Genetic and Phenotypic Characterization of Cryphonectria hypovirus 1 from Eurasian Georgia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
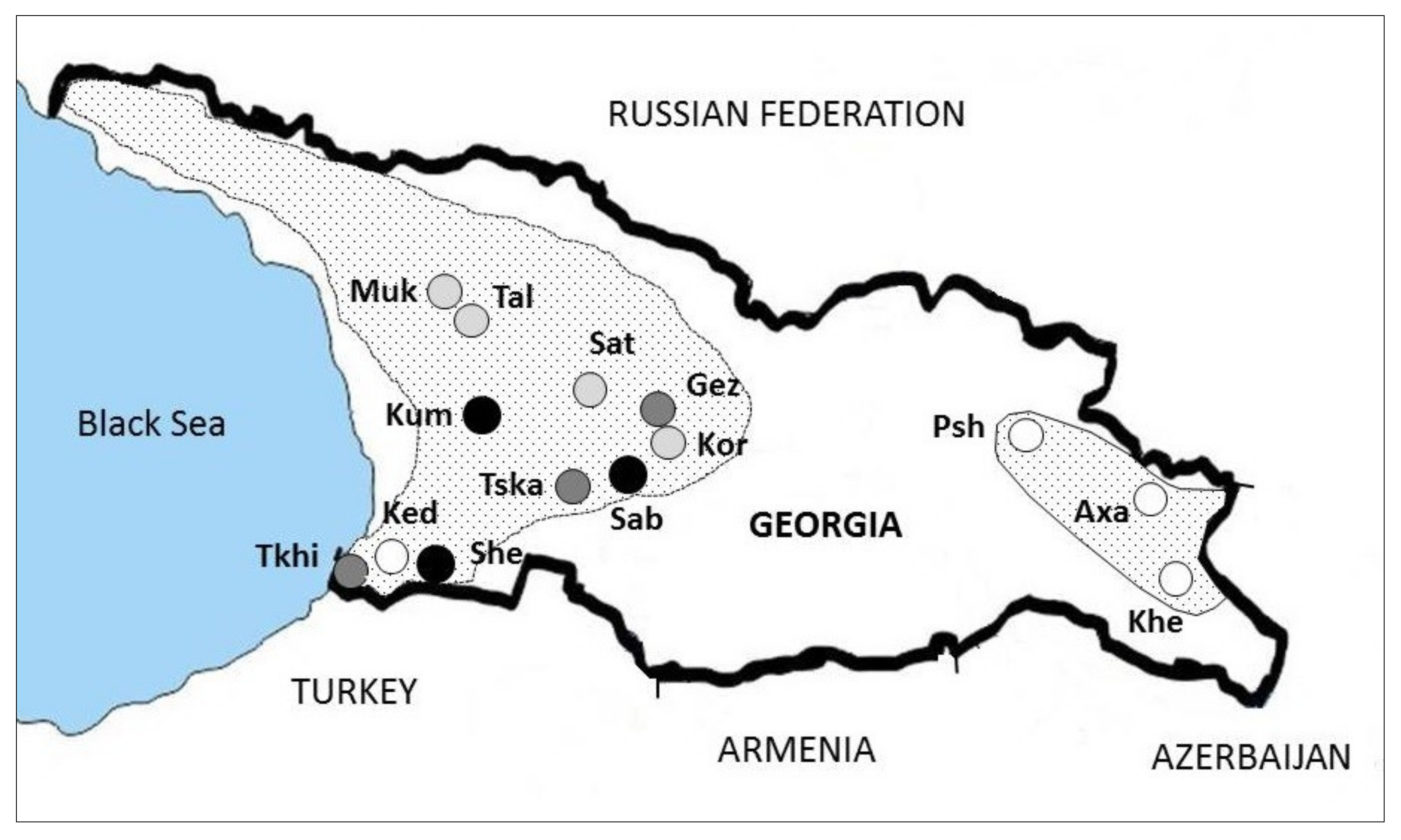
2.1. Fungal Isolates
2.2. RNA Extraction and RT-PCR
2.3. Sequencing of CHV-1
2.4. Phylogenetic Analysis of CHV-1
2.5. Phenotypic Effects of the Georgian CHV-1 Subtype on C. parasitica
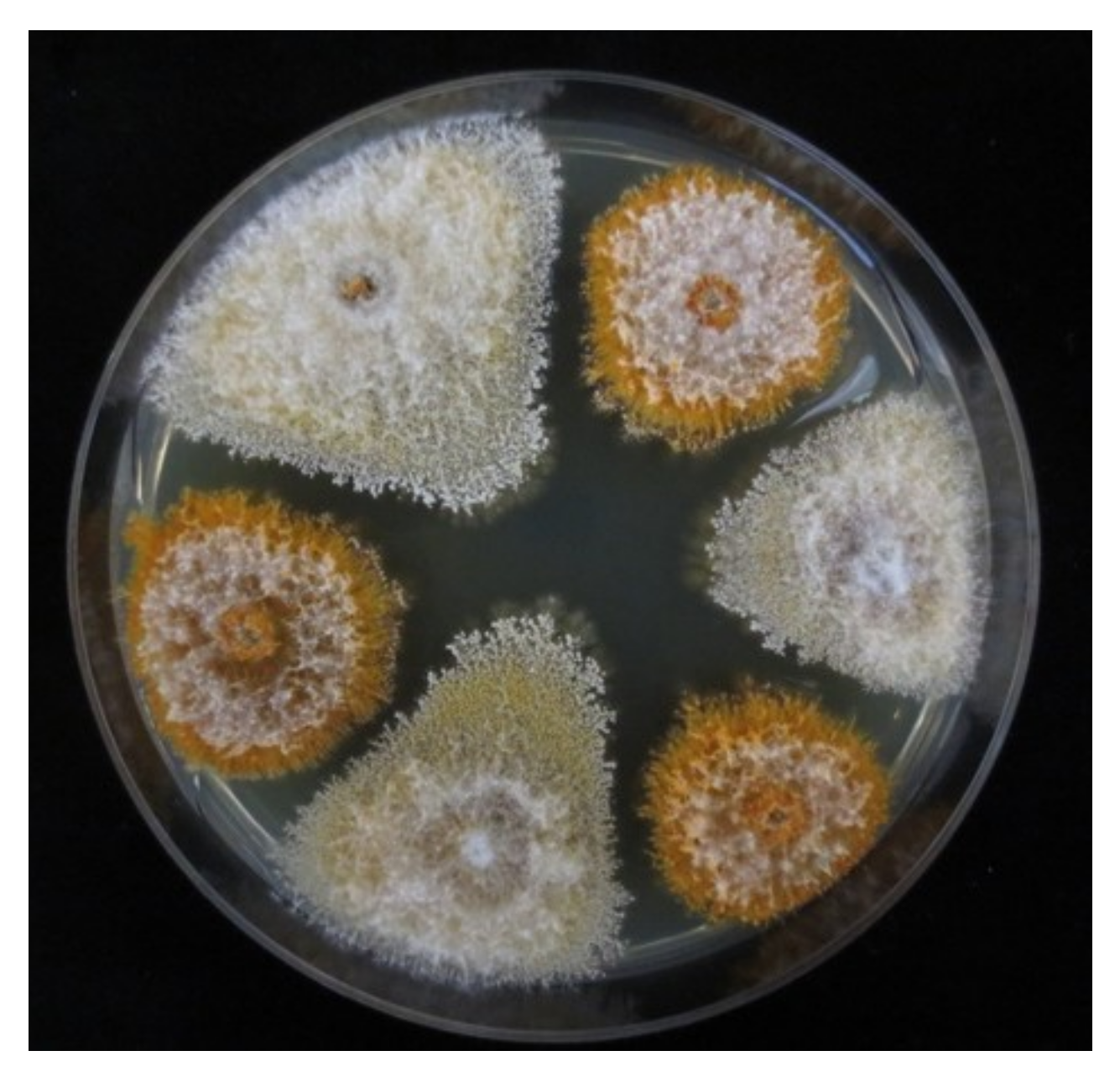
2.5.1. Growth on PDA Plates
2.5.2. Growth on Dormant Chestnut Stems
2.6. Data Analysis
3. Results
3.1. Prevalence and Genetic Characterization of CHV-1
3.2. Phylogenetic Analysis
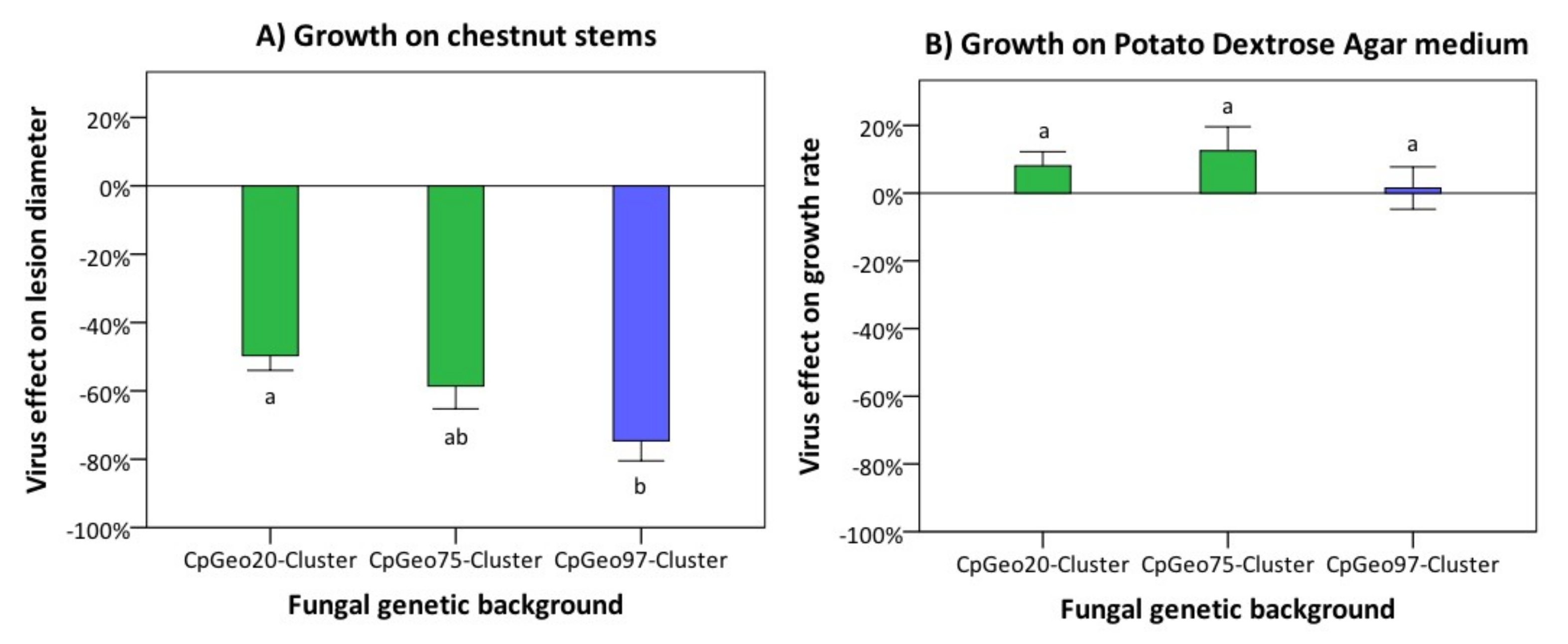
3.3. Effect of the Georgian CHV-1 Subtype on Growth of C. parasitica
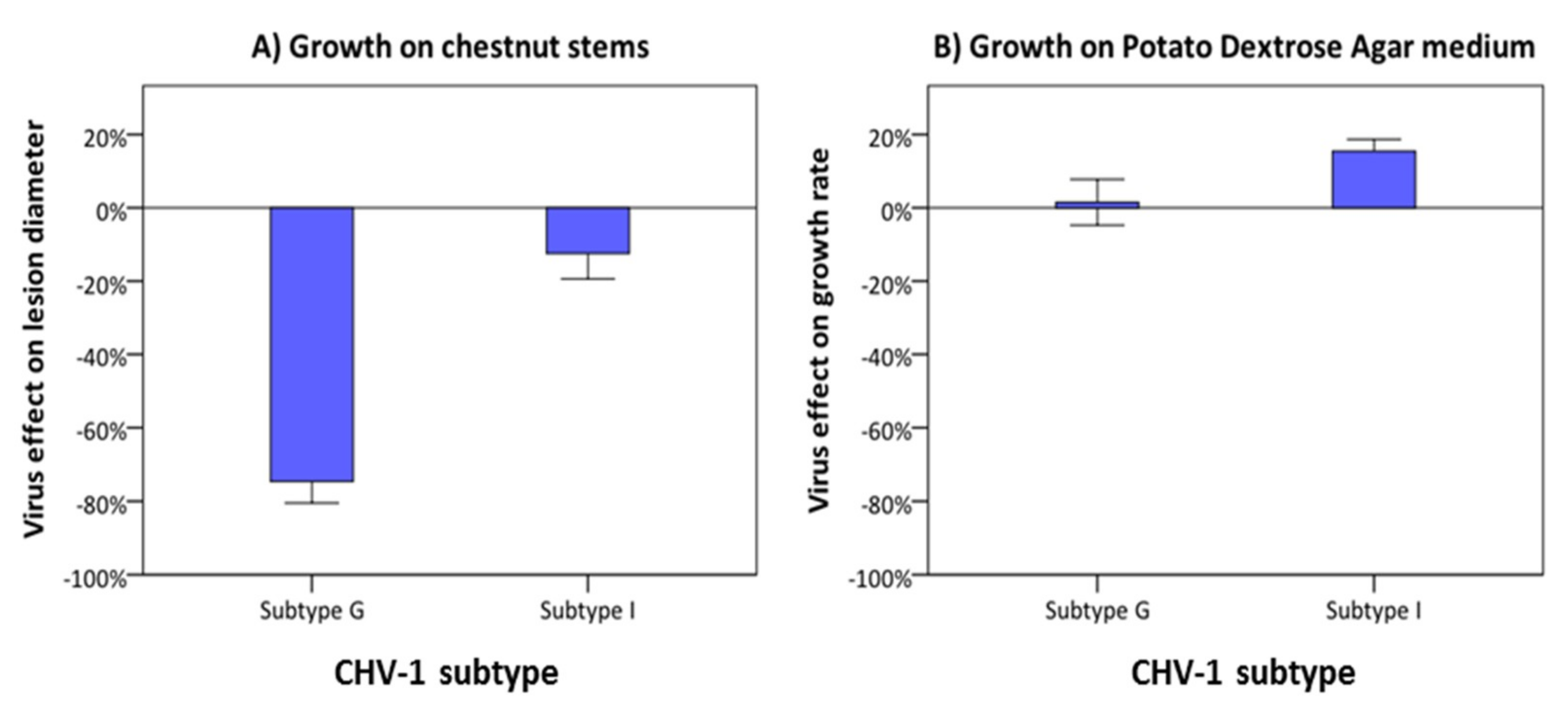
3.4. Comparison of the Georgian and Italian CHV-1 Subtypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal-plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Rigling, D.; Prospero, S. Cryphonectria parasitica, the causal agent of chestnut blight: Invasion history, population biology and disease control. Mol. Plant Pathol. 2018, 19, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Anagnostakis, S.L. Chestnut blight—The classical problem of an introduced pathogen. Mycologia 1987, 79, 23–37. [Google Scholar] [CrossRef]
- Heiniger, U.; Rigling, D. Biological control of chestnut blight in Europe. Annu. Rev. Phytopathol. 1994, 32, 581–599. [Google Scholar] [CrossRef]
- Choi, G.H.; Nuss, D.L. Hypovirulence of chestnut blight fungus conferred by an infectious viral cDNA. Science 1992, 257, 800–803. [Google Scholar] [CrossRef]
