Vertical and Horizontal Transmission of Pospiviroids


Abstract
:1. Introduction
2. Factors Affecting Vertical Transmission of Viroids
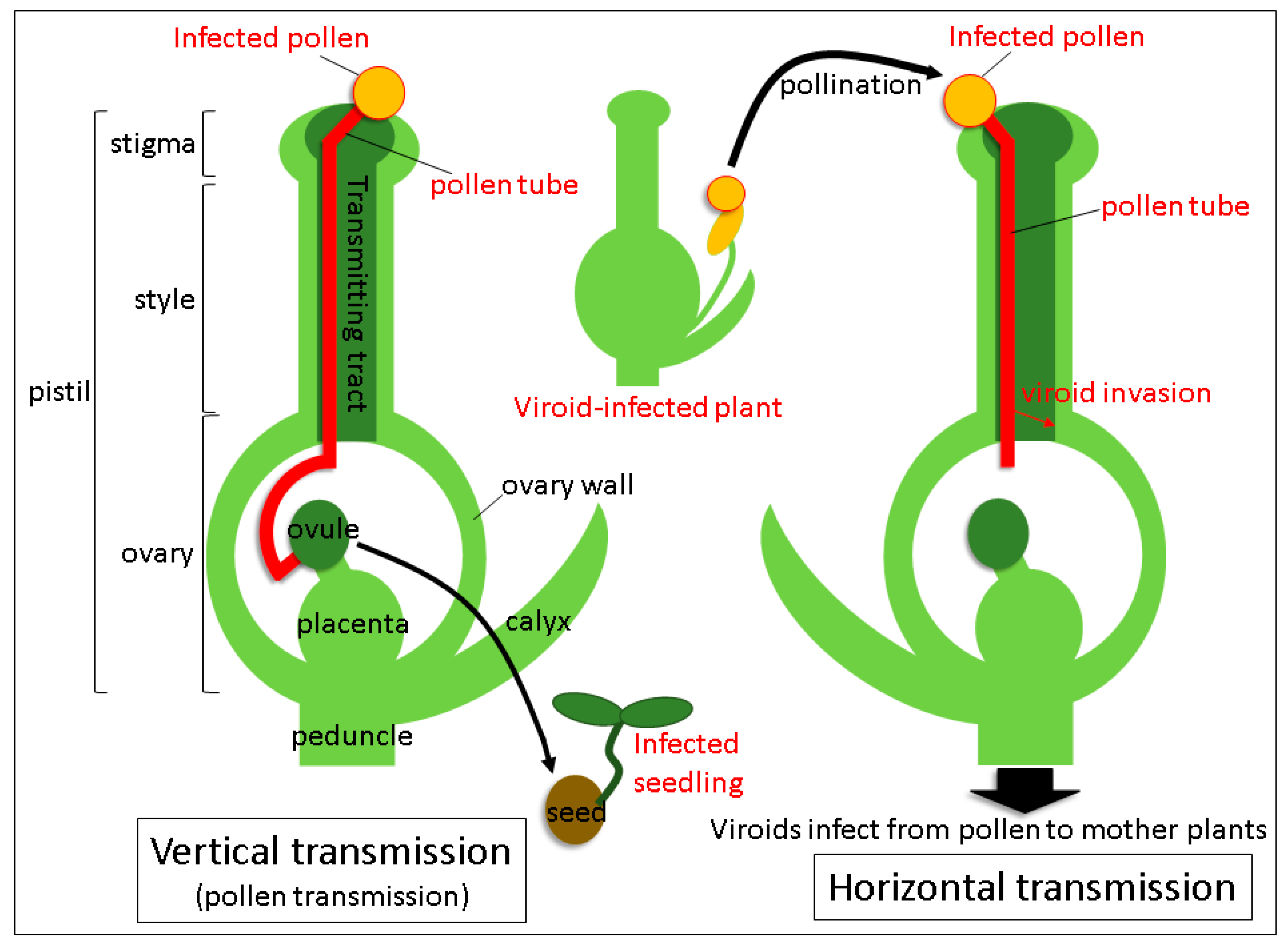
