1. Introduction
Infectious bronchitis virus (IBV) is a pathogenic gamma coronavirus that causes respiratory symptoms, egg drops, nephritis and proventriculitis in domestic fowls [
1]. Due to the persistent infection IBVs are difficult to eradicate and easy to mutate into new recombinant viruses in infected flocks [
2,
3]. The single-stranded, positive-sense RNA genome is approximately 27 kb and encodes RNA polymerase/transcriptase (1ab), spike (S), envelope (E), membrane (M), nonstructural (3a, 3b, 5a, 5b), and nucleocapsid (N) proteins [
1]. Approximately 15 nonstructural proteins (nsp) are generated from the large 1a and 1ab proteins by viral proteases, and these nsp play roles in virus replication and pathogenicity [
4,
5]. The spike protein is a protective antigen that is essential for virus infection in host cells [
1].
In Korea, several genotypes of IBV, including K-I, KM91-like, New cluster 1 (NC1), QX-like and recurrent QX-like, have been reported [
6,
7,
8]. Among them, nephropathogenic KM91-like viruses were first reported in 1991 and became prevalent in the field. An inactivated oil emulsion vaccine and an embryo-adapted, attenuated live vaccine (K2 strain) have been used to prevent KM91-like virus infection, but occasionally, KM91-like viruses have been isolated in the field [
8].
Cold adaptation has been used for the attenuation of various viruses, and a cold-adapted influenza A virus (IAV) has been successfully applied to the generation of attenuated human vaccine strains [
9,
10]. Cold-adapted IBV strains have also been established and characterized; however, the biological traits and genetic backgrounds of these cold-adapted IBVs have not been completely elucidated [
9,
10,
11,
12]. IBVs replicate firstly in the trachea then disseminate into internal organs. The temperature of the trachea is lower than internal organs, and it may be valuable to characterize the traits of cold-adapted IBVs. Furthermore, in comparison with respiratory and kidney pathogenicity tests, reproductive organ pathogenicity tests have not been easy to perform due to the long periods of observation necessary for the sexual maturation of hens [
6]. Therefore, a time-saving and reproducible animal model to test reproductive organ pathogenicity is required [
13].
In this study, we established a cold-adapted IBV (BP-caKII), and we characterized its pathogenicity in embryos and chickens, tissue tropism, and persistence of infection. We applied a premature reproductive tract pathogenicity model to the differentiation of the pathogenicity of IBVs and performed comparative genomics to shed light on the genetic background of the embryo adaptation of K2 and the cold adaptation of BP-caKII.
2. Materials and Methods
2.1. Virus, Egg and Chicken
SNU9106 was isolated from pooled tissues of the cecal tonsil and trachea of commercial layers (104-day-old) and was sent to the Laboratory of Avian Diseases of Seoul National University for diagnosis in 2009. A live attenuated commercial vaccine strain, K2 was previously established by 172 times passages of embryonated eggs [
8]. A nephropathogenic field strain, KM91 that had been passaged 5 times through embryonated eggs was kindly provided by Avian Disease Division, animal and Plant Quarantine Agency in Korea. The viruses were propagated by inoculating into 10-day-old SPF embryonated eggs (Valo Biomedia, Adel, IA, USA) via the allantoic cavity route and incubated for 48 to 72 h, after which they were chilled at 4 °C overnight. The allantoic fluid was harvested, and the supernatant was stored at −70 °C after centrifugation at 3000 rpm (1915×
g) for 10 min.
2.2. Primers, RNA Extraction, Real-Time and Conventional RT-PCR, and Genome Analysis
A primer set for real-time RT-PCR was designed based on the conserved region of the
nsp 3 genes of 14 reference strains in the GenBank (
Table 1). The reference strains used for the primer design are as follows: LX, Peafowl/GD/KQ6, KM91, QIA-03342, QIA-KR/D79/05, QIA-Q43, SNU-9106, SNU-10043, M41, Turkey coronavirus, SNU-8067, ITA/90254/2005, and ArkDPI11. Their accession numbers are available in Figure 3. Primers for genome amplification and sequencing were described previously and additional new primers were summarized in
Table 1 [
6].
Viral genomic RNA was extracted from 100 µL of infectious allantoic fluid using the Viral Gene-Spin Kit (iNtRON Biotechnology, Seongnam, Korea). SYBR real-time one-step RT-PCR (real-time RT-PCR) was performed using an Applied Biosystems StepOne real-time PCR machine and a one-step RT-PCR kit (KAPA-Biosystems, Boston, MA, USA) according to the manufacturer’s instructions. Total reaction volumes were adjusted to 10 µL and 1 µL of the extracted RNA used.