- Peever, T.L.; Liu, Y.C.; Cortesi, P.; Milgroom, M.G. Variation in tolerance and virulence in the chestnut blight fungus-hypovirus interaction. Appl. Environ. Microb. 2000, 66, 4863–4869. [Google Scholar] [CrossRef]
- Cortesi, P.; McCulloch, C.E.; Song, H.Y.; Lin, H.Q.; Milgroom, M.G. Genetic control of horizontal virus transmission in the chestnut blight fungus, Cryphonectria parasitica. Genetics 2001, 159, 107–118. [Google Scholar]
- Prospero, S.; Conedera, M.; Heiniger, U.; Rigling, D. Saprophytic activity and sporulation of Cryphonectria parasitica on dead chestnut wood in forests with naturally established hypovirulence. Phytopathology 2006, 96, 1337–1344. [Google Scholar] [CrossRef]
- Grente, M.J. Les formes hypovirulentes d’Endothia parasitica et les espoirs de lutte contre le chancre du châtaignier. Académie D’agriculture de France 1965, 51, 1033–1036. [Google Scholar]
- Diamandis, S. Management of chestnut blight in Greece using hypovirulence and silvicultural interventions. Forests 2018, 9, 492. [Google Scholar] [CrossRef]
- Bryner, S.F.; Prospero, S.; Rigling, D. Dynamics of Cryphonectria hypovirus infection in chestnut blight cankers. Phytopathology 2014, 104, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Gobbin, D.; Hoegger, P.J.; Heiniger, U.; Rigling, D. Sequence variation and evolution of Cryphonectria hypovirus 1 (CHV-1) in Europe. Virus Res. 2003, 97, 39–46. [Google Scholar] [CrossRef]
