
Quorum Sensing Regulation of Competence and Bacteriocins in Streptococcus pneumoniae and mutans


Abstract
:1. Introduction
2. A Brief Introduction to Quorum Sensing
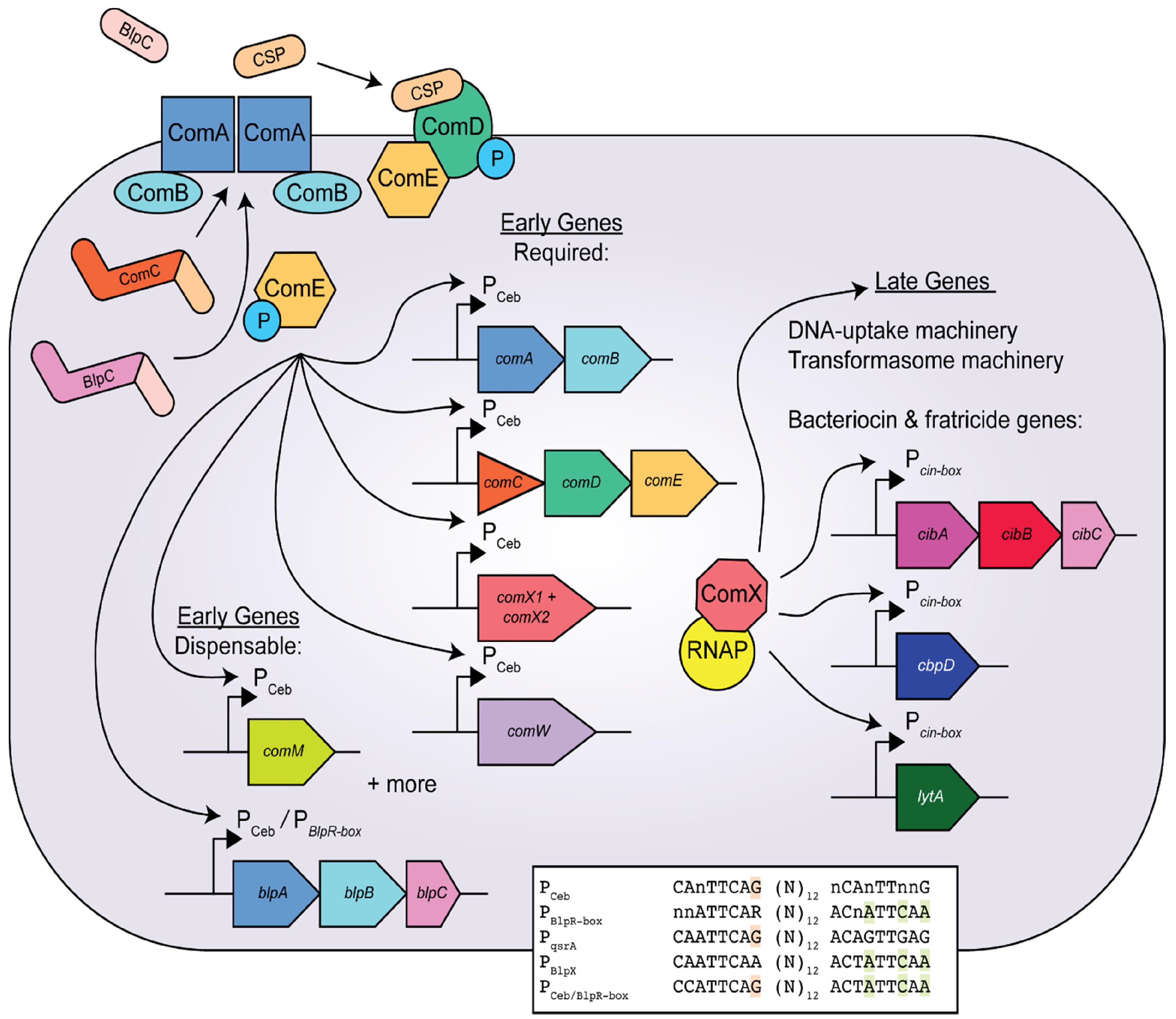
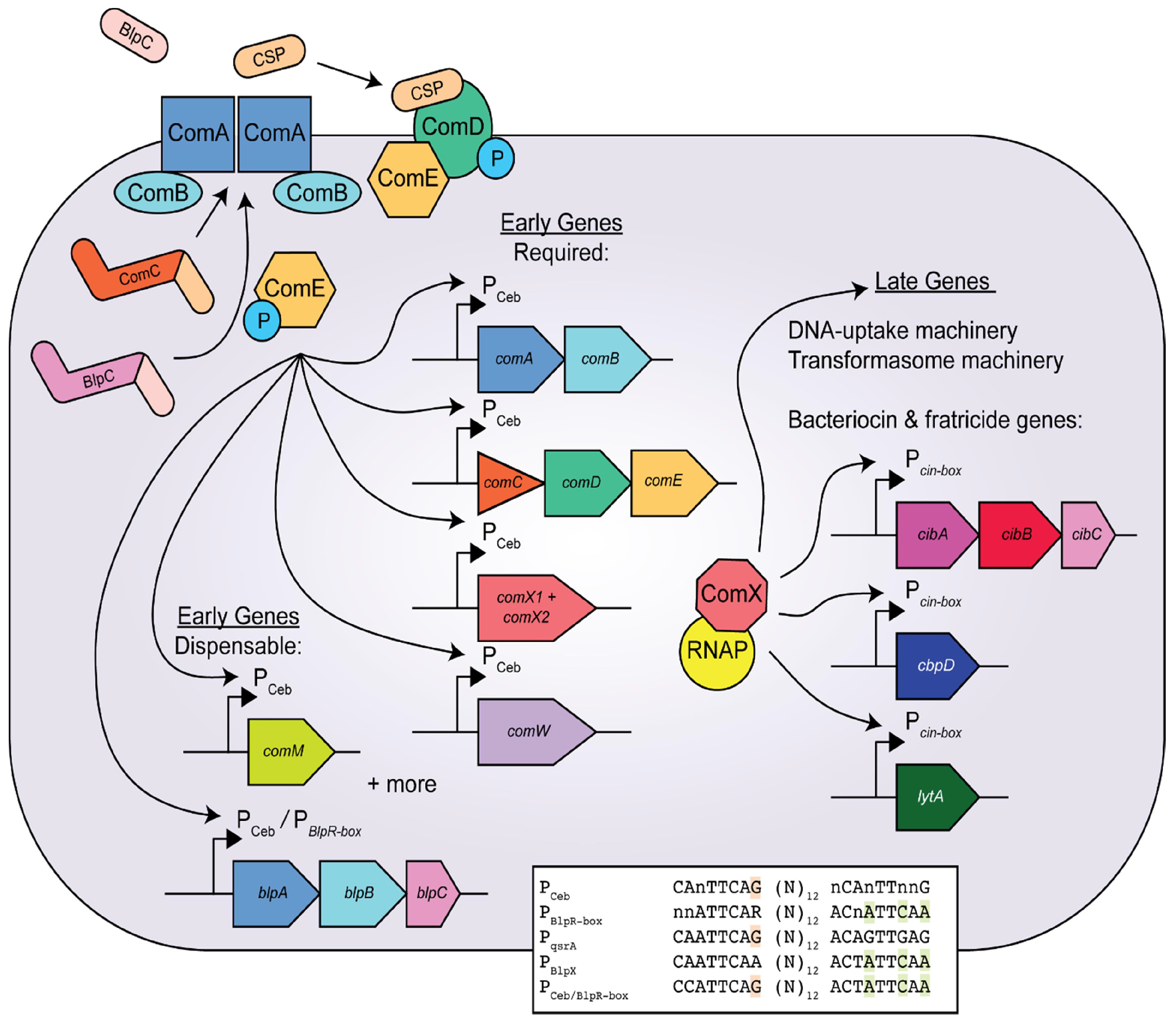
3. The Competence Pathway in Streptococcus pneumoniae
4. Bacteriocin Regulation and Function in S. pneumoniae
