Regulation of Innate Immune Responses by Bovine Herpesvirus 1 and Infected Cell Protein 0 (bICP0)

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
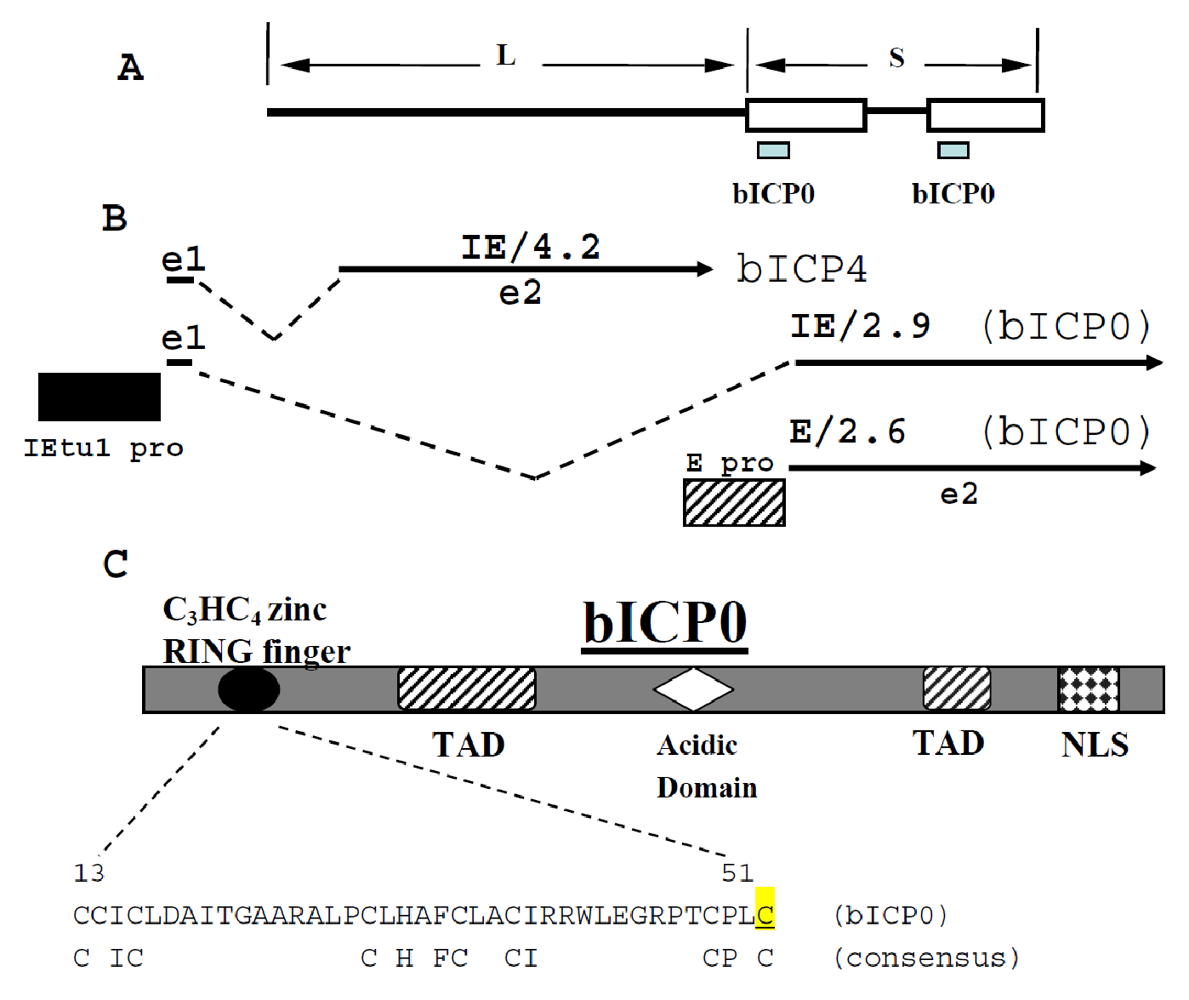
2.1. bICP0 is encoded within immediate early transcript unit 1 (IEtu1)

2.2. bICP0 contains multiple functional domains
2.3. bICP0 interacts with chromatin remodeling enzymes
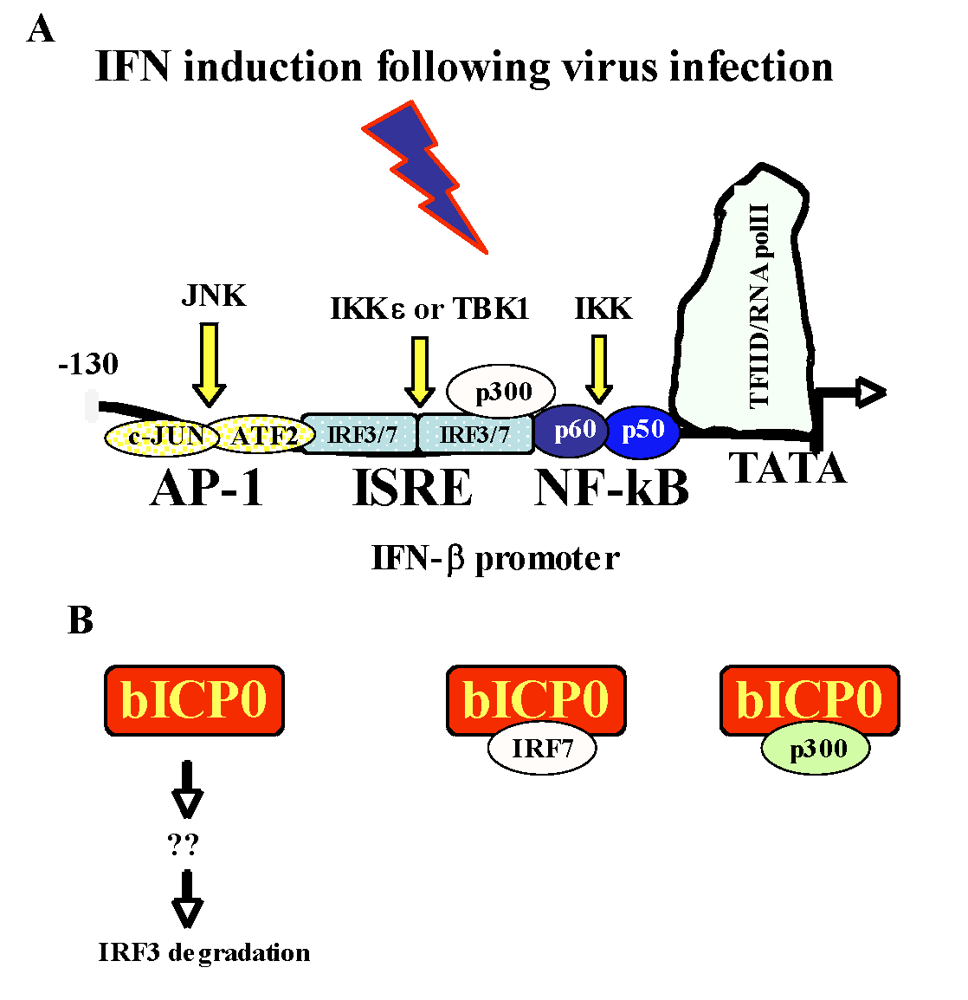
2.4. Activation of the interferon pathway following infection
2.5. bICP0 inhibits beta interferon promoter (IFN-β) promoter activity
2.5.a. Activation of the IFN-β promoter is an early event after virus infection

2.5.b. bICP0 inhibits IRF3 functions by inducing its degradation

2.5.c. bICP0 interacts with IRF7
2.6. Growth properties of bICP0 mutant viruses
2.7. Regulation of immune responses by other BoHV-1 genes
2.7.a. Inhibition of CD8+ T cell recognition in infected cells
2.7.b. Inhibition of CD4+ T cell functions
2.7.c. BoHV-1 encodes a protein that interacts with chemokines
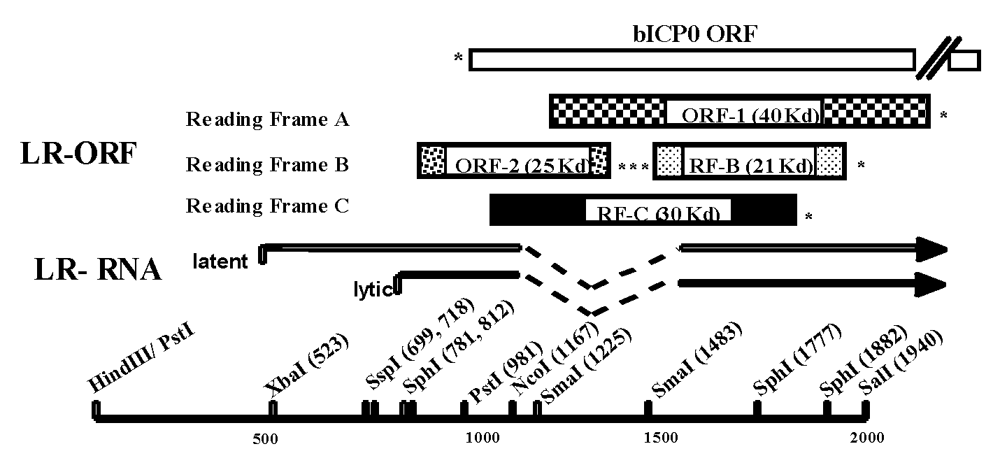
2.7.d. LR RNA can influence immune responses
3. Conclusions
Acknowledgments
References and Notes
- 1995-96 Agricultural Statistics; National Agricultural Statistics Service (NASS): Agricultural Statistics Board, U.S. Department of Agriculture, 1996.
- Aaronson, D.S.; Horvath, C.M. A road map for those who don't know JAK-STAT. Science 2002, 296, 1653–1655. [Google Scholar] [CrossRef] [PubMed]
- Abril, C.; Engels, M.; Limman, A.; Hilbe, M.; Albini, S.; Franchini,M.; Suter, M.; Ackerman, M. Both viral and host factors contribute to neurovirulence of bovine herpesvirus 1 and 5 in interferon receptor-deficient mice. J. Virol. 2004, 78, 3644–3653. [Google Scholar] [CrossRef] [PubMed]
- Aderem, A.; Hume, D.A. How do you see CG? Cell 2000, 103, 993–996. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.; Meyer, T.H.; Uebel, S.; Sempe, P.; Djaballah, H.; Yang, Y.; Peterson, P.A.; Fruh, K.; Tampe, R. Molecular mechanism and species specificity of TAP inhibition by herpes simplex virus ICP47. EMBO J. 1996, 15, 3247–3255. [Google Scholar] [PubMed]
- Akdemir, F.; Farkas, R.; Chen, P.; Juhasz, G.; Medved'ova, L; Saas, M.; Wang, L.; Wang, X.; Chittaranjan, S.; Gorski, S.M.; Rodriguez, A.; Abrams, J. M. Autophagy occurs upstream or parallel to the apoptosome during histolytic cell death. Development 2006, 133, 1457–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Ambagala, A.P.; Gopinath, R.S.; Srikumaran, S. Peptide transport activity of the transporter associated with antigen processing (TAP) is inhibited by an early protein of equine herpesvirus-1. J. Gen. Virol. 2004, 66, 2383–2394. [Google Scholar]
- Au, W.C.; Moore, P.A.; LaFleur, D.W.; Tombal, B.; Pitha, P.M. Characterization of the interferon regulatory factor-7 and its potential role in the transcription activation of interferon A genes. J. Biol. Chem. 1998, 273, 29210–29217. [Google Scholar] [CrossRef] [PubMed]
- Ayer, D.E.; Kretzner, L.; Eisenman, R.N. Mad: a heterodimeric partner for Max that antagonizes Myc transcriptional activity. Cell 1993, 72, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Baehrecke, E.H. Autophagy: dual roles in life and death? Nat. Rev. Molec. Cell Biol. 2006, 6, 505–510. [Google Scholar] [CrossRef]
- Baggiolini, M. Chemokines and leukocyte traffic. Nature 1998, 392, 565–568. [Google Scholar] [CrossRef] [PubMed]
- Boutell, C.; Everett, R.D. The herpes simplex virus type 1 (HSV-1) regulatory protein ICP0 interacts with an ubiquitinates p53. J. Biol. Chem. 2003, 278, 36596–36602. [Google Scholar] [CrossRef] [PubMed]
- Boutell, C.; Sadis, S.; Everett, R.D. Herpes simplex virus type 1 immediate-early protein ICP0 and its isolated RING finger domain act as ubiquitin E3 ligases in vitro. J. Virol. 2002, 76, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Bowland, S.L.; Shewen, P.E. Bovine respiratory disease: commercial vaccines currently available in Canada. Can. Vet. J. 2000, 41, 33–48. [Google Scholar]
- Bowles, D.E.; Holden, V.R.; Zhao, Y.; O'Callaghan, D.J. The ICP0 protein of equine herpesvirus 1 is an early protein that independently transactivates expression of all classes of viral promoters. J. Virol. 1997, 71, 4904–4914. [Google Scholar] [PubMed]
- Bowles, D.E.; Kim, S.K.; O'Callaghan, D.J. Characterization of the trans-activation properties of equine herpesvirus 1 EICP0 protein. J. Virol. 2000, 74, 1200–1208. [Google Scholar] [CrossRef] [PubMed]
- Boyes, J.; Byfield, P.; Nakatani, Y.; Ogryzko, V. Regulation of activity of the transcription factor GATA-1 by acetylation. Nature 1998, 396, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Bratanich, A.C.; Hanson, N.D.; Jones, C. The latency-related gene of bovine herpesvirus 1 inhibits the activity of immediate-early transcription unit 1. Virology 1992, 191, 988–991. [Google Scholar] [CrossRef] [PubMed]