- Feau, N.; Dutech, C.; Brusini, J.; Rigling, D.; Robin, C. Multiple introductions and recombination in Cryphonectria hypovirus 1: Perspective for a sustainable biological control of chestnut blight. Evol. Appl. 2014, 7, 580–596. [Google Scholar] [CrossRef] [PubMed]
- Bryner, S.F.; Rigling, D.; Brunner, P.C. Invasion history and demographic pattern of Cryphonectria hypovirus 1 across European populations of the chestnut blight fungus. Ecol. Evol. 2012, 2, 3227–3241. [Google Scholar] [CrossRef] [PubMed]
- Robin, C.; Lanz, S.; Soutrenon, A.; Rigling, D. Dominance of natural over released biological control agents of the chestnut blight fungus Cryphonectria parasitica in South-Eastern France is associated with fitness-related traits. Biol. Control 2010, 53, 55–61. [Google Scholar] [CrossRef]
- Akilli, S.; Serce, C.U.; Katircioglu, Y.Z.; Maden, S.; Rigling, D. Characterization of hypovirulent isolates of the chestnut blight fungus, Cryphonectria parasitica from the Marmara and Black Sea regions of Turkey. Eur. J. Plant Pathol. 2013, 135, 323–334. [Google Scholar] [CrossRef]
- Peters, F.S.; Busskamp, J.; Prospero, S.; Rigling, D.; Metzler, B. Genetic diversification of the chestnut blight fungus Cryphonectria parasitica and its associated hypovirus in Germany. Fungal Biol. 2014, 118, 193–210. [Google Scholar] [CrossRef]
- Trapiello, E.; Rigling, D.; Gonzalez, A.J. Occurrence of hypovirus-infected Cryphonectria parasitica isolates in northern Spain: An encouraging situation for biological control of chestnut blight in Asturian forests. Eur. J. Plant Pathol. 2017, 149, 503–514. [Google Scholar] [CrossRef]
