Distribution and Abundance of MAAs in 33 Species of Microalgae across 13 Classes

Abstract
:1. Introduction
2. Results and Discussion
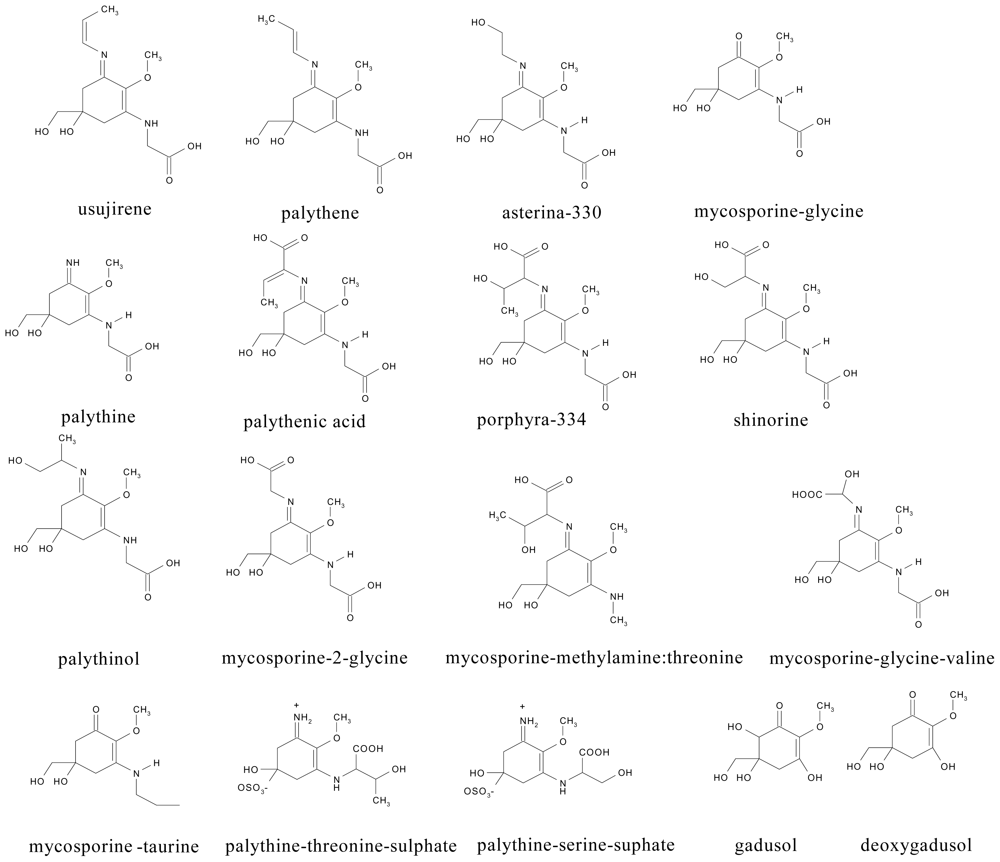
2.1. Chromatographic separation and detection of MAAs
2.2. Distribution and abundance of MAAs in cultures
2.3. Characteristics of MAAs in microalgal cultures
2.4. Factors influencing induction and distribution of MAAs
2.5. Comparison with natural samples
2.6. Spectrophotometric analysis of cultures
3. Experimental
3.1. Culturing and sampling
3.2. Extraction and HPLC analysis
3.3. MAA identification and quantification
4. Conclusions
Acknowledgements
- Samples Availability: Not available.