Conventional RT-PCR for genome amplification was performed using a one-step RT-PCR kit (Qiagen GmbH, Hilden, Germany) according to the manufacturer’s instructions. The RT-PCR conditions were as follows: cDNA synthesis at 50 °C for 30 min, inactivation of the reverse transcriptase at 95 °C for 15 min, 40 cycles of denaturation at 94 °C for 30 s, annealing at 50 °C for 30 s, and extension at 72 °C for 2 min, with a final extension at 72 °C for 5 min. The amplicons were purified using a PCR purification kit (MEGA-quick-spin Total Fragment DNA Purification Kit, iNtRON Biotechnology) and sequenced with PCR and sequencing primers using an ABI3711 automatic sequencer (Macrogen Co., Seoul, Korea). The overlapping gene fragments were assembled to obtain a single complete genome sequence using ChromasPro version 1.5 (Technelysium Pty Ltd., Brisbane, Australia). Nucleotide (nt) and amino acid (a.a) identity estimates and amino acid translations were obtained using BioEdit (ver. 5.0.9.1.). The full genome sequences of BP-caKII and K2 were submitted to the GenBank database. Phylogenetic analyses of the complete
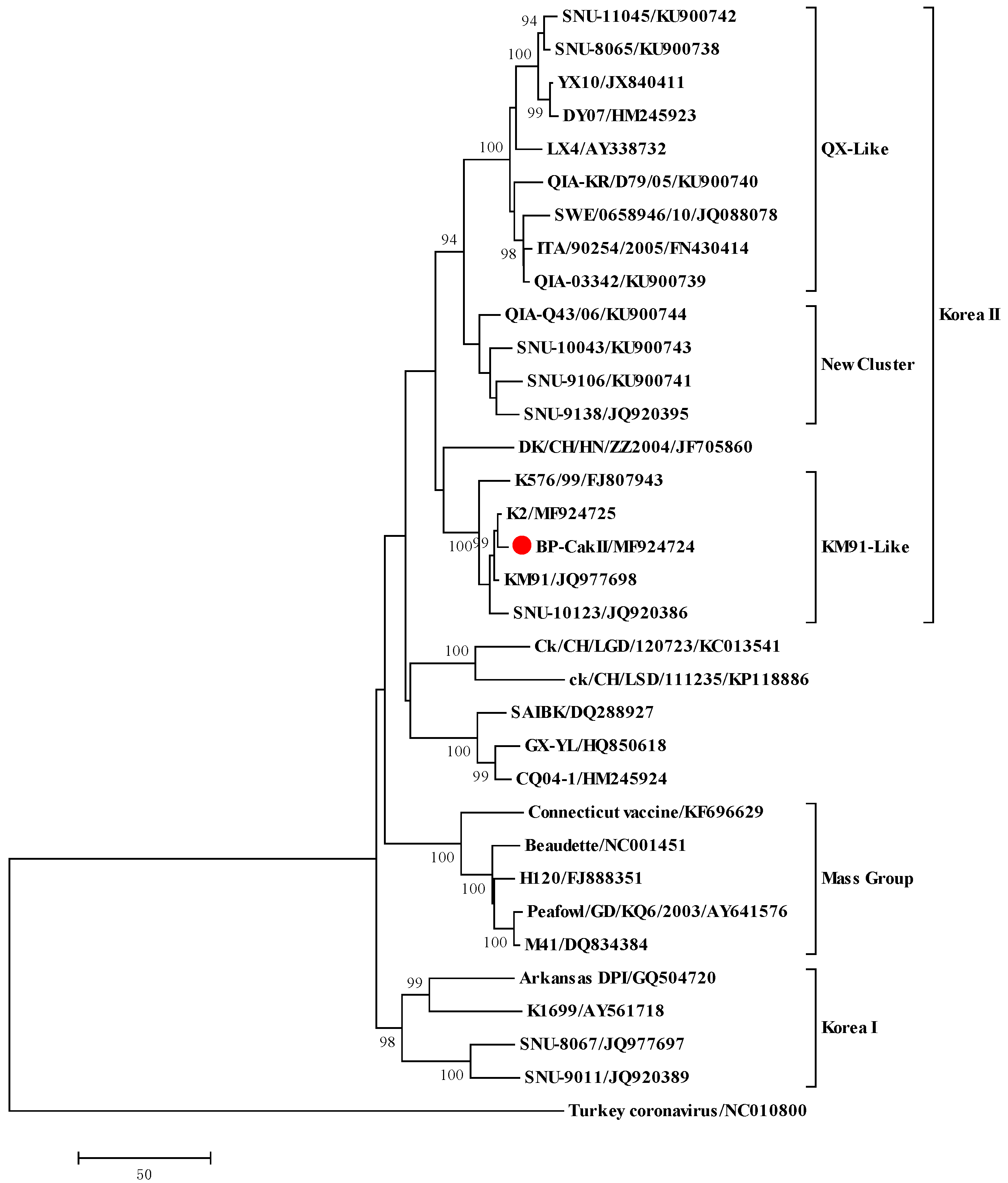
S1 gene of the IBV strains were conducted using MEGA software (ver. 5.0.5, neighbor-joining method with Tamura-Nei distance and 1000 repeats of bootstrapping) [
14].