- Brukman, A; Enquist, L.W. Pseudorabies virus EP0 protein counteracts an interferon-induced antiviral state in a species-specific manner. J. Virol. 2006, 80, 10871–10873. [Google Scholar] [CrossRef] [PubMed]
- Bryant, N.A.; Davis-Poynter, N.; Vanderplasschen, A.; Alcami, A. Glycoprotein G isoforms from some alphaherpesvirus function as broad-spectrum chemokine binding proteins. EMBO J. 2003, 22, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Burch, W. The autophagosomal-lysosomal compartment in programmed cell death. Cell Death and Differentiation 2001, 8, 569–581. [Google Scholar] [CrossRef]
- Canning, M.; Boutell, C.; Parkinson, J.; Everett, R.D. A RING finger ubiquitin ligase activity is protected from autocatalyzed ubiquitination and degradation by binding to ubiquitin-specific protease USP27. J. Biol. Chem. 2004, 279, 38160–38168. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.J.; Weinberg, A.D.; Pollard, A.; Reeves, R.; Magnuson, J.A.; Magnuson, N.S. Inhibition of T-lymphocyte mitogenic responses and effects on cell functions by bovine herpesvirus 1. J. Virol. 1989, 63, 1525–1530. [Google Scholar] [PubMed]
- Chen, H.; Lin, R.J.; Xie, W. Regulation of hormone-induced histone hyperacetylation and gene activation via acetylation of an acetylase. Cell 1999, 98, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Ciacci-Zanella, J.; Stone, M.; Henderson, G.; Jones, C. The latency-related gene of bovine herpesvirus 1 inhibits programmed cell death. J. Virol. 1999, 73, 9734–9740. [Google Scholar] [PubMed]
- Coull, J.J.; Romerio, F.; Sun, J.M.; Volker, J.L.; Galvin, K.M.; Davie, J.R.; Shi, Y.; Hansen, U.; Margolis, D.M. The human factors YY1 and LSF repress the human immunodeficiency virus type 1 long terminal repeat via recruitment of histone deacetylase 1. J. Virol. 2000, 74, 6790–6799. [Google Scholar] [CrossRef] [PubMed]
- Devireddy, L.R.; Jones, C. Activation of caspases and p53 by bovine herpesvirus 1 infection results in programmed cell death and efficient virus release. J. Virol. 1999, 73, 3778–3788. [Google Scholar] [PubMed]
- Dia, L.; Zhang, B.; Fan, J.; Gao, X.; Sun, S.; Yang, K.; Xin, D.; Jin, N.; Geng, Y.; Wang, C. Herpes virus proteins ICP0 and BICP0 can activate NF-kB by catalyzing IkBa ubiquitination. Cellular Signalling 2005, 17, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Diao, L.; Qiao, W.; Chen, Q.; Wang, C.; Geng, Y. bICP0 and its RING domain act as ubiquitin E3 ligases in vitro. Chinese Science Bulletin 2005, 50, 636–640. [Google Scholar] [CrossRef]
- Doetzlhofer, A.; Rotheneder, H.; Koranda, M.; Kurtev, V.; Brosch, G.; Wintersberger, E.; Seiser, C. Histone deacetylase 1 can repress transcription by binding to Sp1. Mol. Cell. Biol. 1999, 19, 5504–5511. [Google Scholar] [PubMed]
- Doyle, S.; Vaidya, S.; O'Connell, R.; Dadgostar, H.; Dempsey, P.; Wu, T.; Rao, G.; Sun, R.; Haberland, M.; Modlin, R.; Cheng, G. IRF3 mediates a TLR3/TLR4-specific antiviral gene program. Immunity 2002, 17, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.; Waldner, C.; Rhodes, C.; Ricketts, V. Longevity of protective immunity to experimental bovine herpesvirus-1 infection following inoculation with a combination modified-live virus vaccine in beef calves. J. Am. Vet. Med. Assoc. 2005, 227, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Everett, R.; O'Hare, P.; O'Rourke, D.; Barlow, P.; Orr, A. Point mutations in the herpes simplex virus type 1 Vmw110 RING finger helix affect activation of gene expression, viral growth, and interaction with PML-containing nuclear structures. J. Virol. 1995, 69, 7339–7344. [Google Scholar] [PubMed]
- Everett, R.D. Analysis of the functional domains of herpes simplex virus type 1 immediate-early polypeptide Vmw110. J. Mol. Biol. 1988, 202, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Everett, R.D. ICP0, a regulator of herpes simplex virus during lytic and latent infection. Bioessays 2000, 22, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Everett, R.D.; Barlow, P.; Milner, A.; Luisi, B.; Orr, A.; Hope, G.; Lyon, D. A novel arrangement of zinc-binding residues and secondary structure in the C3HC4 motif of an alpha herpes virus protein family. J. Mol. Biol. 1993, 234, 1038–1047. [Google Scholar] [CrossRef] [PubMed]
- Everett, R.D.; Earnshaw, W.C.; Findlay, J.; Lomonte, P. Specific destruction of kinetochore protein CENP-C and disruption of cell division by herpes simplex virus immediate-early protein Vmw110. EMBO J. 1999, 18, 1526–1538. [Google Scholar] [CrossRef] [PubMed]