- Mlinarec, J.; Nuskern, L.; Jezic, M.; Rigling, D.; Curkovic-Perica, M. Molecular evolution and invasion pattern of Cryphonectria hypovirus 1 in Europe: Mutation rate, and selection pressure differ between genome domains. Virology 2018, 514, 156–164. [Google Scholar] [CrossRef]
- Bryner, S.F.; Rigling, D. Hypovirus virulence and vegetative incompatibility in populations of the chestnut blight fungus. Phytopathology 2012, 102, 1161–1167. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.S.; Nuss, D.L. Infectious cDNA clone of hypovirus CHV1-Euro7: A comparative virology approach to investigate virus-mediated hypovirulence of the chestnut blight fungus Cryphonectria parasitica. J. Virol. 1999, 73, 985–992. [Google Scholar] [PubMed]
- Bryner, S.F.; Rigling, D. Temperature-dependent genotype-by-genotype interaction between a pathogenic fungus and its hyperparasitic virus. Am. Nat. 2011, 177, 65–74. [Google Scholar] [CrossRef]
- Hillman, B.I.; Suzuki, N. Viruses of the chestnut blight fungus, Cryphonectria parasitica. Adv. Virus Res. 2004, 63, 423–472. [Google Scholar]
- Prospero, S.; Lutz, A.; Tavadze, B.; Supatashvili, A.; Rigling, D. Discovery of a new gene pool and a high genetic diversity of the chestnut blight fungus Cryphonectria parasitica in Caucasian Georgia. Infect. Genet. Evol. 2013, 20, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Aghayeva, D.N.; Rigling, D.; Prospero, S. Low genetic diversity but frequent sexual reproduction of the chestnut blight fungus Cryphonectria parasitica in Azerbaijan. For. Pathol. 2017, 47, e12357. [Google Scholar] [CrossRef]