2.3. Cold Adaptation of the Virus
SNU9106, propagated via three blind passages, was diluted by 10−3 and inoculated into 10-day-old SPF embryonated eggs (Valo Biomedia, Adel, IA, USA) via the allantoic cavity route; the embryonated eggs were incubated at 32 °C for 48 h, after which they were chilled at 4 °C overnight. The allantoic fluid was harvested, and the supernatant was stored at −70 °C after centrifugation at 3000 rpm (1915× g) for 10 min. The allantoic fluid harvested from the inoculated embryonated eggs was used for the real-time RT-PCR. The allantoic fluid with the lowest Ct value was inoculated into the embryonated eggs, and the same procedure was repeated 20 times to become BP-caKII.
2.4. Virus Titration, Embryo Pathogenicity and Embryo Mortality Rate by Virus
To test virus titer and embryo pathogenicity, BP-caKII was inoculated into 10-day-old SPF embryonated eggs (Valo Biomedia) via the allantoic cavity route and incubated at 37 °C for 48 h. The infected allantoic fluid was harvested after chilling at 4 °C overnight. Then, BP-caKII and K2 titers were determined by inoculating 10-fold serial dilutions (10
−1–10
−7) of the virus into five 10-day-old SPF embryonated eggs via the allantoic cavity. The inoculated embryonated eggs were observed for death and dwarfism for 5 days at 37 °C, and the 50% chicken embryo infectious dose (EID
50/mL) was calculated using the Reed-Muench formula [
15]. The mortality rate of BP-caKII and K2 was measured by mean death time (MDT), and absolute lethal dose (ALD). MDT was measured as previously reported [
16,
17,
18]. Briefly, BP-caKII and K2 were diluted within the range of 10
2 to 10
4 EID
50/100 µL and 100 µL was injected into 10-day-old SPF embryonated eggs via the allantoic cavity. Five embryonated eggs per dilution were inoculated at 8 a.m., and inoculation was repeated at 4 p.m. with the same dose. The candling of embryonated eggs for mean death time was performed at eight-hour intervals within one day. ALD was determined by the highest dilution at which all embryos died.
2.5. Growth Kinetics
BP-caKII and K2 diluted to 1 × 102 EID50/100 µL were injected into 10-day-old SPF embryonated eggs (Valo BioMedia, Osterholz-Scharmbeck, Germany) via the allantoic cavity route and incubated at 32 °C and 37 °C for 60 h. After 0, 12, 18, 24, 30, 36, 48 and 60 h, three eggs inoculated with each virus were chilled at 4 °C overnight. The allantoic fluid of all the eggs was harvested and viral RNA was extracted for the real-time PCR. The Ct values of samples were measured, and the ΔCt was calculated by subtract Ct value of each sample from that of the 0 h sample. In addition, we performed regression analysis with Ct values of 2-fold diluted BP-caKII and K2 viruses and fitted linear equations of BP-caKII and K2. Theoretically 1 unit of ΔCt is the 2-fold difference of RNA copies, but we calculated the real fold difference of RNA of each virus by using the slope value of the linear equation (BP-caKII, y = 0.259x + 9.94, R2 = 0.997, 1.81-fold difference/Ct; K2, y = 0.267x + 11.1, R2 = 0.997, 1.84-fold difference/Ct). The independent experiment was repeated and each sample was tested in triplicate in an experiment.