- Everett, R.D.; Lomonte, P.; Orr, A. Cell cycle regulation of PML modification and ND10 composition. J. Cell Sci. 1999, 112, 4581–4588. [Google Scholar] [PubMed]
- Everett, R.D.; Meredith, M.; Orr, A.; Cross, A.; Kathoria, M.; Parkinson, J. A novel ubiquitin-specific protease is dynamically associated with the PML nuclear domain and binds to a herpesvirus regulatory protein [corrected and republished article originally printed in EMBO J.1997, 16, 566-577]. EMBO J. 1997, 16, 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- Fischle, W.; Emiliani, S.; Hendzel, M.J.; Nagase, T.; Nomura, N.; Voelter, W.; Verdin, E. A new family of human histone deacetylases related to Saccharomyces cerevisiae HDA1p. J. Biol. Chem. 1999, 274, 11713–11720. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.A.; McWhirter, S.M.; Faja, K.L.; Rowe, D.C.; Latz, E.; Golenbock, D.T.; Coyle, A.J.; Liao, S.-M.; Maniatis, T. IKKe and TBKI are essential components of the IRF3 signaling pathway. Nature 2003, 4, 491–496. [Google Scholar]
- Fraefel, C.; Zeng, J.; Choffat, Y.; Engels, M.; Schwyzer, M.; Ackermann, M. Identification and zinc dependence of the bovine herpesvirus 1 transactivator protein BICP0. J. Virol. 1994, 68, 3154–3162. [Google Scholar] [PubMed]
- Geiser, V.; Jones, C. The latency related gene encoded by bovine herpesvirus 1 encodes a small regulatory RNA that inhibits cell growth. J. Neurovirol. 2005, 11, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Geiser, V.; Inman, M.; Zhang, Y.; Jones, C. The latency related (LR) gene of bovine herpes virus 1 (BHV-1) can inhibit the ability of bICP0 to activate productive infection. J. Gen. Virol. 2002, 83, 2965–2971. [Google Scholar] [PubMed]
- Geiser, V.; Rose, S.; Jones, C. The bovine herpes virus 1 bICP0 protein regulates toxicity in a cell type dependent fashion. Molec. Path. 2008, 44, 459–466. [Google Scholar]
- Geiser, V.; Zhang, Y. Characterization of a BHV-1 strain that does not express the major regulatory protein, bICP0. J. Gen. Virol. 2005, 86, 1987–1996. [Google Scholar] [CrossRef] [PubMed]
- Griebel, P.; Ohmann, H.B.; Lawman, M.J.; Babiuk, L.A. The interaction between bovine herpesvirus type 1 and activated bovine T lymphocytes. J. Gen. Virol. 1990, 71, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Griebel, P.; Qualtiere, L.; Davis, W.C.; Gee, A.; Bielefeldt Ohmann, H.; Lawman, M.J.; Babiuk, L.A. T lymphocyte population dynamics and function following a primary bovine herpesvirus type-1 infection. Viral. Immunol. 1987, 1, 287–304. [Google Scholar] [CrossRef]
- Griebel, P.J.; Qualtiere, L.; Davis, W.C.; Lawman, M.J.; Babiuk, L.A. Bovine peripheral blood leukocyte subpopulation dynamics following a primary bovine herpesvirus-1 infection. Viral Immunol. 1987, 1, 267–286. [Google Scholar] [CrossRef]
- Grozinger, C.M.; Hassig, C.A.; Schreiber, S.L. Three proteins define a class of human histone deacetylases related to yeast Hda1p. Proc. Natl. Acad. Sci. U.S.A. 1999, 96, 4868–4873. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Liang, Y.; Mandel, G.; Roizman, B. Components of the REST/CoREST/histone deacytlase repressor complex are disrupted, modified, and translocated in HSV-1-infected cells. Proc. Nat. Acad. Sci. U.S.A. 2005, 102, 7571–7576. [Google Scholar] [CrossRef]
- Gu, W.; Roeder, R.G. Activation of p53 sequence-specific DNA binding by acetylation of the p53 C-terminal domain. Cell 1997, 90, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Peters, K.L.; Sen, G.C. Induction of the human protein P56 by interferon, double stranded RNA, or virus infection. Virology 2000, 267, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Hacker, H.; Vabulas, R.M.; Takeuchi, O.; Hoshino, K.; Akira, S.; Wagner, H. Immune cell activation by bacterial CpG-DNA through myeloid differentiation marker 88 and tumor necrosis factor receptor-associated factor (TRAF) 6. J. Exp. Med. 2000, 192, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Hariharan, M.J.; Nataraj, C.; Srikumaran, S. Down regulation of murine MHC class I expression by bovine herpesvirus 1. Viral Immunol. 1993, 6, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Heinzel, T.; Lavinsky, R.M.; Mullen, T.M.; Soderstrom, M.; Laherty, C.D.; Torchia, J.; Yang, W.M.; Brard, G.; Ngo, S.D.; Davie, J.R.; Seto, E.; Eisenman, R.N.; Rose, D.W.; Glass, C.K.; Rosenfeld, M.G. A complex containing N-CoR, mSin3 and histone deacetylase mediates transcriptional repression [see comments]. Nature 1997, 387, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Perng, G.-C.; Nesburn, A.; Wechsler, S.; Jones, C. The latency related gene of bovine herpesvirus 1 can suppress caspase 3 and caspase 9 during productive infection. J. Neurovirol. 2004, 10, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Zhang, Y.; Jones, C. The bovine herpesvirus 1 gene encoding infected cell protein 0 (bICP0) can inhibit interferon-dependent transcription in the absence of other viral genes. J. Gen. Virol. 2005, 86, 2697–2702. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Zhang, Y.; Inman, M.; D. Jones, C. Jones. The infected cell protein 0 encoded by bovine herpes virus 1 (bICP0) can activate caspase 3 when over-expressed in transfected cells. J. Gen. Virol. 2004, 85, 3511–3516. [Google Scholar] [CrossRef] [PubMed]