References
- Dunlap, WC; Yamamoto, Y. Small-molecule antioxidants in marine organisms: Antioxidant activity of mycosporine-glycine. Comp Biochem Physiol B 1995, 112, 105–114. [Google Scholar]
- Oren, A; Gunde-Cimerman, N. Mycosporines and mycosproine-like amino acids: UV protectants or multipurpose secondary metabolites. FEMS Microbiol Lett 2007, 269, 1–10. [Google Scholar]
- Favre-Bonvin, J; Arpin, N; Brevard, C. Structure de la mycosporine (p310). Can J Chem 1976, 54, 1105–1113. [Google Scholar]
- Karentz, D. McClintock, JB, Baker, BJ, Eds.; Chemical defences of marine organisms against solar radiation exposure: UV-absorbing mycosporine-like amino acids and scytonemin. In Marine Chemical Ecology; CRC Press: Boca Raton, Florida, 2001; pp. 481–520. [Google Scholar]
- Shick, JM; Dunlap, WC. Mycosporine-like amino acids and related gadusols: Biosynthesis, accumulation and UV-protective functions in aquatic organisms. Ann Rev Physiol 2002, 64, 223–262. [Google Scholar]
- Dunlap, WC; Chalker, BE. Identification and quantification of near UV absorbing compounds (s320) in a hermatypic scleractinian. Coral Reefs 1986, 5, 155–159. [Google Scholar]
- Marchant, HJ; Davidson, AT; Kelly, GJ. UV-B protecting compounds in the marine alga Phaeocystis pouchetii from Antarctica. Mar Biol 1991, 109, 391–395. [Google Scholar]
- Karentz, D; McCuen, FS; Land, MC; Dunlap, WC. Survey of mycosporine-like amino acid compounds in Antarctic marine organisms: Potential protection from ultraviolet exposure. Mar Biol 1991, 108, 157–166. [Google Scholar]
- Favre-Bonvin, J; Bernillon, J; Salin, N; Arpin, N. Biosynthesis of mycosporines: Mycosporine glutaminol in Trichothecium roseum. Phytochemistry 1987, 29, 2509–2514. [Google Scholar]
- Shick, JM; Romaine-Lioud, S; Ferrier-Pages, C; Gattuso, J-P. Ultraviolet-B radiation stimulates shikimate pathway-dependent accumulation of mycosporine-like amino acids in the coral Stylophora pistillata despite decreases in its population of symbiotic dinoflagellates. Limnol Oceanogr 1999, 44, 1667–1682. [Google Scholar]
- Portwich, A; Garcia-Pichel, F. Biosynthetic pathway of mycosporines (mycosporine-like amino acids) in the cyanobacterium Chlorogloeopsis sp strain PCC 6912. Phycologia 2003, 42, 384–392. [Google Scholar]
- Cockell, CS; Knowland, J. Ultraviolet radiation screening compounds. Biol Rev 1999, 74, 311–345. [Google Scholar]
- Waller, RF; Slamovits, CH; Keeling, PJ. Lateral gene transfer of a multigene region from cyanobacteria to dinoflagellates resulting in a novel plastid-targeted fusion protein. Mol Biol Evol 2006. [Google Scholar]
- Starcevic, A; Akthar, S; Dunlap, WC; Shick, JM; Hranueli, D; Cullum, J; Long, PF. Enzymes of the shikimic acid pathway encoded in the genome of a basal metazoan, Nematostella vectensis have microbial origins. PNAS 2008, 105, 2533–2537. [Google Scholar]
- Bandaranayake, WM. Mycosporines: Are they nature’s sunscreens. Nat Prod Rep 1998, 159–172. [Google Scholar]
- Wu Won, JJ; Chalker, B; Rideout, JA. Two new UV-absorbing compounds from Stylophora pistallata: Sulfate esters of mycosporine-like amino acids. Tetrahedron Lett 1997, 38, 2525–2526. [Google Scholar]
- Böhm, GA; Pfleiderer, W; Böger, P; Scherer, S. Structure of a novel oligosaccharide-mycosporine-amino acid ultraviolet A/B sunscreen pigment from the terrestrial cyanobacterium Nostoc commune. J Biol Chem 1995, 270, 9–17. [Google Scholar]