- Bissegger, M.; Rigling, D.; Heiniger, U. Population structure and disease development of Cryphonectria parasitica in European chestnut forests in the presence of natural hypovirulence. Phytopathology 1997, 87, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart model selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef]
- Vijaykrishna, D.; Mukerji, R.; Smith, G.J.D. RNA virus reassortment: An evolutionary mechanism for host jumps and immune evasion. PLoS Pathog. 2015, 11, e1004902. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Mendes, F.K.; Hahn, Y.; Hahn, M.W. Gene tree discordance can generate patterns of diminishing convergence over time. Mol. Biol. Evol. 2016, 33, 3299–3307. [Google Scholar] [CrossRef] [PubMed]
- Allemann, C.; Hoegger, P.; Heiniger, U.; Rigling, D. Genetic variation of Cryphonectria hypoviruses (CHV1) in Europe, assessed using restriction fragment length polymorphism (RFLP) markers. Mol. Ecol. 1999, 8, 843–854. [Google Scholar] [CrossRef]
- Wani, S.A.; Sahu, A.R.; Mishra, B.P.; Kumar, A.; Priya, G.B.; Padhy, A.; Sahoo, A.P.; Tiwari, A.K.; Gandham, R.K.; Singh, R.K. Whole genome sequence analysis of viruses; moving beyond single/partial gene based phylogenies in context of epidemiology and genetic evolution. Adv. Anim. Vet. Sci. 2015, 3, 435–443. [Google Scholar] [CrossRef]
- Kostaki, E.G.; Karamitros, T.; Stefanou, G.; Mamais, I.; Angelis, K.; Hatzakis, A.; Kramvis, A.; Paraskevis, D. Unravelling the history of hepatitis B virus genotypes A and D infection using a full-genome phylogenetic and phylogeographic approach. eLife 2018, 7, e36709. [Google Scholar] [CrossRef] [PubMed]