- Hinkley, S.; Hill, A.B.; Srikumaran, S. Bovine herpesvirus-1 infection affects the peptide transport activity in bovine cells. Virus Res. 1998, 53, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, W.E.; DeLuca, N.A. 2nd; Perturbation of cell cycle progression and cellular gene expression as a function of herpes simplex virus ICP0. . J. Virol. 1999, 73, 8245–8255. [Google Scholar] [PubMed]
- Honda, K.; Yanai, H.; Negishi, H.; Asagiri, M.; Saton, M.; Mizutani, T.; Shimada, N.; Ohba, Y.; Takaoka, A.; Yoshida, N.; Taniguchi, T. IRF-7 is the master regulator of type-I interferon-dependent immune responses. Nature 2005, 434, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Schang, L.M.; Jones, C. Identification of gene products encoded by the latency-related gene of bovine herpesvirus 1. J. Virol. 1995, 69, 5345–5352. [Google Scholar] [PubMed]
- Hughes, E.A.; Hammond, C.; Cresswell, P. Misfolded major histocompatibilitycomplex class I heavy chains are translocated into the cytoplasm and degraded by the proteosome. Proc. Nat. Acad. Sci. 1997, 94, 1896–1901. [Google Scholar] [CrossRef]
- Irving, U.P. Reference of Dairy Health and Management in the United States; USDA:AHIS:VS, CEAH,National Animal Health Monitoring System, 2002. [Google Scholar]
- Imhof, A.; Yang, X.J.; Ogryzko, V.V.; Nakatani, Y.; Wolffe, A.P.; Ge, H. Acetylation of general transcription factors by histone acetyltransferases. Curr. Biol. 1997, 7, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Inman, M.; Lovato, L.; Doster, A.; Jones, C. A mutation in the latency related gene of bovine herpesvirus 1 interferes with the latency-reactivation cycle of latency in calves. J. Virol. 2002, 76, 6771–6779. [Google Scholar] [CrossRef] [PubMed]
- Inman, M.; Lovato, L.; Doster, A.; Jones, C. A mutation in the latency-related gene of bovine herpesvirus 1 leads to impaired ocular shedding in acutely infected calves. J. Virol. 2001, 75, 8507–8515. [Google Scholar] [CrossRef] [PubMed]
- Inman, M.; Zhang, Y.; Geiser, V.; Jones, C. The zinc ring finger in the bICP0 protein encoded by bovine herpes virus-1 mediates toxicity and activates productive infection. J. Gen. Virol. 2001, 82, 483–492. [Google Scholar] [PubMed]
- Ishmael, W. Gasping for dollars. Angus Beef Bulletin 2001, Available online: www.mycattle.com/health/updates/ gaspingfordollars/. [Google Scholar]
- Jaeschke, A.; Karasarides, M.; Ventura, J.; Ehrhardt, A.; Zhang, C.; Flavell, R.A.; Shokat, K.M.; Davis, R.J. JNK2 Is a Positive Regulator of the cJun Transcription Factor. Mol. Cell. 2006, 23, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Hossain, A.; Winkler, M.T.; Holt, T.; Doster, A.; Jones, C. A protein encoded by the latency-related gene of bovine herpesvirus 1 is expressed in trigeminal ganglionic neurons of latently infected cattle and interacts with cyclin-dependent kinase 2 during productive infection. J. Virol. 1998, 72, 8133–8142. [Google Scholar] [PubMed]
- Jiang, Y.; Inman, M.; Zhang, Y.; Posadas, N.A.; Jones, C. A mutation in the latency related gene of bovine herpesvirus 1 (BHV-1) inhibits protein expression of a protein from open reading frame 2 (ORF-2) and an adjacent reading frame during productive infection. J. Virol. 2004, 78, 3184–3189. [Google Scholar] [CrossRef] [PubMed]
- Jones, C. Alphaherpesvirus latency: its role in disease and survival of the virus in nature. Adv. Virus Res. 1998, 51, 81–133. [Google Scholar] [PubMed]
- Jones, C. Herpes simplex virus type 1 and bovine herpesvirus 1 latency. Clin. Micro. Rev. 2003, 16, 79–95. [Google Scholar] [CrossRef]
- Jones, C.; Geiser, V.; Henderson, G.; Jiang, Y.; Meyer, F.; Perez, S.; Zhang, Y. Functional analysis of bovine herpesvirus 1 (BHV-1) genes expressed during latency. Vet. Micro. 2006, 113, 199–210. [Google Scholar] [CrossRef]