- Carreto, JI; Carignan, MO; Montoya, NG. Comparative studies on mycosporine-like amino acids, paralytic shellfish toxins and pigment profiles of the toxic dinoflagellates Alexandrium tamarense, A. cantenella and A. minutum. Mar Ecol Prog Ser 2001, 223, 49–60. [Google Scholar]
- Sommaruga, R; Garcia-Pichel, F. UV-absorbing compounds in planktonic and benthic organisms from a high-mountain lake. Arch Hydrobiol 1999, 144, 255–269. [Google Scholar]
- Karsten, U; Wiencke, C. Factors controlling the formation of UV-absorbing mycosporine-like amino acids in the marine red alga Palmaria palmata from Spitsbergen (Norway). J Plant Physiol 1999, 155, 407–415. [Google Scholar]
- Xiong, FS; Kopecky, J; Nedbal, L. The occurrence of UV-B absorbing mycosporine-like amino acids in freshwater and terrestrial microalgae (Chlorophyta). Aquat Bot 1999, 63, 37–49. [Google Scholar]
- Jeffrey, SW; Mactavish, HS; Dunlap, WC; Vesk, M; Groenewoud, K. Occurrence of UVA and UVB-absorbing compounds in 152 species (306 strains) of marine microalgae. Mar Ecol Prog Ser 1999, 189, 35–51. [Google Scholar]
- Whitehead, K; Vernet, M. Influence of mycosporine-like amino acids (MAAs) on absorption by particulate and dissolved organic matter in La Jolla Bay. Limnol Oceanog 2000, 45, 1788–1796. [Google Scholar]
- Liu, Z; Häder, DP; Sommaruga, R. Occurrence of mycosporine-like amino acids (MAAs) in the bloom-forming cyanobacterium Microcystis aeruginosa. J Plankton Res 2004, 26, 963–966. [Google Scholar]
- Gröniger, A; Sinha, RP; Klish, M; Häder, DP. Photoprotective compounds in cyanobacteria, phytoplankton and macroalgae—a database. J Photochem Photobiol B Biol 2000, 58, 115–122. [Google Scholar]
- Sinha, RP; Singh, SP; Häder, DP. Database on mycosporine and mycosporine-like amino acids (MAAs) in fungi, cyanobacteria, macroalgae, phytoplankton and animals. J Photochem Photobiol B 2007, 89, 29–35. [Google Scholar]
- Schid, D; Schürch, C; Zülli, F; Nissen, H-P; Prieur, H. Mycosporine-like amino acids: Natural UV sunscreening compounds from red algae to protect the skin against photoaging. SÖFW J 2003, 129, 1–5. [Google Scholar]
- Carreto, JI; Carignan, MO; Montoya, NG. A high-resolution reverse-phase liquid chromatography method for the analysis of mycosporine-like amino acids (MAAs) in marine organisms. Mar Biol 2005, 146, 237–252. [Google Scholar]
- Hannach, G; Sigleo, AC. Photoinduction of UV-absorbing compounds in six species of marine phytoplankton. Mar Ecol Prog Ser 1998, 174, 207–222. [Google Scholar]
- Wängberg, S-A; Persson, A; Karlson, B. Effects of UV-B radiation on synthesis of mycosporine-like amino acid and growth in Heterocapsa triquetra (Dinophyceae). J Photochem Photobiol B 1997, 37, 141–146. [Google Scholar]
- Laurion, I; Roy, S. Growth and photoprotection in three dinoflagellates (including two strains of Alexandrium tamarense) and one diatom exposed to four weeks of natural and enhanced ultraviolet–B radiation. J Phycol 2009, 45, 16–33. [Google Scholar]
- Taira, H; Aoki, S; Yamanoha, B; Taguchi, S. Daily variation in cellular content of UV-absorbing compounds mycosporine-like amino acids in the marine dinoflagellate Scrippsiella sweeneyae. J Photochem Photobiol B 2004, 75, 145–155. [Google Scholar]
- Laurion, I; Blouin, F; Roy, S. Packaging of mycosporine-like amino acids in dinoflagellates. Mar Ecol Prog Ser 2004, 279, 297–303. [Google Scholar]
- Shick, JM; Dunlap, WC; Pearse, JS; Pearse, VB. Mycosporine-like amino acid content in four species of sea anemones in the Genus Anthopleura reflects phylogenetic but not environmental or symbiotic relationships. Biol Bull 2002, 203, 315–330. [Google Scholar]