- Simon-Loriere, E.; Holmes, E.C. Why do RNA viruses recombine? Nat. Rev. Microbiol. 2011, 9, 617–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamora, P.; Martin, A.B.; Rigling, D.; Diez, J.J. Diversity of Cryphonectria parasitica in western Spain and identification of hypovirus-infected isolates. For. Pathol. 2012, 42, 412–419. [Google Scholar] [CrossRef]
- Bryner, S.F.; Rigling, D. Virulence not only costs but also benefits the transmission of a fungal virus. Evolution 2012, 66, 2540–2550. [Google Scholar] [CrossRef]
- Brusini, J.; Robin, C. Mycovirus transmission revisited by in situ pairings of vegetatively incompatible isolates of Cryphonectria parasitica. J. Virol. Methods 2013, 187, 435–442. [Google Scholar] [CrossRef]
- Jezic, M.; Krstin, L.; Rigling, D.; Curkovic-Perica, M. High diversity in populations of the introduced plant pathogen, Cryphonectria parasitica, due to encounters between genetically divergent genotypes. Mol. Ecol. 2012, 21, 87–99. [Google Scholar] [CrossRef]
- Prospero, S.; Rigling, D. Invasion genetics of the chestnut blight fungus Cryphonectria parasitica in Switzerland. Phytopathology 2012, 102, 73–82. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population 1 | Region | N C.p. 2 | N CHV-1 3 | % CHV-1 |
|---|---|---|---|---|
| Keda (Ked) | Adjara | 48 | 0 | 0 |
| Shemoqmedi (She) | Adjara | 47 | 14 | 29.8 |
| Tkilnari (Tkhi) | Adjara | 48 | 5 | 10.4 |