5. A Second Bacteriocin QS System Cryptically Linked to Competence in S. pneumoniae
6. A Direct Link between blp and com Is Established in S. pneumoniae
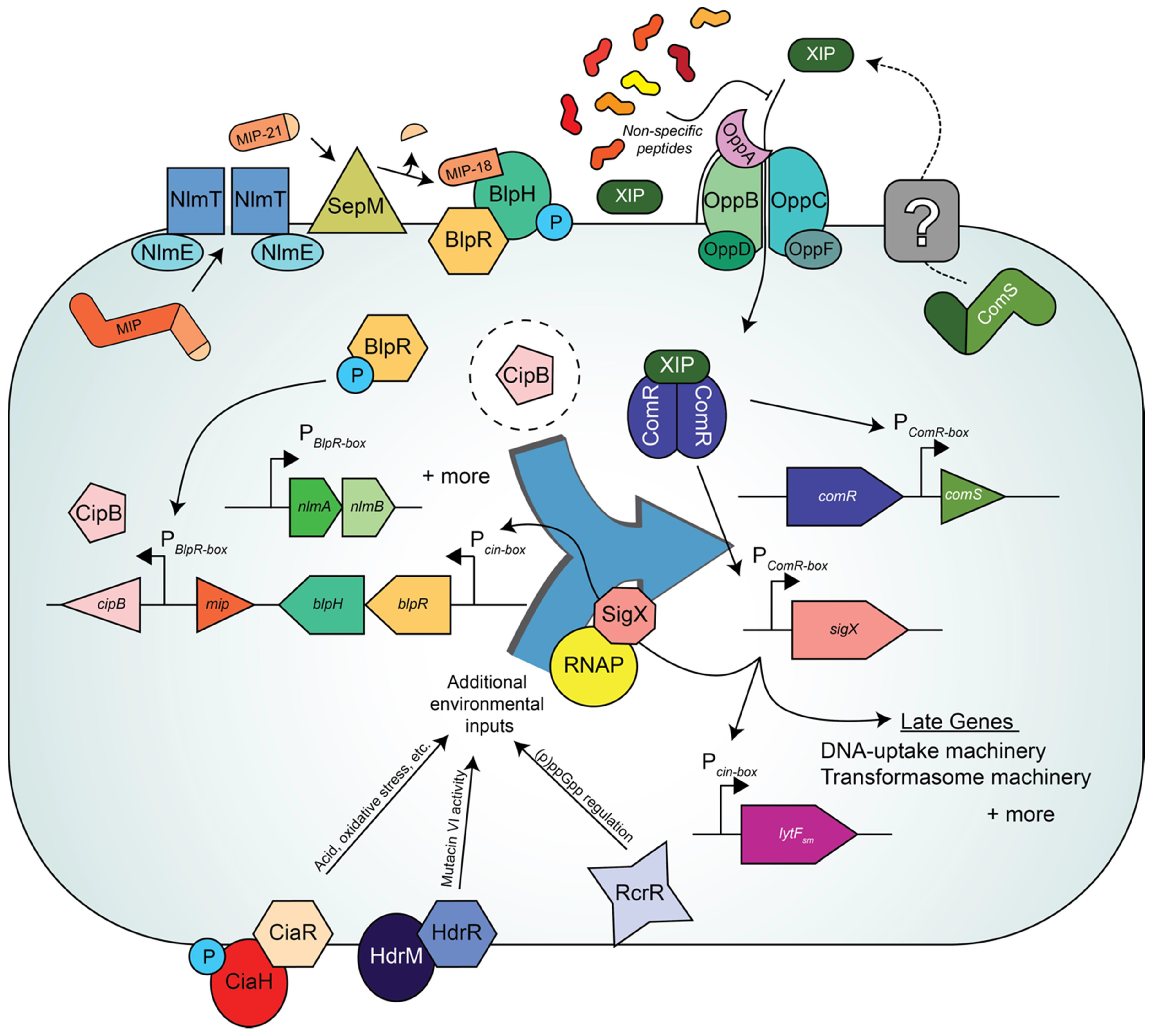
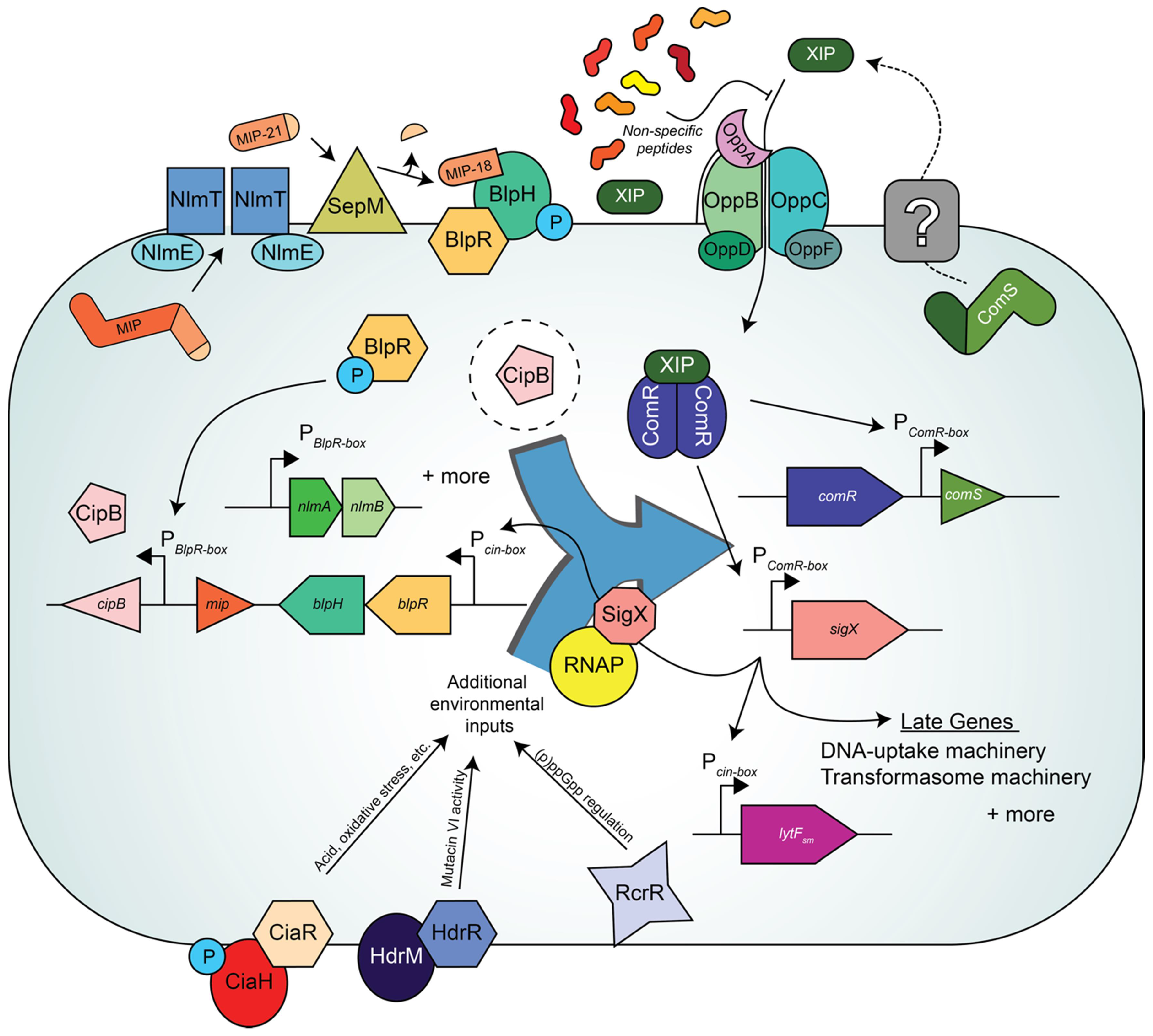
7. The Multiple Layers of Competence Regulation in Streptococcus mutans
8. How Does Peptide-Rich Versus Peptide-Poor Medium Affect Competence and Bacteriocin QS Pathways?: “The Medium Is the Message”—Marshall McLuhan [78]
9. Steps towards Resolving the Link between BlpRH and ComRS in S. mutans
10. Additional Inputs Regulating Competence in S. mutans
11. Uncovering Conservation Patterns of Competence and Bacteriocin QS Pathways in Streptococcus: Further Indications of Regulatory Interconnectedness?
12. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Griffith, F. The Significance of Pneumococcal Types. J. Hyg. (Lond) 1928, 27, 113–159. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Monnet, V.; Juillard, V.; Gardan, R. Peptide conversations in Gram-positive bacteria. Crit. Rev. Microbiol. 2014, 42, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.; Martin, B.; Fichant, G.; Polard, P.; Claverys, J.P. Bacterial transformation: distribution, shared mechanisms and divergent control. Nat. Rev. Microbiol. 2014, 12, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Perez-Pascual, D.; Monnet, V.; Gardan, R. Bacterial Cell-Cell Communication in the Host via RRNPP Peptide-Binding Regulators. Front. Microbiol. 2016, 7, 706. [Google Scholar] [CrossRef] [PubMed]
- Mell, J.C.; Redfield, R.J. Natural competence and the evolution of DNA uptake specificity. J. Bacteriol. 2014, 196, 1471–1483. [Google Scholar] [CrossRef] [PubMed]
- Havarstein, L.S.; Coomaraswamy, G.; Morrison, D.A. An unmodified heptadecapeptide pheromone induces competence for genetic transformation in Streptococcus pneumoniae. Proc. Natl. Acad. Sci. USA 1995, 92, 11140–11144. [Google Scholar] [CrossRef] [PubMed]
- Eijsink, V.G.; Axelsson, L.; Diep, D.B.; Havarstein, L.S.; Holo, H.; Nes, I.F. Production of class II bacteriocins by lactic acid bacteria; an example of biological warfare and communication. Antonie Leeuwenhoek 2002, 81, 639–654. [Google Scholar] [CrossRef] [PubMed]
- Håvarstein, L.S.; Holo, H.; Nes, I.F. The leader peptide of colicin V shares consensus sequences with leader peptides that are common among peptide bacteriocins produced by Gram-positive bacteria. Microbiology 1994, 140, 2383–2389. [Google Scholar] [CrossRef] [PubMed]
- Havarstein, L.S.; Diep, D.B.; Nes, I.F. A family of bacteriocin ABC transporters carry out proteolytic processing of their substrates concomitant with export. Mol. Microbiol. 1995, 16, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.S.; Biswas, I. An extracelluar protease, SepM, generates functional competence-stimulating peptide in Streptococcus mutans UA159. J. Bacteriol. 2012, 194, 5886–5896. [Google Scholar] [CrossRef] [PubMed]
- Havarstein, L.S.; Gaustad, P.; Nes, I.F.; Morrison, D.A. Identification of the streptococcal competence-pheromone receptor. Mol. Microbiol. 1996, 21, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Pestova, E.V.; Havarstein, L.S.; Morrison, D.A. Regulation of competence for genetic transformation in Streptococcus pneumoniae by an auto-induced peptide pheromone and a two-component regulatory system. Mol. Microbiol. 1996, 21, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.; Soulet, A.L.; Mirouze, N.; Prudhomme, M.; Mortier-Barriere, I.; Granadel, C.; Noirot-Gros, M.F.; Noirot, P.; Polard, P.; Claverys, J.P. ComE/ComE~P interplay dictates activation or extinction status of pneumococcal X-state (competence). Mol. Microbiol. 2013, 87, 394–411. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Morrison, D.A. Identification of a new regulator in Streptococcus pneumoniae linking quorum sensing to competence for genetic transformation. J. Bacteriol. 1999, 181, 5004–5016. [Google Scholar] [PubMed]
- Martin, B.; Granadel, C.; Campo, N.; Henard, V.; Prudhomme, M.; Claverys, J.P. Expression and maintenance of ComD-ComE, the two-component signal-transduction system that controls competence of Streptococcus pneumoniae. Mol. Microbiol. 2010, 75, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zhu, L.; Lau, G.W. Disentangling competence for genetic transformation and virulence in Streptococcus pneumoniae. Curr. Genet. 2016, 62, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, L.; Wahl, A.; Flechard, M.; Mignolet, J.; Hols, P. Regulation of competence for natural transformation in streptococci. Infect. Genet. Evol. 2015, 33, 343–360. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.; Cline, R.T.; Tettelin, H.; Sharov, V.; Morrison, D.A. Gene expression analysis of the Streptococcus pneumoniae competence regulons by use of DNA microarrays. J. Bacteriol. 2000, 182, 6192–6202. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.N.; Sung, C.K.; Cline, R.; Desai, B.V.; Snesrud, E.C.; Luo, P.; Walling, J.; Li, H.Y.; Mintz, M.; Tsegaye, G.; et al. Identification of competence pheromone responsive genes in Streptococcus pneumoniae by use of DNA microarrays. Mol. Microbiol. 2004, 51, 1051–1070. [Google Scholar] [CrossRef] [PubMed]
- Ween, O.; Gaustad, P.; Havarstein, L.S. Identification of DNA binding sites for ComE, a key regulator of natural competence in Streptococcus pneumoniae. Mol. Microbiol. 1999, 33, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.A.; Jones, M.B.; Peterson, S.N.; Cvitkovitch, D.G.; Levesque, C.M. Peptide alarmone signalling triggers an auto-active bacteriocin necessary for genetic competence. Mol. Microbiol. 2009, 72, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Hale, J.D.; Ting, Y.T.; Jack, R.W.; Tagg, J.R.; Heng, N.C. Bacteriocin (mutacin) production by Streptococcus mutans genome sequence reference strain UA159: Elucidation of the antimicrobial repertoire by genetic dissection. Appl. Environ. Microbiol. 2005, 71, 7613–7617. [Google Scholar] [CrossRef] [PubMed]