- Kaashoek, M. J.; Fijsewijk, F.A.M.; Ruuls, R.C.; Keil, G.M.; Thiry, E.; Pastoret, P.P.; Van Oirschot, J.T. Virulence, immunogenicity and reactivation of bovine herpesvirus 1 mutants with a deletion in the gC, gG, gI, gE, or in both the gI and gE gene. Vaccine 1998, 16, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Kapil, S.; Basaraba, R.J. Infectious bovine rhinotracheitis, parainfluenza-3, and respiratory coronavirus. Bovine respiratory disease update. In Food Animal Practice; Veterinary Clinics of North America:, 1997; pp. 455–461. [Google Scholar]
- Katze, M.G.; Heng, Y.; Gale, M. Viruses and interferon: fight for supremacy. Nat. Rev. Immuno. 2002, 2, 675–686. [Google Scholar] [CrossRef]
- Koppers-Lalic, D.; Reits, E.A.J.; Ressing, M.E.; Lipinska, A.D.; Abele, R.; Koch, J.; Rezende, M.M.; Admiraal, P.; van Leeuwen, D.; Bienkowska-Szewczyk, K.; Mettenleiter, T.C.;Rijsewijk; Tampé, R.; Neefjes, J.; Wiertz, E.J.H J. Varicelloviruses avoid T cell recognition by UL49.5-mediated inactivation of the transporter associated with antigen processing. Proc. Nat. Acad. Sci. 2005, 102, 5144–5149. [Google Scholar] [CrossRef]
- Kouzarides, T. Acetylation: a regulatory modification to rival phosphorylation? EMBO J. 2000, 19, 1176–1179. [Google Scholar] [CrossRef] [PubMed]
- Kouzarides, T. Histone acetylases and deacetylases in cell proliferation. Curr. Opin. Genet. Dev. 1999, 9, 40–48. [Google Scholar] [CrossRef]
- Kutish, G.; Mainprize, T.; Rock, D. Characterization of the latency-related transcriptionally active region of the bovine herpesvirus 1 genome. J. Virol. 1990, 64, 5730–5737. [Google Scholar] [PubMed]
- Lees-Miller, S.P.; Long, M.C.; Kilvert, M.A.; Lam, V.; Rice, S.A.; Spencer, C.A. Attenuation of DNA-dependent protein kinase activity and its catalytic subunit by the herpes simplex virus type 1 transactivator ICP0. J. Virol. 1996, 70, 7471–7477. [Google Scholar] [PubMed]
- Levy, D.E.; Darnell, Jr., J.E. Stats: transcriptional control and biological impact. Nature Reviews Molecular Cell Biology 2002, 3, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Chow, B.; Raggo, C.; Babiuk, L.A. Bovine herpesvirus 1 UL49.5 homolog gene encodes a novel viral envelope protein that forms a disulfide-linked complex with a second virion structural. J. Virol. 1996, 70, 1448–1454. [Google Scholar] [PubMed]
- Liang, X.; Tang, M.; Manns, B.; Babiuk, L.A.; Zamb, T.J. Identification and deletion mutagenesis of the bovine herpesvirus 1 dUTPase gene and a gene homologous to herpes simplex virus UL49.5. Virology 1993, 195, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Noyce, R.S.; Collins, S.E.; Everett, R.D.; Mossman, K.L. The herpes simplex virus ICP0 RING finger domain inhibits IRF3- and IRF7-mediated activation of interferon-stimulated genes. J. Virol. 2004, 78, 1675–1684. [Google Scholar] [CrossRef] [PubMed]
- Lipińska, A.D.; Koppers-Lalic, D.; Rychlowski, M.; Admiraal, P.; Rijsewijk, F.A.M.; Bieńkowska-Szewczyk, K.; Wiertz, E.J. Bovine herpesvirus 1 UL49.5 protein inhibits the transporter associated with antigen processing despite complex formation with glycoprotein M. J. Virol. 2006, 81, 5822–5832. [Google Scholar] [CrossRef]
- Lirong, D.; Wenato, Q.; Qimin, C.; Chen, W.; Yunqi, G. bICP0 and its RING domain act as ubiquitin E3 ligases in vitro. Chinese Science Bulletin 2005, 50, 636–640. [Google Scholar] [CrossRef]
- Liu, S.F.; Brum, M.C.S.; Doster, A.; Jones, C.; Chowdhury, S.I. A bovine herpesvirus type 1 mutant virus specifying a carboxyl-terminal truncation of glycoprotein E is defective in anterograde neuronal transport in rabbits and calves. J. Virol. 2008, 82, 7432–7442. [Google Scholar] [CrossRef] [PubMed]
- Lium, E.K. Mutational analysis of the herpes simplex virus type 1 ICP0 C3HC4 zinc ring finger reveals a requirement for ICP0 in the expression of the essential alpha27 gene. J. Virol. 1997, 71, 8602–8614. [Google Scholar] [PubMed]
- Lomonte, P.; Thomas, J..; Pascale, T.; Caron, C.; Khochbin, S.; Epstein, A.L. Interaction ebtween class II histone deacetylases and ICP0 of herpes simplex virus type 1. J. Virol. 2004, 78, 6744–6757. [Google Scholar] [CrossRef] [PubMed]
- Lovato, L.; Inman, M.; Henderson, G.; Doster, A.; Jones, C. Infection of cattle with a bovine herpesvirus 1 (BHV-1) strain that contains a mutation in the latency related gene leads to increased apoptosis in trigeminal ganglia during the transition from acute infection to latency. J. Virol. 2003, 77, 4848–4857. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, P.; Welander, P.; Han, X.; Cantin, E. Herpes simplex virus type 1 DNA is immunostimulatory in vitro and in vivo. J. Virol. 2003, 77, 11158–11169. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.X.; Postigo, A.A.; Dean, D.C. Rb interacts with histone deacetylase to repress transcription. Cell 1998, 92, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Magnaghi-Jaulin, L.; Groisman, R.; Naguibneva, I.; Robin, P.; Lorain, S.; Le Villain, J.P.; Troalen, F.; Trouche, D.; Harel-Bellan, A. Retinoblastoma protein represses transcription by recruiting a histone deacetylase [see comments]. Nature 1998, 391, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Marie, I.; Smith, E.; Prakash, A.; Levy, D.E. Phosphorylation-induced dimerization of interferon regulatory factor 7 unmasks DNA binding and a bipartite transactivation domain. Molec. Cell. Biol. 2000, 20, 8803–8814. [Google Scholar] [CrossRef]