- Singh, SP; Klisch, M; Sinha, RP; Häder, DP. Genome mining of mycosporine-like amino acid (MAA) synthesizing and non-synthesizing cyanobacteria: A bioinformatics study. Genomics 2010, 95, 120–128. [Google Scholar]
- Riegger, L; Robinson, D. Photoinduction of UV-absorbing compounds in Antarctic diatoms and Phaeocystis antarctica. Mar Ecol Prog Ser 1997, 160, 13–25. [Google Scholar]
- Moisan, TA; Mitchell, BG. UV absorption by mycosporine-like amino acids in Phaeocystis antarctica Karsten induced by photosynthetically available radiation. Mar Biol 2001, 138, 217–227. [Google Scholar]
- Llewellyn, CA; Harbour, D. A temporal study of UV-sunscreen compounds in surface water phytoplankton from the English Channel; correlation with natural solar UV radiation. J Mar Biol Ass UK 2003, 83, 1–9. [Google Scholar]
- Buma, AGJ; Wright, SW; van den Enden, R; van de Poll, WH; Davidson, HT. PAR acclimation and UVBR-induced DNA damage in Antarctic marine microalgae. Mar Ecol Prog Ser 2006, 315, 33–42. [Google Scholar]
- Helbling, EW; Chalker, BE; Dunlap, WC; Holm-Hansen, O; Villafañe, VE. Photoacclimation of Antarctic marine diatoms to solar ultraviolet radiation. J Exp Mar Biol Ecol 1996, 204, 85–101. [Google Scholar]
- Karsten, U; Friedl, T; Schumann, R; Hoyer, K; Lembcke, S. Mycosporine-like amino acids and phylogenies in green algae: Prasiola and its relatives from the Trebouxiophyceae (Chlorophyta). J Phycol 2005, 41, 557–566. [Google Scholar]
- Llewellyn, CA; Mantoura, RFC. A UV absorbing compound in HPLC pigment chromatograms from Icelandic Basin phytoplankton. Mar Ecol Prog Ser 1997, 158, 283–287. [Google Scholar]
- Neale, PJ; Banaszak, AT; Jarriel, CR. Ultraviolet sunscreens in Gymnodinium sanguineum (Dinophyceae): Mycosprine-like amino acids protect against inhibition of photosynthesis. J Phycol 1998, 34, 928–938. [Google Scholar]
- Franklin, LA; Krabs, G; Kuhlenkamp, R. Blue light and UV-A radiation control the synthesis of mycosporine-like amino acids in Chondrus crispus (Florideophyceae). J Phycol 2001, 37, 257–270. [Google Scholar]
- Kräbs, G; Bischof, K; Hanelt, D; Karsten, U; Wiencke, C. Wavelength-dependent induction of UV-absorbing mycosporine-like amino acids in the red algae Chondrus crispus under natural solar radiation. J Exp Mar Biol Ecol 2002, 268, 69–82. [Google Scholar]
- Hoyer, K; Karsten, U; Wiencke, C. Induction of sunscreen compounds in Antarctic macroalgae by different radiation conditions. Mar Biol 2002, 141, 619–627. [Google Scholar]
- Banaszak, AT; LaJeunesse, TC; Trench, RK. The synthesis of mycosporine-like amino acids (MAAs) by cultured, symbiotic dinoflagellates. J Exp Mar Biol Ecol 2000, 249, 219–233. [Google Scholar]
- Peinado, NK; Diaz, RTA; Figueroa, FL; Helbling, EW. Ammonium and UV radiation stimulate the accumulation of mycosporine-like amino acids in Porphyra columbina (Rhodophyta) from Patagonia, Argentina. J Phycol 2004, 40, 248–259. [Google Scholar]
- Litchman, E; Neale, PJ; Banaszak, AT. Increased sensitivity to ultraviolet radiation in nitrogen- limited dinoflagellates: Photoprotection and repair. Limnol Oceanogr 2002, 47, 86–94. [Google Scholar]
- White, Daniel. Plymouth Marine Laboratory: Plymouth, UK, Personal communication; 2009.
- Suzuki, R; Ishimaru, T. Characteristics of photosynthetic pigment composition of Gymnodinium mikimotoi. J Oceanog 1992, 48, 367–375. [Google Scholar]
