]
- Sandifer, PA; Holland, AF; Rowles, TK; Scott, GI. The Oceans and Human Health. Environ Health Perspect 2004, 112(8), A454–455. [Google Scholar]
- Tyson, FL; Rice, DL; Dearry, A. Connecting the Oceans and Human Health. Environ Health Perspect 2004, 112(8), A455–A456. [Google Scholar]
- Pierce, RH; Henry, MS; Blum, PC. Brevetoxin Abundance and Composition during ECOHAB-Florida Field Monitoring Cruises in the Gulf of Mexico. Continent Shelf Res 2008, 28, 45–58. [Google Scholar]
- Cheng, YS; Zhou, Y; Irvin, CM; Pierce, RH; Naar, J; Backer, LC; Fleming, LE; Kirkpatrick, B; Baden, DG. Characterization of Marine Aerosol for Assessment of Human Exposure to Brevetoxins. Environ Health Perspect 2005, 113(5), 638–643. [Google Scholar]
- Bourdelais, J; Jacobs, HM; Wright, JLC; Bigwarf, PM; Baden, DG. A New Polyether Ladder Compound Produced by the Dinoflagellate, Karenia brevis. J Nat Prod 2005, 68, 2–6. [Google Scholar]
- Abraham, A; Plakas, SM; Wang, Z; Jester, EL; El Said, KR; Granade, HR; Henry, MS; Blum, PC; Pierce, RH; Dickey, RW. Characterization of Polar Brevetoxin Derivatives Isolated from Karenia brevis Cultures and Natural Blooms. Toxicon 2006, 48(1), 104–115. [Google Scholar]
- Backer, LC; Fleming, LE; Rowan, A; Cheng, Y-S; Benson, J; Pierce, RH; Zaias, J; Bean, J; Bossart, GD; Johnson, D; Quimbo, R; Baden, DG. Recreational Exposure to Aerosolized Brevetoxins during Florida Red Tide Events. Harmful Algae 2003, 2, 19–29. [Google Scholar]
- Backer, LC; Kirkpatrick, B; Fleming, LE; Cheng, YS; Pierce, R; Bean, JA; Clark, R; Johnson, D; Wanner, A; Tamer, R; Zhou, Y; Baden, DG. Occupational Exposure to Aerosolized Brevetoxins during Florida Red Tide Events: Effects on a Healthy Worker Population. Environ Health Perspect 2005, 113(5), 644–649. [Google Scholar]
- Kirkpatrick, B; Fleming, LE; Squicciarini, D; Backer, LC; Clark, R; Abraham, W; Benson, J; Cheng, YS; Johnson, D; Pierce, R; Zaias, J; Bossart, GD; Baden, DG. Literature Review of Florida Red Tide: Implications for Human Health Effects. Harmful Algae 2004, 3(2), 99–115. [Google Scholar]
- Fleming, LE; Backer, LC; Baden, DG. Overview of Aerosolized Florida Red Tide Toxins: Exposures and Effects. Environ Health Perspect 2005, 113(5), 618–620. [Google Scholar]
- Fleming, LE; Kirkpatrick, B; Backer, LC; Bean, JA; Wanner, A; Dalpra, D; Tamer, R; Zaias, J; Cheng, YS; Pierce, R; Naar, J; Abraham, W; Clark, R; Zhou, Y; Henry, MS; Johnson, D; Van de Bogart, G; Bossart, GD; Harrington, M; Baden, DG. Initial Evaluation of the Effects of Aerosolized Florida Red Tide Toxins (Brevetoxins) in Persons with Asthma. Environ Health Perspect 2005, 113(5), 650–657. [Google Scholar]
- Fleming, LE; Kirkpatrick, B; Backer, LC; Bean, JA; Wanner, A; Reich, A; Zaias, J; Cheng, YS; Pierce, R; Naar, J; Abraham, WM; Baden, DG. Aerosolized Red-Tide Toxins (Brevetoxins) and Asthma. Chest 2007, 131(1), 187–194. [Google Scholar]
- Kirkpatrick, B; Fleming, LE; Backer, LC; Bean, JA; Tamer, R; Kirkpatrick, G; Kane, T; Wanner, A; Dalpra, D; Reich, A; Baden, DG. Environmental Exposures to Florida Red Tides: Effects on Emergency Room Respiratory Diagnoses Admissions. Harmful Algae 2006, 5, 526–33. [Google Scholar]