- van der Ploeg, J.R. Regulation of bacteriocin production in Streptococcus mutans by the quorum-sensing system required for development of genetic competence. J. Bacteriol. 2005, 187, 3980–3989. [Google Scholar] [CrossRef] [PubMed]
- Reck, M.; Tomasch, J.; Wagner-Dobler, I. The Alternative Sigma Factor SigX Controls Bacteriocin Synthesis and Competence, the Two Quorum Sensing Regulated Traits in Streptococcus mutans. PLoS Genet. 2015, 11, e1005353. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Lau, P.C.; Lee, J.H.; Ellen, R.P.; Cvitkovitch, D.G. Natural genetic transformation of Streptococcus mutans growing in biofilms. J. Bacteriol. 2001, 183, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Hale, J.D.; Heng, N.C.; Jack, R.W.; Tagg, J.R. Identification of nlmTE, the locus encoding the ABC transport system required for export of nonlantibiotic mutacins in Streptococcus mutans. J. Bacteriol. 2005, 187, 5036–5039. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Kuramitsu, H.K. Multiple streptococcus mutans genes are involved in biofilm formation. Appl. Environ. Microbiol. 2002, 68, 6283–6291. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.; Quentin, Y.; Fichant, G.; Claverys, J.P. Independent evolution of competence regulatory cascades in streptococci? Trends Microbiol. 2006, 14, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Tang, N.; Aspiras, M.B.; Lau, P.C.; Lee, J.H.; Ellen, R.P.; Cvitkovitch, D.G. A quorum-sensing signaling system essential for genetic competence in Streptococcus mutans is involved in biofilm formation. J. Bacteriol. 2002, 184, 2699–2708. [Google Scholar] [CrossRef] [PubMed]
- Mashburn-Warren, L.; Morrison, D.A.; Federle, M.J. A novel double-tryptophan peptide pheromone controls competence in Streptococcus spp. via an Rgg regulator. Mol. Microbiol. 2010, 78, 589–606. [Google Scholar] [CrossRef] [PubMed]
- Petersen, F.C.; Scheie, A.A. Genetic transformation in Streptococcus mutans requires a peptide secretion-like apparatus. Oral Microbiol. Immunol. 2000, 15, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Wholey, W.Y.; Kochan, T.J.; Storck, D.N.; Dawid, S. Coordinated Bacteriocin Expression and Competence in Streptococcus pneumoniae Contributes to Genetic Adaptation through Neighbor Predation. PLoS Pathog. 2016, 12, e1005413. [Google Scholar] [CrossRef] [PubMed]
- Kjos, M.; Miller, E.; Slager, J.; Lake, F.B.; Gericke, O.; Roberts, I.S.; Rozen, D.E.; Veening, J.W. Expression of Streptococcus pneumoniae Bacteriocins Is Induced by Antibiotics via Regulatory Interplay with the Competence System. PLoS Pathog. 2016, 12, e1005422. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Li, H.; Morrison, D.A. Identification of ComW as a new component in the regulation of genetic transformation in Streptococcus pneumoniae. Mol. Microbiol. 2004, 54, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Sung, C.K.; Morrison, D.A. Two distinct functions of ComW in stabilization and activation of the alternative sigma factor ComX in Streptococcus pneumoniae. J. Bacteriol. 2005, 187, 3052–3061. [Google Scholar] [CrossRef] [PubMed]
- Tovpeko, Y.; Morrison, D.A. Competence for genetic transformation in Streptococcus pneumoniae: mutations in sigmaA bypass the comW requirement. J. Bacteriol. 2014, 196, 3724–3734. [Google Scholar] [CrossRef] [PubMed]
- de Saizieu, A.; Gardes, C.; Flint, N.; Wagner, C.; Kamber, M.; Mitchell, T.J.; Keck, W.; Amrein, K.E.; Lange, R. Microarray-based identification of a novel Streptococcus pneumoniae regulon controlled by an autoinduced peptide. J. Bacteriol. 2000, 182, 4696–4703. [Google Scholar] [CrossRef] [PubMed]
- Havarstein, L.S.; Martin, B.; Johnsborg, O.; Granadel, C.; Claverys, J.P. New insights into the pneumococcal fratricide: relationship to clumping and identification of a novel immunity factor. Mol. Microbiol. 2006, 59, 1297–1307. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, E.; Ween, O.; Havarstein, L.S. Two separate quorum-sensing systems upregulate transcription of the same ABC transporter in Streptococcus pneumoniae. J. Bacteriol. 2004, 186, 3078–3085. [Google Scholar] [CrossRef] [PubMed]
- Claverys, J.P.; Martin, B.; Polard, P. The genetic transformation machinery: composition, localization, and mechanism. FEMS Microbiol. Rev. 2009, 33, 643–656. [Google Scholar] [CrossRef] [PubMed]
- Steinmoen, H.; Knutsen, E.; Havarstein, L.S. Induction of natural competence in Streptococcus pneumoniae triggers lysis and DNA release from a subfraction of the cell population. Proc. Natl. Acad. Sci. USA 2002, 99, 7681–7686. [Google Scholar] [CrossRef] [PubMed]