- Whitehead, K; Hedges, JI. Electrospray ionisation tandem mass spectrometric and electron impact mass spectrometric characterization of mycosporine-like amino acids. Rapid Commun Mass Spectrom 2003, 17, 2133–2138. [Google Scholar]



{kind=link}
{kind=link}
| (a) | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Class | Bacillario phyceae | Chlorara chniophyceae | Chloro phyceae | Chryso phyceae | Crytophyceae | Cyano phyceae | Dinophyceae | |||||||||||||
| Species | P. tricornatum | T. weissflogii | C. reptans | C. concordia | D. tertiolecta | P. subviridis | R. maritime | C. maculata | C. salina | H brunnescens | R. reticulata | N. commune | A. tamarense | A carterae | G. foliaceum | G.galatheanum | G. venificum | H. triquetra | ||
| Peak No. | λmax (nm) | MAA | ||||||||||||||||||
| 1 | 330,346,366 | |||||||||||||||||||
| 2 | 292,306,320 | 2 | ||||||||||||||||||
| 3 | 278 | x | x | x | x | x | ||||||||||||||
| 4 | 354 | Usijerene | ||||||||||||||||||
| 5 | 360 | Palythene | 12 | 109 | ||||||||||||||||
| 6 | 314 | |||||||||||||||||||
| 7 | 280 | x | x | x | x | x | ||||||||||||||
| 8 | 330 | Asterina | ||||||||||||||||||
| 9 | 310 | M-glycine | 2 | 125 | 310 | |||||||||||||||
| 10 | 320 | Palythine | 5 | 111 | 503 | |||||||||||||||
| 11 | 276 | x | x | x | x | x | x | |||||||||||||
| 12 | 342 | 35 | 7 | |||||||||||||||||
| 13 | 294 | x | x | x | x | x | x | x | x | x | x | |||||||||
| 14 | 310 | |||||||||||||||||||
| 15 | 338 | Palyth. acid | 0 | 373 | 1273 | 553 | ||||||||||||||
| 16 | 334 | Porphyra-334 | x | 41 | 72 | 293 | ||||||||||||||
| 17 | 336 | x | ||||||||||||||||||
| 18 | 334 | Shinorine | 12 | 1 | 6497 | 32 | 10 | |||||||||||||
| 19 | 352 | |||||||||||||||||||
| 20 | 320 | 2 | ||||||||||||||||||
| 21 | 334 | x | ||||||||||||||||||
| 22 | 336 | 89 | 15 | |||||||||||||||||
| 23 | 328 | |||||||||||||||||||
| 24 | 384 | |||||||||||||||||||
| 25 | 334/336 | 6 | 44 | 30 | ||||||||||||||||
| 26 | 334 | |||||||||||||||||||
| 27 | 334 | |||||||||||||||||||
| 28 | 308 | x | ||||||||||||||||||
| 29 | 374 | x | ||||||||||||||||||
| fg cell−1 | 12 | 10 | 61 | 6913 | 1737 | 1900 | ||||||||||||||
| (b) | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Class Species | Dino phyceae cont’d | Eugleno phyceae | Eustigmato phyceae | Prasino phyceae | Prymnesio phyceae | Raphido phyceae | Rhodo phyceae | ||||||||||||
| O. marina | P lima | P. micans | P. minimum | S. trochoidea | E. gracilis | N. oculata | N. rotunda | E. huxleyi | I. galbana | P. globosa | H. akashiwo | P. purpureum | R. marina | R. baltica | Plankton 7/07/2003 | Plankton 11/08/2003 | |||
| Peak No. | λmax (nm) | MAA | |||||||||||||||||
| 1 | 330,346, 366 | 54 | |||||||||||||||||
| 2 | 292,306, 320 | ||||||||||||||||||
| 3 | 278 | x | x | x | |||||||||||||||
| 4 | 354 | Usijerene | |||||||||||||||||
| 5 | 360 | Palythene | 95 | 67 | 40 | 296 | |||||||||||||
| 6 | 314 | x | x | ||||||||||||||||
| 7 | 280 | x | x | x | x | x | x | x | 6 | ||||||||||
| 8 | 330 | Asterina | 7 | ||||||||||||||||
| 9 | 310 | M-glycine | 37 | 17 | 149 | 35 | 176 | 47 | 24 | ||||||||||
| 10 | 320 | Palythine | 1 | 13 | 15 | 11 | |||||||||||||
| 11 | 276 | x | x | x | x | x | x | ||||||||||||
| 12 | 342 | 32 | 4 | ||||||||||||||||
| 13 | 294 | x | x | x | x | x | x | ||||||||||||
| 14 | 310 | 28 | 140 | ||||||||||||||||
| 15 | 338 | Palyth. acid | x | x | 12 | 18 | 28 | 6 | |||||||||||
| 16 | 334 | Porphyra-334 | 314 | 4 | 5324 | 7 | 15 | ||||||||||||