- Marie, I.; Durbin, J.E.; Levy, D.E. Differential viral induction of distinct interferon-alpha genes by positive feedback through interferon regulatory factor-7. EMBO J. 1998, 17, 6660–6669. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Balbas, M.A.; Bauer, U.M.; Nielsen, S.J.; Brehm, A.; Kouzarides, T. Regulation of E2F1 activity by acetylation. EMBO J. 2000, 19, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Maul, G.G.; Everett, R.D. The nuclear location of PML, a cellular member of the C3HC4 zinc- binding domain protein family, is rearranged during herpes simplex virus infection by the C3HC4 viral protein ICP0. J. Gen. Virol. 1994, 75, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Maul, G.G.; Guldner, H.H.; Spivack, J.G. Modification of discrete nuclear domains induced by herpes simplex virus type 1 immediate early gene 1 product (ICP0). J. Gen. Virol. 1993, 74, 2679–2690. [Google Scholar] [CrossRef] [PubMed]
- Megyer, K.; Au, W.-C.; Rosztoczy, I.; Babu, N.; Raj, K.; Miller, R.L.; Tomai, M.A.; Pitha, P.M. Stimulation of interferon and cytokine gene expression by imiquimod and stimulation by Sendai virus utilize similar signal transduction pathways. Molec. Cell. Biol. 1995, 15, 2207–2218. [Google Scholar] [PubMed]
- Meyer, F.; Perez, S.; Jiang, Y.; Zhou, Y.; Henderson, G.; Jones, C. Identification of a novel protein encoded by the latency-related gene of bovine herpesvirus 1. J. Neurovirol. 2007, 13, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Meyer, F.; Perez, S.; Geiser, V.; Sintek, M.; Inman, M.; Jones, C. A protein encoded by the bovine herpes virus 1 (BHV-1) latency related gene interacts with specific cellular regulatory proteins, including the CCAAT enhancer binding protein alpha (C/EBP-a). J. Virol. 2007, 81, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Miska, E.A.; Karlsson, C.; Langley, E.; Nielsen, S.J.; Pines, J.; Kouzarides, T. HDAC4 deacetylase associates with and represses the MEF2 transcription factor. EMBO J. 1999, 18, 5099–5107. [Google Scholar] [CrossRef] [PubMed]
- Mossman, K.L.; Saffran, H.A.; Smiley, J.R. Herpes simplex virus ICP0 mutants are hypersensitive to interferon. J. Virol. 2000, 74, 2052–2056. [Google Scholar] [CrossRef] [PubMed]
- Mossman, K.L.; Macgregor, P.F.; Rozmus, J.J.; Edwards, A.M.; Smiley, J.R. Herpes simplex virus triggers and then disarms a host antiviral response. J. Virol. 2001, 75, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Mossman, K.L.; Smiley, J.R. Herpes simplex virus ICP0 and ICP34.5 counteract distinct interferon- induced barriers to virus replication. J. Virol. 2002, 76, 1995–1998. [Google Scholar] [CrossRef] [PubMed]
- Munshi, N.; Merika, M.; Yie, J.; Senger, K.; Chen, G.; Thanos, D. Acetylation of HMG I(Y) by CBP turns off IFN beta expression by disrupting the enhanceosome. Mol. Cell 1998, 2, 457–467. [Google Scholar] [CrossRef]
- Nataraj, C.; Eidmann, S.; Hariharan, M.J.; Sur, J.H.; Perry, G.A.; Srikumaran, S. Bovine herpesvirus 1 downregulates the expression of bovine MHC class I molecules. Viral Immunol. 1997, 10, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Noyce, R.S.; Collins, S.E.; Mossman, K.L. Identification of a novel pathway essential for the immediate-early, interferon-independent antiviral repsonse to enveloped virions. J. Virol. 2006, 80, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, J.; Everett, R.D. Alphaherpesvirus proteins related to herpes simplex virus type 1 ICP0 affect cellular structures and proteins. J. Virol. 2000, 74, 10006–10017. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, J.; Lees-Miller, S.P.; Everett, R.D. Herpes simplex virus type 1 immediate-early protein vmw110 induces the proteasome-dependent degradation of the catalytic subunit of DNA- dependent protein kinase. J. Virol. 1999, 73, 650–657. [Google Scholar] [PubMed]
- Perez, S.; Meyer, F.; Saira, K.; Doster, A.; Jones, C. Premature expression of the latency-related RNA encoded by bovine herpesvirus 1 correlates with higher levels of beta interferon RNA expression in productively infected cells. J. Gen. Virol. 2008, 89, 1338–1345. [Google Scholar] [CrossRef] [PubMed]