2.6. Tissue Tropism and Persistence Test
To determine tissue tropism and the persistence of IBV strains, we performed experimental viral infection using 80 7-day-old SPF chicks that were assigned to 4 groups (n = 20 per group). All chicks were treated with PBS (negative control), BP-caKII, K2, or KM91 (1.5 × 105 EID50/100 µL/chick) via ocular and intranasal routes, and observed for clinical signs and mortality for 4 weeks after the challenge. Each week, five chicks from each group were euthanized by cervical dislocation, and the pathological lesions of internal organs were observed throughout necropsy. The tracheal, cecal tonsil and kidney tissue samples were collected and examined using the real-time RT-PCR without virus titer calculation.
2.7. Generation of a Premature Reproductive Tract Pathogenicity Model Using DES (Diethylstilbestrol)
To investigate the influence of IBV on the premature reproductive organs of the chicks, we induced the sexual precocity by DES (Sigma) treatment [
13]. A total of 48 1-day-old SPF female chicks (BioPOA Co., Yongin, Korea) were distributed into four different groups. Three groups were inoculated with BP-caKII, K2, and KM91 (10
6.5 EID
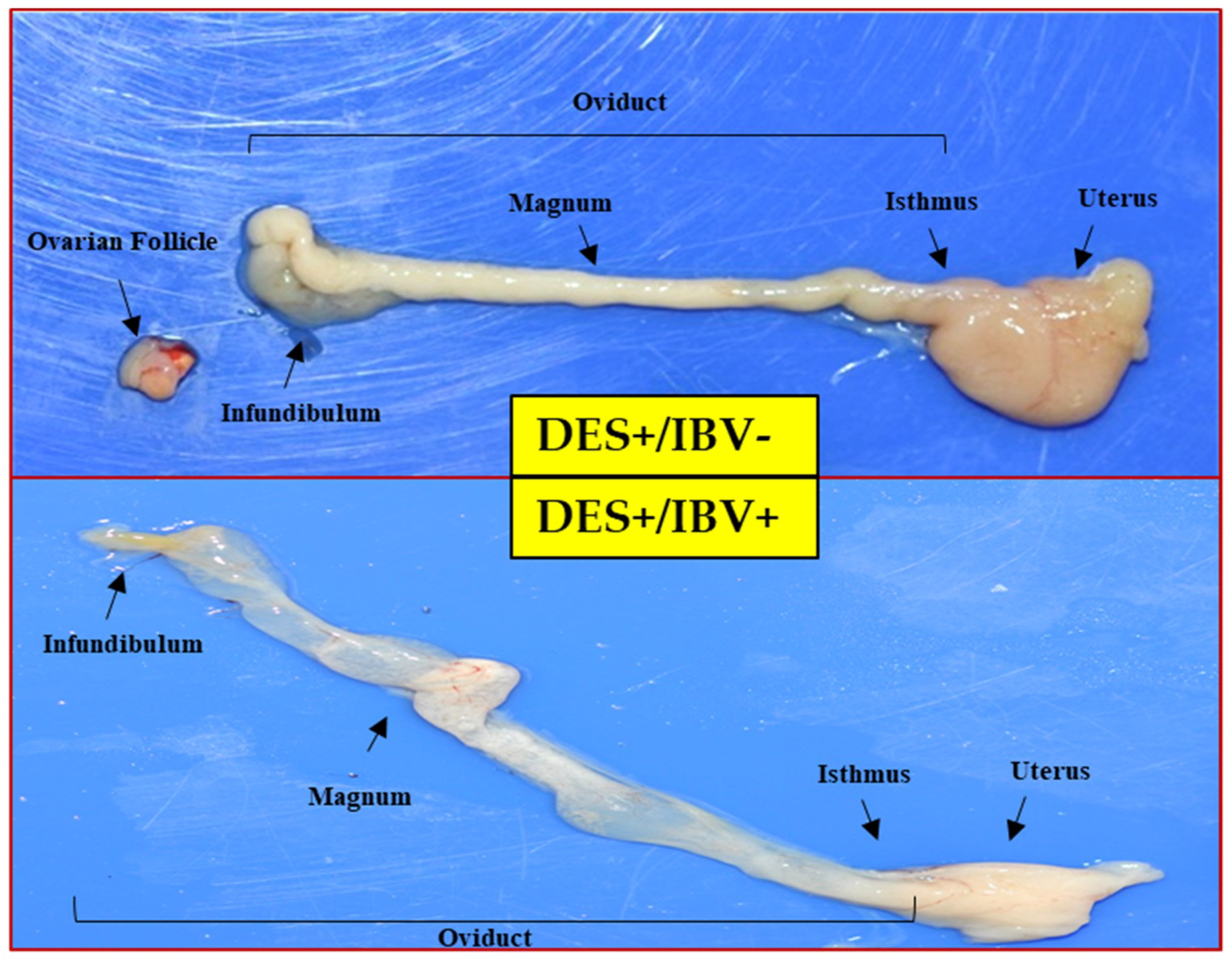
50/100 µL/chick) via ocular and intranasal routes, and a DES-control group was not inoculated with virus. A total of 12 birds were assigned to each group. After inoculation with each virus, the dead chicks were excluded when comparing reproductive tract lesions. At 5, 8 and 11 days after virus infection, all chicks were inoculated with DES intramuscularly (1 mg/100 µL) and were observed for clinical signs and mortality for 2 weeks after the challenge and were euthanized by cervical dislocation at 2 weeks post-infection. During the experiment, all chicks were reared in air-filtered isolators (THREESHINE, Daejeon, Korea) and feed and water were provided ad libitum. Gross lesions of the reproductive tract were examined based on cyst formation (four-grade-scoring: Negative, moderate, + (cyst covering less than 20% of oviduct), marked, ++ (covering 20% to 50%), and severe, +++ (covering more than 50% of oviduct)), and the presence or absence of caseous material in the oviduct.