| 17 | 336 | x | 1 | ||||||||||||||||
| 18 | 334 | Shinorine | 296 | 8 | 66 | 31 | 277 | 41 | 7 | ||||||||||
| 19 | 352 | 1 | |||||||||||||||||
| 20 | 320 | x | x | 4 | 21 | ||||||||||||||
| 21 | 334 | x | |||||||||||||||||
| 22 | 336 | ||||||||||||||||||
| 23 | 328 | 1 | |||||||||||||||||
| 24 | 384 | x | 1 | ||||||||||||||||
| 25 | 334/336 | x | 5 | 28 | |||||||||||||||
| 26 | 334 | x | x | ||||||||||||||||
| 27 | 334 | x | 2 | ||||||||||||||||
| 28 | 308 | x | |||||||||||||||||
| 29 | 374 | x | x | ||||||||||||||||
| fg cell−1 | 387 | 121 | 597 | 146 | 6155 | 12 | 7 | 140 | 173 | 59 | |||||||||
| Culture | Group | a Culture code | Culture medium | Cells harvested (number * 106) |
|---|---|---|---|---|
| Thalassiosira weissflogii | Bacillariophyceae | INT | f/2-Si | 23 |
| Phaeodactylum tricornutum | “ | PLY 100 | f/2-Si | 9000 |
| Chlorarachnion reptans | Chlorarachniophyceae | CCAP 815/1 | f/2 | 10 |
| Chlamydomonas concordia | Chlorophyceae | PLY 491 | f/2 | 45 |
| Dunaliella tertiolecta | “ | PLY 83 | f/2 | not quantified |
| Ruttnera maritima | Chrysophyceae | PLY 177 | QA | 120 |
| Pelagococcus subviridis | “ | PLY 542 | f/2 | 590 |
| Cryptomonas maculata | Cryptophyceae | PLY 175 | f/2 | 95 |
| Chroomonas salina | “ | PLY 544 | f/2 | 210 |
| Rhinomonas reticulata | “ | PLY 358 | Q/A | 4.6 |
| Hemiselmis brunnescens | “ | PLY 14 | Erdschreiber | 17 |
| bNostoc commune | Cyanophyta | CCAP 1453/24 | BG11 | 240 |
| Amphidinium carterae | Dinophyceae | PLY 127 | f/2 | 3.4 |
| Prorocentrum lima | “ | PLY 558A | f/2 | 99 |
| Prorocentrum micans | “ | INT | f/2 | 29 |
| Alexandrium tamarense | “ | PLY 173A | f/2 | 140 |
| cOxyrrhis marina | “ | INT | f/2 | 4.6 |
| Prorocentrum minimum | “ | PLY 18B | f/2 | 32 |
| Scrippsiella trochoidea | “ | PLY 104 | Erdschreiber | 5.4 |
| Glenodinium foliaceum | “ | PLY 499 | Erdschreiber | 1.5 |
| Gymnodinium galatheanum | “ | PLY 517 | Erdschreiber | 0.62 |
| Heterocapsa triquetra | “ | PLY 169 | Erdschreiber | 0.05 |
| Gymnodinium venificum | “ | PLY 103 | Erdschreiber | 8 |
| dEuglena gracilis | Euglenophyceae | INT | QA | 150 |
| Nannochloropsis oculata | Eustigmatophyceae | CCAP 849/1 | f/2 | 125000 |
| Nephroselmis rotunda | Prasinophyceae | PLY 210 | QA | 150 |
| Phaeocystis globosa | Prymnesiophyceae | PLY 64 | f/2 | not quantified |
| Emiliania huxleyi | “ | PLY 92 | f/2 | 580 |
| Isochrysis galbana | “ | INT | f/2 | 1300 |
| Heterosigma akashiwo | Raphidophyceae | PLY 461 | f/2 | 380 |
| Porphyridinium purpureum | Rhodophyceae | PLY 539 | f/2 | 16 |
| Rhodomonas marina | “ | INT | f/2 | 66 |
| Rhodomonas baltica | “ | INT | f/2 | 7.6 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Llewellyn, C.A.; Airs, R.L. Distribution and Abundance of MAAs in 33 Species of Microalgae across 13 Classes. Mar. Drugs 2010, 8, 1273-1291. https://doi.org/10.3390/md8041273
Llewellyn CA, Airs RL. Distribution and Abundance of MAAs in 33 Species of Microalgae across 13 Classes. Marine Drugs. 2010; 8(4):1273-1291. https://doi.org/10.3390/md8041273
Chicago/Turabian StyleLlewellyn, Carole Anne, and Ruth Louise Airs. 2010. "Distribution and Abundance of MAAs in 33 Species of Microalgae across 13 Classes" Marine Drugs 8, no. 4: 1273-1291. https://doi.org/10.3390/md8041273
APA StyleLlewellyn, C. A., & Airs, R. L. (2010). Distribution and Abundance of MAAs in 33 Species of Microalgae across 13 Classes. Marine Drugs, 8(4), 1273-1291. https://doi.org/10.3390/md8041273