- Perez, S.; Lovato, L.; Zhou, J.; Doster, A.; Jones, C. Comparison of inflammatory infiltrates in trigeminal ganglia of cattle infected with wild type BHV-1 versus a virus strain containing a mutaition in the LR (latency-related) gene. J. Neurovirol. 2006, 12, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Perez, S.; Inman, M.; Doster, A.; Jones, C. Latency-related gene encoded by bovine herpesvirus 1 promotes virus growth and reactivation from latency in tonsils of infected calves. J. Clin. Micro. 2005, 43, 393–401. [Google Scholar] [CrossRef]
- Perkins, N.D. Integrating cell-signalling pathways with NF-kB and IKK functions. Nature Reviews Molecular Cell Biology 2007, 8, 49–62. [Google Scholar] [CrossRef]
- Peters, G.A.; Khoo, D.; Mohr, I.; Sen, G.S. Inhibition of PACT-mediated activation of PKR by the herpes simplex simplex virus type 1 Us11 protein. J. Virol. 2002, 75, 11054–11064. [Google Scholar] [CrossRef]
- Poon, A.P.; Gu, H.; Roizman, B. ICP0 and the Us3 protein kinase of herpes simplex virus 1 independently block histone deactylation to enable gene expression. Proc. Nat. Acad. Sci. 2006, 103, 9993–9998. [Google Scholar] [CrossRef]
- Powell,J. Bovine Respiratory Disease, University of Arkansas Division of Agriculture Cooperative Extension Service, 2005, FSA:3082.
- Preston, C.M.; Harman, A.N.; Nicholl, M.J. Activation of interferon response factor-3 in human cells infected with herpes simplex virus type 1 or human cytomegalovirus. J. Virol. 2001, 75, 8909–8916. [Google Scholar] [CrossRef] [PubMed]
- Rock, D.; Lokensgard, J.; Lewis, T.; Kutish, G. Characterization of dexamethasone-induced reactivation of latent bovine herpesvirus 1. J. Virol. 1992, 66, 2484–2490. [Google Scholar] [PubMed]
- Rock, D.L.; Beam, S.L.; Mayfield, J.E. Mapping bovine herpesvirus type 1 latency-related RNA in trigeminal ganglia of latently infected rabbits. J. Virol. 1987, 61, 3827–3831. [Google Scholar] [PubMed]
- Saira, K. Functional analysis of the bovine herpesvirus-1 gene encoding bICP0, a promiscuous trans-activator, that stimulates productive infection and interferon signaling pathways. In PhD Thesis; University of Nebraska,: Lincoln, 2008. [Google Scholar] [CrossRef] [PubMed]
- Saira, K.; Zhou, Y.; Jones, C. The infected cell protein 0 encoded by bovine herpesvirus 1 (bICP0) induces degradation of interferon response factor 3 (IRF3), and consequently inhibits beta interferon promoter activity. J. Virol. 2007, 81, 3077–3086. [Google Scholar] [CrossRef] [PubMed]
- Saira, K.; Jones, C. The infected cell protein 0 encoded by bovine herpesvirus 1 (bICP0) associates with interferon regulatory factor 7 (IRF7), and consequently inhibits beta interferon promoter activity. J. Virol. 2009, 83, 3977–3981. [Google Scholar] [CrossRef] [PubMed]
- Saira, S.; Chowdhury, S.; Gaudreault, N.; Henderson, G.; Doster, A.; Jones, C. The zinc RING finger of the bovine herpesvirus 1 encoded bICP0 protein is crucial for viral replication and virulence. J. Virol. 2008, 82, 12060–12068. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Tanaka, N.; Hata, N.; Oda, E.; Taniguchi, T. Involvement of the IRF family transcription factor IRF-3 in virus-induced activation of the IFN-beta gene. FEBS Lett. 1998, 425, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Taniguchi, T.; Tanaka, N. The interferon system and interferon regulatory factor transcription factors-studies from gene knockout mice. Cytokine Growth Factor Rev. 2001, 12, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Schang, L.M. The latency-related gene of bovine herpesvirus 1 encodes a product which inhibits cell cycle progression. J. Virol. 1996, 70, 3807–3814. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; tenOever, B.R.; Grandvaux, N.; Zhou, G.-P.; Lin, R.; Hiscott, J. Triggering the interferon antiviral response through and IKK-related pathway. Science, 2003, 300, 1148–1151. [Google Scholar] [CrossRef] [PubMed]
- Sheffy, B.E.; Davies, D.H. Reactivation of a bovine herpesvirus after corticosteroid treatment. Proc. Soc. Exp. Biol. Med. 1972, 140, 974–976. [Google Scholar] [PubMed]
- Sterner, R.; Vidali, G.; Allfrey, V.G. Studies of acetylation and deacetylation in high mobility group proteins. Identification of the sites of acetylation in HMG-1. J. Biol. Chem. 1979, 254, 11577–83. [Google Scholar] [PubMed]
- Tikoo, S.K.; Campos, M.; Babiuk, L.A. Bovine herpesvirus 1 (BHV-1): biology, pathogenesis, and control. Adv. Virus Res. 1995, 45, 191–223. [Google Scholar] [PubMed]
- USDA Health Management and Biosecurity in U.S. Feedlots, 1999.