2.8. Statistical Analysis
The significance of growth kinetics and embryo pathogenicity were evaluated by two way analysis-of-variance (ANOVA) and a Man-Whitney test, respectively. The difference in virus frequency in tissues and lesions in the premature reproductive tract pathogenicity model was assessed using chi-squared distribution (95% confidence intervals).
2.9. Ethical Statement
All animal experiments were performed at BioPOA Co. (Suwon, South Korea) following a protocol that adhered to the National Institutes of Health’s Public Health Service Policy on the Humane Care and Use of Laboratory Animals. The protocol was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of BioPOA Co. (tissue tropism and persistence test, identification code: BP-2016-003-2; date of approval: 15 January 2016; premature reproductive tract pathogenicity test, identification code: BP-2016-001-2; date of approval: 15 January 2016).
4. Discussion
Passages of pathogenic IBVs through embryonated eggs resulted in virus attenuation and efficient virus replication, and all of the attenuated IBV vaccine strains, such as H120, K2 and YX10p90, have been established by embryonic passages [
8,
21,
22]. Cold adaptation was another virus attenuation method, and a cold-adapted IAV vaccine strain has been used to generate vaccine strains against seasonal flu [
9,
10]. In a previous study, cold adaptation of IBV was reported, but the attenuation of pathogenicity was incomplete, and the genomic backgrounds of the attenuated strains were unavailable [
11]. In this study, we established a cold-adapted, attenuated IBV strain, BP-caKII, and characterized its pathobiological and genomic traits.
Although we started embryo passages with a SNU9106 isolate that had been classified into the new cluster I genotype, the E5, E10, E15 and BP-caKII strains were classified into the KM91-like genotype. SNU9106 is a recombinant virus possessing the
1ab gene of the KM91-like genotype and a recombinant
S1 gene containing a partial segment from the QX-like virus [
20]. Therefore, BP-caKII was unlikely to have originated from SNU9106. Although the K2 strain has been used commercially since 2010, it might have been under clinical evaluation in the field for governmental approval for commercial use during this period, including 2009 [
23]. The multiple shared mutations between K2 and BP-caKII, and the fact that both strains shared almost the same mutational patterns following cold-passaging (20 times), indicate that the K2 and BP-caKII strains may be closely related.
Although BP-caKII showed less pathogenicity in embryonated eggs than K2 they showed more delayed viral dissemination to the kidneys and cecal tonsils than did KM91. KM91 was detected and persisted in the trachea and kidneys for the first 3 weeks and in the cecal tonsils throughout the whole period of observation (4 weeks). Pathogenic field isolates of QX-like and variant IBVs also persisted for a long time in the trachea, proventriculus, kidneys and cecal tonsils of infected chickens [
24]. Therefore, embryo and cold adaptation apparently decreased pathogenicity and changed the tissue tropism of K2 and BP-caKII in embryonated eggs and chickens. However, in certain conditions, even vaccine strains may persist for a long time before being excreted into the trachea and cloaca [
3]. Thus, vaccine strains cleared within a short period of time, but with enough time to induce protective immunity, may be preferable to minimize the conversion of pathogenicity and the occurrence of recombinant viruses.