| Gezruli (Gez) | Imereti | 45 | 8 | 17.8 |
| Korbouli (Kor) | Imereti | 44 | 2 | 4.5 |
| Kumistavi (Kum) | Imereti | 50 | 12 | 24.0 |
| Satsable (Sab) | Imereti | 47 | 11 | 23.4 |
| Satsire (Sat) | Imereti | 71 | 3 | 4.2 |
| Tskalthashua (Tska) | Imereti | 46 | 4 | 8.7 |
| Mukhuri (Muk) | Samegrelo-Zemo Svaneti | 48 | 1 | 2.1 |
| Taleri (Tal) | Samegrelo-Zemo Svaneti | 43 | 2 | 4.7 |
| Axalsopeli (Axa) | Kakheti | 29 | 0 | 0 |
| Khecili (Khe) | Kakheti | 49 | 0 | 0 |
| Pshaveli (Psh) | Kakheti | 49 | 0 | 0 |
| Total | 664 | 62 | 9.3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rigling, D.; Borst, N.; Cornejo, C.; Supatashvili, A.; Prospero, S. Genetic and Phenotypic Characterization of Cryphonectria hypovirus 1 from Eurasian Georgia. Viruses 2018, 10, 687. https://doi.org/10.3390/v10120687
Rigling D, Borst N, Cornejo C, Supatashvili A, Prospero S. Genetic and Phenotypic Characterization of Cryphonectria hypovirus 1 from Eurasian Georgia. Viruses. 2018; 10(12):687. https://doi.org/10.3390/v10120687
Chicago/Turabian StyleRigling, Daniel, Nora Borst, Carolina Cornejo, Archil Supatashvili, and Simone Prospero. 2018. "Genetic and Phenotypic Characterization of Cryphonectria hypovirus 1 from Eurasian Georgia" Viruses 10, no. 12: 687. https://doi.org/10.3390/v10120687
APA StyleRigling, D., Borst, N., Cornejo, C., Supatashvili, A., & Prospero, S. (2018). Genetic and Phenotypic Characterization of Cryphonectria hypovirus 1 from Eurasian Georgia. Viruses, 10(12), 687. https://doi.org/10.3390/v10120687