- Bossart, GD; Baden, DG; Ewing, R; Roberts, B; Wright, S. Brevetoxicosis in Manatees (Trichechus manatus latirostris) from the 1996 Epizootic: Gross, Histopathologic, and Immunocytochemical Features. Toxicol Pathol 1998, 26, 276–282. [Google Scholar]
- Walsh, CJ; Luer, CA; Noyes, DR. Effects of Environmental Stressors on Lymphocyte Proliferation in Florida Manatees, Trichechus manatus latirostris. Vet Immunol Immunopathol 2005, 103, 247–256. [Google Scholar]
- Kreuder, C; Mazet, JAK; Bossart, GD; Carpenter, TE; Holyoak, M; Elie, MS; Wright, SD. Clinicopathologic Features of Suspected Brevetoxicosis in Double-Crested Cormorants (Phalacrocorax auritus) Along the Florida Gulf Coast. J Zoo Wildlife Med 2002, 33, 8–15. [Google Scholar]
- Benson, JM; Tischler, DL; Baden, DG. Uptake, Tissue Distribution, and Excretion of Brevetoxin 3 Administered to Rats by Intratracheal Instillation. J Toxicol Environ Health A 1999, 57, 345–355. [Google Scholar]
- Benson, J; Hahn, F; March, T; McDonald, J; Sopori, M; Seagrave, J; Gomez, A; Bourdelais, A; Naar, J; Zaias, J; Bossart, G; Baden, D. Inhalation Toxicity of Brevetoxin 3 in Rats Exposed for 5 Days. J Toxicol Environ Health A 2004, 67(18), 1443–1456. [Google Scholar]
- Benson, JM; Hahn, FF; March, TH; McDonald, JD; Gomez, AP; Sopori, MJ; Bourdelais, AJ; Naar, J; Zaias, J; Bossart, GD; Baden, DG. Inhalation Toxicity of Brevetoxin 3 in Rats Exposed for Twenty-two Days. Environ Health Perspect 2005, 113, 626–631. [Google Scholar]
- Sayer, A; Hu, Q; Bourdelais, AJ; Baden, DG; Gibson, JE. The Effect of Brevetoxin-Induced DNA Damage in Human Lymphocytes. Genotoxicology 2005, 79, 683–688. [Google Scholar]
- Han, TK; Derby, M; Martin, DF; Wright, SD; Dao, ML. Effects of Brevetoxins on Murine Myeloma SP2/O Cells. Aberrant Cellular Division. Int J Toxicol 2002, 22, 73–80. [Google Scholar]
- Sayer, A; Hu, Q; Bourdelais, AJ; Baden, DG; Gibson, JE. The Inhibition of CHO-K1-BH4 Cell Proliferation and Induction of Chromosomal Aberrations by Brevetoxins in vitro. Food Chem Toxicol 2006, 44, 1082–1092. [Google Scholar]
- Abraham, RT; Weiss, A. Jurkat T cells and development of the T-cell receptor signaling paradigm. Nat Rev Immunol 2004, 4, 301–308. [Google Scholar]
- Desai, R; Peretz, A; Idelson, H; Lazarovici, P; Attali, B. Ca2+ -activated K+ channels in human leukemic Jurkat T cells. J Biol Chem 2000, 275, 39954–39963. [Google Scholar]
- Morimoto, T; Ohya, S; Hayashi, H; Onozaki, K; Imaizumi, Y. Cell-cycle-dependent regulation of Ca2+ -activated K+ channel in Jurkat T-Lymphocyte. J Pharmacol Sci 2007, 104, 94–98. [Google Scholar]
- Eisenbrand, G; Pool-Zobel, B; Baker, V; Balls, M; Blaauboer, BJ; Boobis, A; Carere, A; Kevekordes, S; Pieters, R; Kleiner, J. Methods of in vitro toxicology. Food Chem Toxicol 2002, 40, 193–206. [Google Scholar]
- Radwan, FF; Wang, Z; Ramsdell, JS. Identification of a Rapid Detoxification Mechanism for Brevetoxin in Rats. Toxicol Sci 2005, 85(2), 839–846. [Google Scholar]
- Woofter, R; Dechraoui, MY; Garthwaite, I; Towers, NR; Gordon, CJ; Córdova, J; Ramsdell, JS. Measurement of Brevetoxin Levels by Radioimmunoassay of Blood Collection Cards after Acute, Long-term, and Low Dose Exposure in Mice. Environ Health Perspect 2003, 111, 1595–1600. [Google Scholar]