To date, various pathogenicity models of IBVs have been developed, but testing pathogenicity on reproductive organs requires a long period before the sexual maturation of hens occurs [
6]. DES has been known to induce abnormalities in female reproductive tracts and to induce a premature phenotype in the reproductive tracts of female chicks [
13,
25]. Previously, the premature reproductive tract pathogenicity model was established and used to evaluate IBV pathogenicity [
13]. In this study, we verified the pathogenicity of BP-caKII, K2 and KM91 by using this model. KM91 showed a significantly higher pathogenicity than BP-caKII and K2, but the pathogenicity of BP-caKII was insignificantly different from K2. Considering the differences in pathogenicity in embryos and premature reproductive tracts, as well as the differences in tissue tropism, BP-caKII may be less pathogenic than K2.
The comparative genomics study of BP-caKII and K2 revealed mutations acquired during embryo and cold adaptation. K2 acquired relatively more missense mutations in nsp13, nsp14, nsp15 and
nsp16 genes than other genes during embryo adaptation. Nsp13 is multifunctional, and its RNA 5’ triphosphatase and helicase domains are located on its N- and C-terminals, respectively [
26,
27]. The helicase domain of nsp13 is subdivided into the RecA1 and RecA2 domains, and four out of the five mutations identified were located in RecA2 [
26]. Nsp14 is composed of the N-terminal exonuclease (ExoN) and C-terminal AdoMet-dependent guanosine N7-methyltransferase (N7-MTase) domains, and all the mutations were located in both domains [
28,
29,
30]. ExoN and N7-MTase play roles in the correction of replication errors and in mRNA capping, respectively [
28,
29]. Nsp16 is an AdoMet-dependent 2’-O-methyltransferase (2’-O-MTase) that plays a role in the capping of viral RNAs, which helps in the evasion of innate immunity [
31]. Nsp15 is an endoribonuclease that plays an important role in the evasion of dsRNA sensors, such as RIG1 and MDA5, in infected cells [
32]. The mutations shared by K2 and BP-caKII in the
nsp14, 15, and
16 genes may facilitate or impair innate immunity evasion in embryos and chickens, respectively. However, the exact innate immunity differences between embryos and chickens have never been elucidated. To date, embryo-adapted strains of IBVs have shown high mutation rates in the
nsp3 genes, including a large nucleotide deletion in the 3’ end of the
N gene and partial coding region of gene 6 [
22,
33]. Therefore, mutations for embryo adaptation may be strain-specific.
BP-caKII acquired multiple additional missense mutations in the
nsp 3,
4,
12,
S and
N genes during cold adaptation, but the exact functions of these mutations were unknown. Enzyme activity may be affected at different temperatures.
Nsp3 has multiple domains, and three out of the four mutations were located in the PL (papain-like protease, L978F) and 3Ecto (NSP3-ectodomain, I1167del and V1168I) domains [
34]. Nsp12 is an RNA-dependent RNA polymerase (RdRp), and the P214L mutation and the R626C and P812I mutations are located in the Nidovirus RdRp-associated nucleotidyltransferase (NiRAN) and the RdRp domains, respectively.
Nsp4 is involved in the formation of the replication and transcription complex (RTC) with nsp3, and nsp12 is an essential component of the RTC. Therefore, further study on the roles of the multiple mutations of nsp3, 4 and 12 seen during viral replication at the lower temperature may be interesting. The R118M mutation in S1 NTD of K2 and BP-caKII is located in the partial ceiling region of the receptor binding site, and V66A of BP-caKII is located in the vicinity of the R118M mutation site [
35]. Evolutionarily, coronaviruses developed a ceiling to protect the receptor binding site in S1 NTD from host immune surveillance, and the R118M and V66A mutations may be related to the function of the partial ceiling [
35]. The accumulation of multiple mutations in the
N gene was interesting, but their functions in cold adaptation are unclear. The cold-adapted IAV showed mutations in its nucleoprotein, as well as the polymerase genes [
36]. The RNA binding domain and SR (serine and arginine-rich) region of N interact with the ubiquitin-like domain of nsp3, and K136T and I204V mutations are located in the NTD (N-terminal domain-a.a 19–162) RNA binding site (136–190) and LKR (Linker region) (a.a 163–218), respectively [
37,
38]. The V317L and T343S mutations are in the putative dimerization core (a.a 250–349) of CTD (C-terminal domain-a.a 219–349) that binds single-stranded RNA [
39]. Therefore, multigenic mutations may be acquired during cold adaptation of coronavirus.

 ,
, 



{kind=link}
{kind=link}
{kind=link}