- Steinmoen, H.; Teigen, A.; Havarstein, L.S. Competence-induced cells of Streptococcus pneumoniae lyse competence-deficient cells of the same strain during cocultivation. J. Bacteriol. 2003, 185, 7176–7183. [Google Scholar] [CrossRef] [PubMed]
- Guiral, S.; Mitchell, T.J.; Martin, B.; Claverys, J.P. Competence-programmed predation of noncompetent cells in the human pathogen Streptococcus pneumoniae: genetic requirements. Proc. Natl. Acad. Sci. USA 2005, 102, 8710–8715. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Hill, C.; Ross, R.P. Bacteriocins: developing innate immunity for food. Nat. Rev. Microbiol. 2005, 3, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Diep, D.B.; Skaugen, M.; Salehian, Z.; Holo, H.; Nes, I.F. Common mechanisms of target cell recognition and immunity for class II bacteriocins. Proc. Natl. Acad. Sci. USA 2007, 104, 2384–2389. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Sieiro, P.; Montalban-Lopez, M.; Mu, D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Puelles, J.M.; Ronda, C.; Garcia, J.L.; Garcia, P.; Lopez, R.; Garcia, E. Searching for autolysin functions. Characterization of a pneumococcal mutant deleted in the lytA gene. Eur. J. Biochem. 1986, 158, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Johnsborg, O.; Havarstein, L.S. Regulation of natural genetic transformation and acquisition of transforming DNA in Streptococcus pneumoniae. FEMS Microbiol. Rev. 2009, 33, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Berg, K.H.; Biornstad, T.J.; Johnsborg, O.; Havarstein, L.S. Properties and biological role of streptococcal fratricins. Appl. Environ. Microbiol. 2012, 78, 3515–3522. [Google Scholar] [CrossRef]
- Throup, J.P.; Koretke, K.K.; Bryant, A.P.; Ingraham, K.A.; Chalker, A.F.; Ge, Y.; Marra, A.; Wallis, N.G.; Brown, J.R.; Holmes, D.J.; et al. A genomic analysis of two-component signal transduction in Streptococcus pneumoniae. Mol. Microbiol. 2000, 35, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Son, M.R.; Shchepetov, M.; Adrian, P.V.; Madhi, S.A.; de Gouveia, L.; von Gottberg, A.; Klugman, K.P.; Weiser, J.N.; Dawid, S. Conserved mutations in the pneumococcal bacteriocin transporter gene, blpA, result in a complex population consisting of producers and cheaters. MBio 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Reichmann, P.; Hakenbeck, R. Allelic variation in a peptide-inducible two-component system of Streptococcus pneumoniae. FEMS Microbiol. Lett. 2000, 190, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Dawid, S.; Roche, A.M.; Weiser, J.N. The blp bacteriocins of Streptococcus pneumoniae mediate intraspecies competition both in vitro and in vivo. Infect. Immun. 2007, 75, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Prudhomme, M.; Attaiech, L.; Sanchez, G.; Martin, B.; Claverys, J.P. Antibiotic stress induces genetic transformability in the human pathogen Streptococcus pneumoniae. Science 2006, 313, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Slager, J.; Kjos, M.; Attaiech, L.; Veening, J.W. Antibiotic-induced replication stress triggers bacterial competence by increasing gene dosage near the origin. Cell 2014, 157, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Stevens, K.E.; Chang, D.; Zwack, E.E.; Sebert, M.E. Competence in Streptococcus pneumoniae is regulated by the rate of ribosomal decoding errors. MBio 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Lange, R.; Wagner, C.; de Saizieu, A.; Flint, N.; Molnos, J.; Stieger, M.; Caspers, P.; Kamber, M.; Keck, W.; Amrein, K.E. Domain organization and molecular characterization of 13 two-component systems identified by genome sequencing of Streptococcus pneumoniae. Gene 1999, 237, 223–234. [Google Scholar] [CrossRef]
- Lemme, A.; Grobe, L.; Reck, M.; Tomasch, J.; Wagner-Dobler, I. Subpopulation-specific transcriptome analysis of competence-stimulating-peptide-induced Streptococcus mutans. J. Bacteriol. 2011, 193, 1863–1877. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, L.; Boutry, C.; de Frahan, M.H.; Delplace, B.; Fremaux, C.; Horvath, P.; Boyaval, P.; Hols, P. A novel pheromone quorum-sensing system controls the development of natural competence in Streptococcus thermophilus and Streptococcus salivarius. J. Bacteriol. 2010, 192, 1444–1454. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Rukke, H.V.; Ricomini Filho, A.P.; Fimland, G.; Arntzen, M.O.; Thiede, B.; Petersen, F.C. Extracellular identification of a processed type II ComR/ComS pheromone of Streptococcus mutans. J. Bacteriol. 2012, 194, 3781–3788. [Google Scholar] [CrossRef] [PubMed]