- Woofter, RT; Brendtro, K; Ramsdell, JS. Uptake and Elimination of Brevetoxin in Blood of Striped Mullet (Mugil cephalus) after Aqueous Exposure to Karenia brevis. Environ Health Perspect 2005, 11(1), 11–16. [Google Scholar]
- Tibbetts, BM; Baden, DG; Benson, JM. Uptake, Tissue Distribution, and Excretion of Brevetoxin-3 Administered to Mice by Intratracheal Instillation. J Toxicol Environ Health A 2006, 69(14), 1325–1335. [Google Scholar]
- Woofter, RT; Spiess, PC; Ramsdell, JS. Distribution of Brevetoxin (PbTx-3) in Mouse Plasma: Association with High-Density Lipoproteins. Environ Health Perspect 2005, 113(11), 1491–1496. [Google Scholar]
- Woofter, RT; Ramsdell, JS. Distribution of Brevetoxin to Lipoproteins in Human Plasma. Toxicon 2007, 49, 1010–1018. [Google Scholar]
- Benson, JM; Gomez, AP; Statom, GL; Tibbetts, BM; Fleming, LE; Backer, LC; Reich, A; Baden, DG. Placental Transport of Brevetoxin-3 in CD-1 Mice. Toxicon 2006, 48, 1018–1026. [Google Scholar]
- Katunuma, N; Matsunaga, Y; Himeno, K; Hayashi, Y. Insights into the Roles of Cathepsin in Antigen Processing and Presentation Revealed by Specific Inhibitors. Biol Chem 2003, 384, 883–890. [Google Scholar]
- Sudarsanam, S; Virca, GD; March, CJ; Srinivasan, S. An Approach to Computer-Aided Inhibitory Design: Application to Cathepsin L. J Comput Aided Mol Des 1992, 6(3), 223–233. [Google Scholar]
- Goyarts, T; Dänicke, S; Tiemann, U; Rothkötter, HJ. Effect of the Fusarium Toxin Deoxynivalenol (DON) on IgA, IgM, and IgG Concentrations and Proliferation of Porcine Blood Lymphocytes. Toxicol In vitro 2006, 20(6), 858–867. [Google Scholar]
- Minervini, F; Giannoccaro, A; Cavallini, A; Visconti, A. Investigations on Cellular Proliferation Induced by Zearalenone and Its Derivatives in Relation to the Estrogenic Parameters. Toxicol Lett 2005, 159(3), 272–283. [Google Scholar]
- Khan, KN; Masuzaki, H; Fujishita, A; Kitajima, M; Kohno, T; Sekine, I; Matsuyama, T; Ishimaru, T. Regulation of Hepatocyte Growth Factor by Basal and Stimulated Macrophages in Women with Endometriosis. Human Reprod 2005, 20, 2004–2013. [Google Scholar]
- Abraham, WM; Bourdelais, AJ; Sabater, JR; Ahmed, A; Lee, TA; Serebriakov, I; Baden, DG. Airway Responses to Aerosolized Brevetoxin in an Animal Model of Asthma. Am J Respir Crit Care Med 2005, 171, 26–34. [Google Scholar]
- Jeglitsch, G; Rein, K; Baden, DG; Adams, DJ. Brevetoxin-3 (PbTx-3) and its Derivatives Modulate Single Tetrodotoxin-Sensitive Sodium Channels in Rat Sensory Neurons. J Pharmacol Exptl Therap 1998, 284(2), 516–525. [Google Scholar]
- Purkerson, SL; Baden, DG; Fieber, LA. Brevetoxin Modulates Neuronal Sodium Channels in Two Cell Lines Derived from Rat Brain. Neurotoxicology 1999, 20(6), 909–920. [Google Scholar]
- Baden, DG; Bourdelais, AJ; Jacocks, H; Michelliza, S; Naar, J. Natural and Derivative Brevetoxins: Historical Background, Multiplicity, and Effects. Environ Health Perspect 2005, 113(5), 621–625. [Google Scholar]
- Washburn, BS; Rein, KS; Baden, DG; Walsh, PJ; Hinton, DE; Tullis, K; Denison, MS. Brevetoxin-6 (PbTx-6), a Nonaromatic Marine Neurotoxin, is a Ligand of the Aryl Hydrocarbon Receptor. Arch Biochem Biophys 1997, 343(2), 149–156. [Google Scholar]