- Valarcher, J.-F.; Furze, J.; Wyld, S.; Cook, R.; Conzelman, K.-K.; Taylor, G. Role of alpha/beta interferons in the attenuation and immunogenecity of recombinant bovine respiratory syncitial viruses lacking NS proteins. J. Virol. 2003, 77, 8426–8439. [Google Scholar] [CrossRef] [PubMed]
- van Drunen Littel-van den Hurk, S.; Myers, D.; Doig, P.A.; Karvonen, B.; Habermehl, M.; Babiuk, L.A.; Jelinski, M.; Van Donkersgoed, J.; Schlesinger, K.; Rinehart, C. Identification of a mutant bovine herpesvirus-1 (BHV-1) in post-arrival outbreaks of IBR in feedlot calves and protection with conventional vaccination. Can. J. Vet. Res. 2001, 65, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Van Sant, C.; Hagglund, R.; Lopez, P.; Roizman, B. The infected cell protein 0 of herpes simplex virus 1 dynamically interacts with proteasomes, binds and activates the cdc34 E2 ubiquitin- conjugating enzyme, and possesses in vitro E3 ubiquitin ligase activity. Proc. Natl. Acad. Sci. U.S.A. 2001, 98, 8815–8820. [Google Scholar] [CrossRef] [PubMed]
- Verdel, A.; Khochbin, S. Identification of a new family of higher eukaryotic histone deacetylases. Coordinate expression of differentiation-dependent chromatin modifiers. J. Biol. Chem. 1999, 274, 2440–2445. [Google Scholar] [CrossRef] [PubMed]
- Wathelet, M.G.; Lin, C.H.; Parekh, B.S.; Ronco, L.V.; Howley, P.M.; Maniatis, T. Virus infection induces the assembly of coordinately activated transcription factors on the IFN-b enhancer in vivo. Mol. Cell 1998, 1, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Weaver, B.K.; Kumar, K.P.; Reich, N.C. Interferon regulatory factor 3 and CREB-binding protein/p300 are subunits of double-stranded RNA-activated transcription factor DRAF1. Molec. Cell. Biol. 1998, 18, 1359–1368. [Google Scholar] [PubMed]
- Wilson, V.; Jeffreys, A.J.; Barrie, P.A. A comparison of vertebrate interferon gene families deteced by hybridization with human interferon DNA. J. Mol. Biol. 1983, 166, 457–475. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.T.; Doster, A.; Jones, C. Bovine herpesvirus 1 can infect CD4(+) T lymphocytes and induce programmed cell death during acute infection of cattle. J. Virol. 1999, 73, 8657–8668. [Google Scholar] [PubMed]
- Winkler, M.T.; Doster, A.; Jones C. Persistence and reactivation of bovine herpesvirus 1 in the tonsil of latently infected calves. J .Virol. 2000, 74, 5337–5346. [Google Scholar] [CrossRef] [PubMed]
- Wirth, U.V.; Fraefel, C.; Vogt, B.; Vlcek, V.; Paces, C.; Schwyzer, M. Immediate-early RNA 2. 9 and early RNA 2.6 of bovine herpesvirus 1 are 3' coterminal and encode a putative zinc finger transactivator protein . J. Virol. 1992, 66, 2763–2772. [Google Scholar] [PubMed]
- Wirth, U.V.; Gunkel, K.; Engels, M.; Schwyzer, M. Spatial and temporal distribution of bovine herpesvirus 1 transcripts. J. Virol. 1989, 63, 4882–4889. [Google Scholar] [PubMed]
- Wirth, U.V.; Vogt, B.; Schwyzer, M. The three major immediate-early transcripts of bovine herpesvirus 1 arise from two divergent and spliced transcription units. J. Virol. 1991, 65, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Workman, A.; Perez, S.; Meyer, F.; Doster, A.; Jones, C. Dexamethasone treatment of calves latently infected with bovine herpesvirus 1 (BHV-1) leads to activation of the bICP0 early promoter. J Virol. 2009, In Press. [Google Scholar] [PubMed]
- Yang, H.; Ma, G.; Lin, C.H.; Orr, M.; Wathelet, M.G. Mechanism for transcriptional synergy between interferon regulatory factor (IRF)-3 and IRF-7 in activation of the interferon-beta gene promoter. Eur. J. Biochem. 2004, 271, 3693–3703. [Google Scholar] [CrossRef] [PubMed]
- Yates, W.D.G. A review of infectious bovine rhinotracheitis, shipping fever pneumonia, and viral-bacterial synergism in respiratory diesease of cattle. Can. J. Comp. Med. 1982, 46, 225–263. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Suhara, W.; Fukuhara, M.; Fukuda, M.; Nishida, E.; Fujita, T. Direct triggering of the type 1 interferon system by virus infection: activation of a transcription factors containing IRF-3 and CBP/p300. EMBO J. 1998, 17, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Yuang, Y.; Lowther, W.; Kellum, M.; Au, W.C.; Lin, R.; Hiscott, J.; Pitha, P.M. Primary activation of interferon A and interferon B gene transcription by interferon regulatory factor 3. Proc. Natl. Acad. Sci. USA 1998, 95, 9837–9842. [Google Scholar] [CrossRef]
- Zhang, L.; Pagano, J.S. Interferon regulatory factor 7 is induced by Epstein-Barr virus latent membrane protein 1. J Virol. 2000, 74, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Bieker, J.J. Acetylation and modulation of erythroid Kruppel-like factor (EKLF) activity by interaction with histone acetyltransferases. Proc. Natl. Acad. Sci. USA 1998, 95, 9855–9860. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jones, C. Identification of functional domains within the bICP0 protein encoded by BHV-1. J. Gen. Virol. 2005, 86, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jones, C. The bovine herpesvirus 1 immediate-early protein (bICP0) associates with histone deacetylase 1 to activate transcription. J. Virol. 2001, 75, 9571–9578. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jiang, Y.; Zhou, J.; Jones, C. The bovine herpes virus 1 (BHV-1) immediate early protein (bICP0) interacts with the histone acetyltransferase p300, and these interactions correlate with stimulation of gC promoter activity. J. Gen. Virol. 2006, 87, 1843–1851. [Google Scholar] [CrossRef] [PubMed]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
Jones, C. Regulation of Innate Immune Responses by Bovine Herpesvirus 1 and Infected Cell Protein 0 (bICP0). Viruses 2009, 1, 255-275. https://doi.org/10.3390/v1020255
Jones C. Regulation of Innate Immune Responses by Bovine Herpesvirus 1 and Infected Cell Protein 0 (bICP0). Viruses. 2009; 1(2):255-275. https://doi.org/10.3390/v1020255
Chicago/Turabian StyleJones, Clinton. 2009. "Regulation of Innate Immune Responses by Bovine Herpesvirus 1 and Infected Cell Protein 0 (bICP0)" Viruses 1, no. 2: 255-275. https://doi.org/10.3390/v1020255
APA StyleJones, C. (2009). Regulation of Innate Immune Responses by Bovine Herpesvirus 1 and Infected Cell Protein 0 (bICP0). Viruses, 1(2), 255-275. https://doi.org/10.3390/v1020255