- Desai, K.; Mashburn-Warren, L.; Federle, M.J.; Morrison, D.A. Development of competence for genetic transformation of Streptococcus mutans in a chemically defined medium. J. Bacteriol. 2012, 194, 3774–3780. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, L.; Goffin, P.; Dubout, H.; Delplace, B.; Baulard, A.; Lecat-Guillet, N.; Chambellon, E.; Gardan, R.; Hols, P. Mechanism of competence activation by the ComRS signalling system in streptococci. Mol. Microbiol. 2013, 87, 1113–1132. [Google Scholar] [CrossRef] [PubMed]
- Okinaga, T.; Xie, Z.; Niu, G.; Qi, F.; Merritt, J. Examination of the hdrRM regulon yields insight into the competence system of Streptococcus mutans. Mol. Oral Microbiol. 2010, 25, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Gardan, R.; Besset, C.; Gitton, C.; Guillot, A.; Fontaine, L.; Hols, P.; Monnet, V. Extracellular life cycle of ComS, the competence-stimulating peptide of Streptococcus thermophilus. J. Bacteriol. 2013, 195, 1845–1855. [Google Scholar] [CrossRef] [PubMed]
- Shanker, E.; Morrison, D.A.; Talagas, A.; Nessler, S.; Federle, M.J.; Prehna, G. Pheromone Recognition and Selectivity by ComR Proteins among Streptococcus Species. PLoS Pathog. 2016, 12, e1005979. [Google Scholar] [CrossRef] [PubMed]
- Talagas, A.; Fontaine, L.; Ledesma-Garca, L.; Mignolet, J.; Li de la Sierra-Gallay, I.; Lazar, N.; Aumont-Nicaise, M.; Federle, M.J.; Prehna, G.; Hols, P.; et al. Structural Insights into Streptococcal Competence Regulation by the Cell-to-Cell Communication System ComRS. PLoS Pathog. 2016, 12, e1005980. [Google Scholar] [CrossRef] [PubMed]
- Dufour, D.; Levesque, C.M. Cell death of Streptococcus mutans induced by a quorum-sensing peptide occurs via a conserved streptococcal autolysin. J. Bacteriol. 2013, 195, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Gardan, R.; Besset, C.; Guillot, A.; Gitton, C.; Monnet, V. The oligopeptide transport system is essential for the development of natural competence in Streptococcus thermophilus strain LMD-9. J. Bacteriol. 2009, 191, 4647–4655. [Google Scholar] [CrossRef] [PubMed]
- Kreth, J.; Hung, D.C.; Merritt, J.; Perry, J.; Zhu, L.; Goodman, S.D.; Cvitkovitch, D.G.; Shi, W.; Qi, F. The response regulator ComE in Streptococcus mutans functions both as a transcription activator of mutacin production and repressor of CSP biosynthesis. Microbiology 2007, 153, 1799–1807. [Google Scholar] [CrossRef] [PubMed]
- Dufour, D.; Cordova, M.; Cvitkovitch, D.G.; Levesque, C.M. Regulation of the competence pathway as a novel role associated with a streptococcal bacteriocin. J. Bacteriol. 2011, 193, 6552–6559. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.J.; Wen, Z.T.; Burne, R.A. Multilevel control of competence development and stress tolerance in Streptococcus mutans UA159. Infect. Immun. 2006, 74, 1631–1642. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Merritt, J.; Lux, R.; Shi, W. Inactivation of the ciaH gene in Streptococcus mutans diminishes mutacin production and competence development, alters sucrose-dependent biofilm formation, and reduces stress tolerance. Infect. Immun. 2004, 72, 4895–4899. [Google Scholar] [CrossRef] [PubMed]
- Levesque, C.M.; Mair, R.W.; Perry, J.A.; Lau, P.C.; Li, Y.H.; Cvitkovitch, D.G. Systemic inactivation and phenotypic characterization of two-component systems in expression of Streptococcus mutans virulence properties. Lett. Appl. Microbiol. 2007, 45, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Merritt, J.; Zheng, L.; Shi, W.; Qi, F. Genetic characterization of the hdrRM operon: a novel high-cell-density-responsive regulator in Streptococcus mutans. Microbiology 2007, 153, 2765–2773. [Google Scholar] [CrossRef] [PubMed]
- Seaton, K.; Ahn, S.J.; Burne, R.A. Regulation of competence and gene expression in Streptococcus mutans by the RcrR transcriptional regulator. Mol. Oral Microbiol. 2015, 30, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Seaton, K.; Ahn, S.J.; Sagstetter, A.M.; Burne, R.A. A transcriptional regulator and ABC transporters link stress tolerance, (p)ppGpp, and genetic competence in Streptococcus mutans. J. Bacteriol. 2011, 193, 862–874. [Google Scholar] [CrossRef] [PubMed]
- McLuhan, M. Understanding Media; The Extensions of Man, 1st ed.; McGraw-Hill: New York, NY, USA, 1964. [Google Scholar]
- Son, M.; Ahn, S.J.; Guo, Q.; Burne, R.A.; Hagen, S.J. Microfluidic study of competence regulation in Streptococcus mutans: Environmental inputs modulate bimodal and unimodal expression of comX. Mol. Microbiol. 2012, 86, 258–272. [Google Scholar] [CrossRef] [PubMed]
- Petersen, F.C.; Fimland, G.; Scheie, A.A. Purification and functional studies of a potent modified quorum-sensing peptide and a two-peptide bacteriocin in Streptococcus mutans. Mol. Microbiol. 2006, 61, 1322–1334. [Google Scholar] [CrossRef] [PubMed]
- Okinaga, T.; Niu, G.; Xie, Z.; Qi, F.; Merritt, J. The hdrRM operon of Streptococcus mutans encodes a novel regulatory system for coordinated competence development and bacteriocin production. J. Bacteriol. 2010, 192, 1844–1852. [Google Scholar] [CrossRef] [PubMed]