- Radwan, FF; Ramsdell, JS. Characterization of In vitro Oxidative and Conjugative Metabolic Pathways for Brevetoxin (PbTx-2). Toxicol Sci 2006, 89(1), 57–65. [Google Scholar]
- Washburn, BS; Baden, DG; Gassman, NJ; Walsh, PJ. Brevetoxin: Tissue Distribution and Effect on Cytochrome P450 Enzymes in Fish. Toxicon 1994, 32(7), 799–805. [Google Scholar]
- Dey, A; Dhawan, A; Kishore Seth, P; Parma, D. Evidence for Cytochrome P450 2E1 Catalytic Activity and Expression in Rat Blood Lymphocytes. Life Sci 2005, 77, 1082–1093. [Google Scholar]
- Haas, CE; Brazeau, D; Cloen, D; Booker, BM; Frerichs, V; Zaranek, C; Frye, RF; Kufel, T. Cytochrome P450 mRNA Expression in Peripheral Blood Lymphocytes as a Predictor of Enzyme Induction. Eur J Clin Parmacol 2005, 61, 583–593. [Google Scholar]
- Van Duursen, MBM; Sanderson, JT; Van den Berg, M. Cytochrome P450 1A1 and 1B1 in Human Blood Lymphocytes are Not Suitable as Biomarkers of Exposure to Dioxin-Like Compounds: Polymorphisms and Interindividual Variation in Expression and Inducibility. Toxicol Sci 2005, 85, 703–712. [Google Scholar]
- Pierce, RH; Henry, MS; Blum, PC; Hamel, SL; Kirkpatrick, B; Cheng, YS; Zhou, Y; Irvin, CM; Naar, J; Weidner, A; Fleming, LE; Backer, LC; Baden, DG. Brevetoxin Composition in Water and Marine Aerosol Along a Florida Beach: Assessing Potential Human Exposure to Marine Biotoxins. Harmful Algae 2005, 4, 965–972. [Google Scholar]
- Baur, H; Kaspare, S; Pfaff, E. Criteria of Viability of Isolated Liver Cells. Hoppe-Seylers Z Physiol Chem 1975, 356, 827–838. [Google Scholar]
- Mosmann, TJ. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J Immunol Meth 1983, 65, 55–63. [Google Scholar]
- Sugawara, J; Fukaya, T; Murakami, T; Yoshida, H; Yajima, A. Hepatocyte Growth Factor Stimulates Proliferation, Migration, and Lumen Formation of Human Endometrial Epithelial Cells in vitro. Biol Reprod 1997, 57, 936–942. [Google Scholar]
- Nicholson, DW; Ali, A; Thornberry, NA; Vaillancourt, JP; Ding, CK; Gallant, M; Gareau, Y; Griffin, PR; Labelle, M; Lazebnik, YA; et al. Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature 1995, 376(6535), 37–43. [Google Scholar]





Share and Cite
Walsh, C.J.; Leggett, S.R.; Strohbehn, K.; Pierce, R.H.; Sleasman, J.W. Effects of in vitro Brevetoxin Exposure on Apoptosis and Cellular Metabolism in a Leukemic T Cell Line (Jurkat). Mar. Drugs 2008, 6, 291-307. https://doi.org/10.3390/md6020291
Walsh CJ, Leggett SR, Strohbehn K, Pierce RH, Sleasman JW. Effects of in vitro Brevetoxin Exposure on Apoptosis and Cellular Metabolism in a Leukemic T Cell Line (Jurkat). Marine Drugs. 2008; 6(2):291-307. https://doi.org/10.3390/md6020291
Chicago/Turabian StyleWalsh, Catherine J., Stephanie R. Leggett, Kathryn Strohbehn, Richard H. Pierce, and John W. Sleasman. 2008. "Effects of in vitro Brevetoxin Exposure on Apoptosis and Cellular Metabolism in a Leukemic T Cell Line (Jurkat)" Marine Drugs 6, no. 2: 291-307. https://doi.org/10.3390/md6020291
APA StyleWalsh, C. J., Leggett, S. R., Strohbehn, K., Pierce, R. H., & Sleasman, J. W. (2008). Effects of in vitro Brevetoxin Exposure on Apoptosis and Cellular Metabolism in a Leukemic T Cell Line (Jurkat). Marine Drugs, 6(2), 291-307. https://doi.org/10.3390/md6020291