- Aspiras, M.B.; Ellen, R.P.; Cvitkovitch, D.G. ComX activity of Streptococcus mutans growing in biofilms. FEMS Microbiol. Lett. 2004, 238, 167–174. [Google Scholar] [PubMed]
- Son, M.; Shields, R.C.; Ahn, S.J.; Burne, R.A.; Hagen, S.J. Bidirectional signaling in the competence regulatory pathway of Streptococcus mutans. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef] [PubMed]
- Barbour, A.; Tagg, J.; Abou-Zied, O.K.; Philip, K. New insights into the mode of action of the lantibiotic salivaricin B. Sci. Rep. 2016, 6, 31749. [Google Scholar] [CrossRef] [PubMed]
- Siryaporn, A.; Goulian, M. Characterizing cross-talk in vivo avoiding pitfalls and overinterpretation. Methods Enzymol. 2010, 471, 1–16. [Google Scholar] [PubMed]
- Kaspar, J.; Kim, J.N.; Ahn, S.J.; Burne, R.A. An essential role for (p)ppGpp in the integration of stress tolerance, peptide signaling, and competence development in Streptococcus mutans. Front. Microbiol. 2016, 7, 1162. [Google Scholar] [CrossRef] [PubMed]
- Dagkessamanskaia, A.; Moscoso, M.; Henard, V.; Guiral, S.; Overweg, K.; Reuter, M.; Martin, B.; Wells, J.; Claverys, J.P. Interconnection of competence, stress and CiaR regulons in Streptococcus pneumoniae: competence triggers stationary phase autolysis of ciaR mutant cells. Mol. Microbiol. 2004, 51, 1071–1086. [Google Scholar] [CrossRef] [PubMed]
- Trappetti, C.; Potter, A.J.; Paton, A.W.; Oggioni, M.R.; Paton, J.C. LuxS mediates iron-dependent biofilm formation, competence, and fratricide in Streptococcus pneumoniae. Infect. Immun. 2011, 79, 4550–4558. [Google Scholar] [CrossRef] [PubMed]
- Mashburn-Warren, L.; Morrison, D.A.; Federle, M.J. The cryptic competence pathway in Streptococcus pyogenes is controlled by a peptide pheromone. J. Bacteriol. 2012, 194, 4589–4600. [Google Scholar] [CrossRef] [PubMed]
- Morrison, D.A.; Guedon, E.; Renault, P. Competence for natural genetic transformation in the Streptococcus bovis group streptococci S. infantarius and S. macedonicus. J. Bacteriol. 2013, 195, 2612–2620. [Google Scholar] [CrossRef] [PubMed]
- Zaccaria, E.; van Baarlen, P.; de Greeff, A.; Morrison, D.A.; Smith, H.; Wells, J.M. Control of competence for DNA transformation in Streptococcus suis by genetically transferable pherotypes. PLoS ONE 2014, 9, e99394. [Google Scholar] [CrossRef] [PubMed]
- Marks, L.R.; Mashburn-Warren, L.; Federle, M.J.; Hakansson, A.P. Streptococcus pyogenes biofilm growth in vitro and in vivo and its role in colonization, virulence, and genetic exchange. J. Infect. Dis. 2014, 210, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Zaccaria, E.; Wells, J.M.; van Baarlen, P. Metabolic Context of the Competence-Induced Checkpoint for Cell Replication in Streptococcus suis. PLoS ONE 2016, 11, e0153571. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Rukke, H.V.; Høvik, H.; Åmdal, H.A.; Chen, T.; Morrison, D.A.; Petersen, F.C.; McFall-Ngai, M.J. Comprehensive transcriptome profiles of Streptococcus mutans UA159 map core streptococcal competence genes. mSystems 2016, 1, e00038-15. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| S. mutans Locus ID | Gene Product Name(s) | Function | References |
|---|---|---|---|
| SMU.1914c | CipB, Mutacin V, NlmC, BsmA | Type II bacteriocin | [22,23,24] |
| SMU.1915 | MIP, ComC/CSP | Type II bacteriocin/pheromone | [25,26] |
| SMU.286 | NlmT, ComA | Bacteriocin transporter | [11,27,28] |
| SMU.287 | NlmE, ComB | NlmT accessory factor | [11,27,28] |
| SMU.1916 | BlpH, ComD | Histidine kinase | [26,29] |
| SMU.1917 | BlpR, ComE | Response regulator | [26,29] |
| SMU.1997 | SigX, ComX | Alternative sigma factor | [30,31] |
| SMU.61 | ComR | Type-II ComR; Transcriptional regulator of competence | [31] |
| N/A | ComS | Pre-peptide of XIP pheromone | [31] |
| SMU.1897-.1898 | CslA, ComA | ABC transporter ATP-binding protein/permease | [30,32] |
| SMU.1900 | CslB, ComB | ABC transporter accessory protein | [30,32] |
| SMU.381c | Smu.381c | Type-I ComR; Putative transcriptional regulator | [31] |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shanker, E.; Federle, M.J. Quorum Sensing Regulation of Competence and Bacteriocins in Streptococcus pneumoniae and mutans. Genes 2017, 8, 15. https://doi.org/10.3390/genes8010015
Shanker E, Federle MJ. Quorum Sensing Regulation of Competence and Bacteriocins in Streptococcus pneumoniae and mutans. Genes. 2017; 8(1):15. https://doi.org/10.3390/genes8010015
Chicago/Turabian StyleShanker, Erin, and Michael J. Federle. 2017. "Quorum Sensing Regulation of Competence and Bacteriocins in Streptococcus pneumoniae and mutans" Genes 8, no. 1: 15. https://doi.org/10.3390/genes8010015
APA StyleShanker, E., & Federle, M. J. (2017). Quorum Sensing Regulation of Competence and Bacteriocins in Streptococcus pneumoniae and mutans. Genes, 8(1), 15. https://doi.org/10.3390/genes8010015