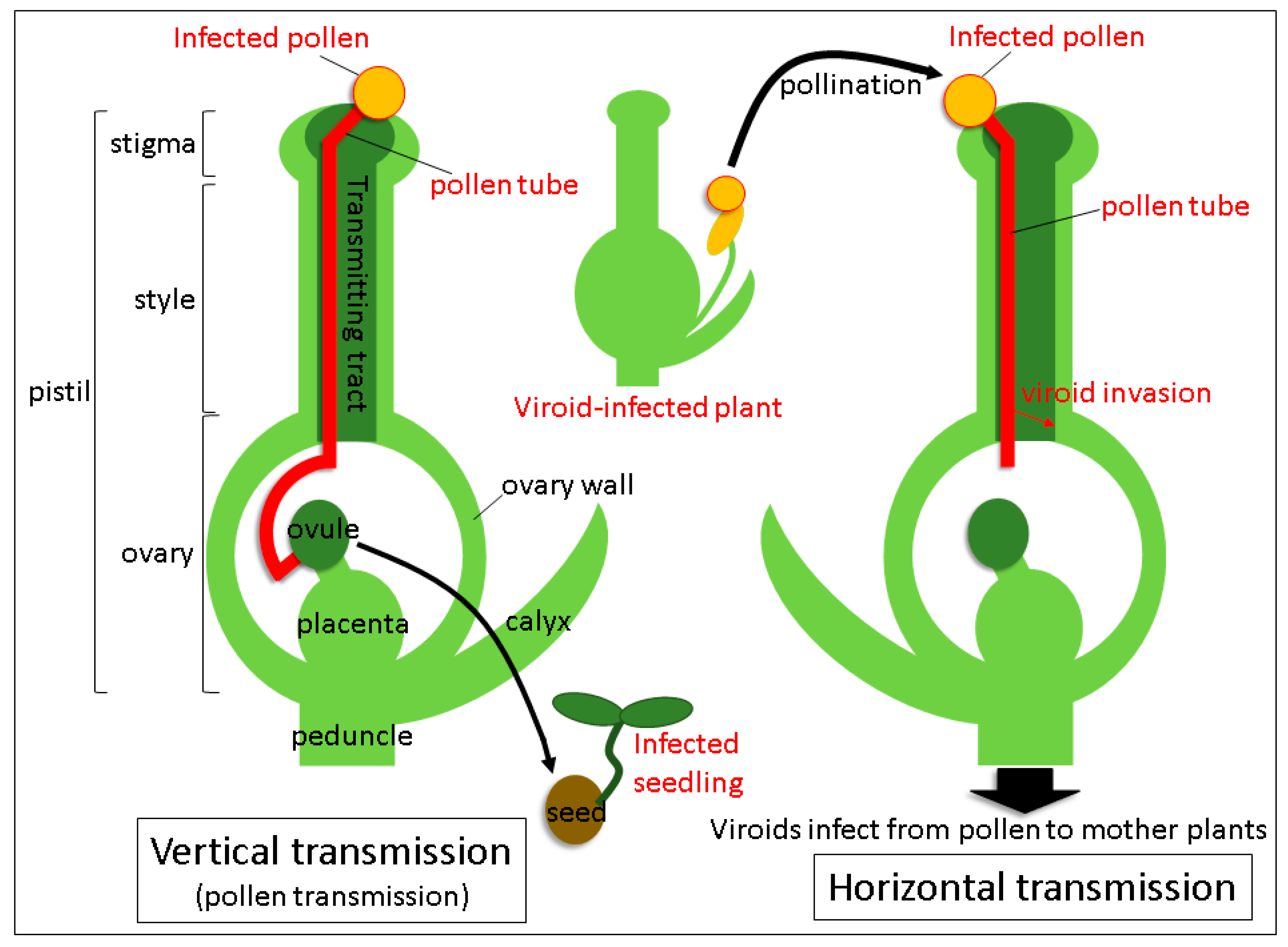
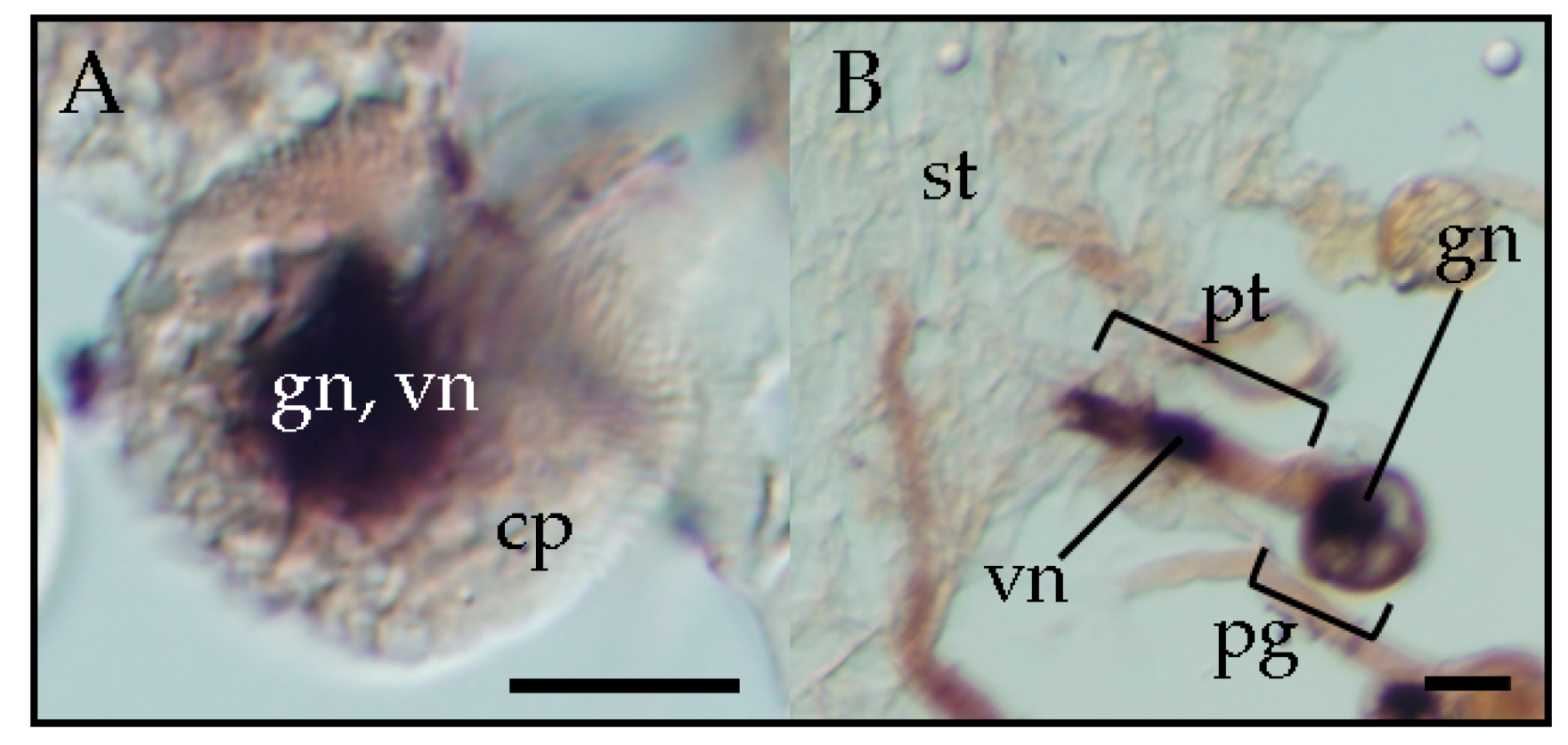
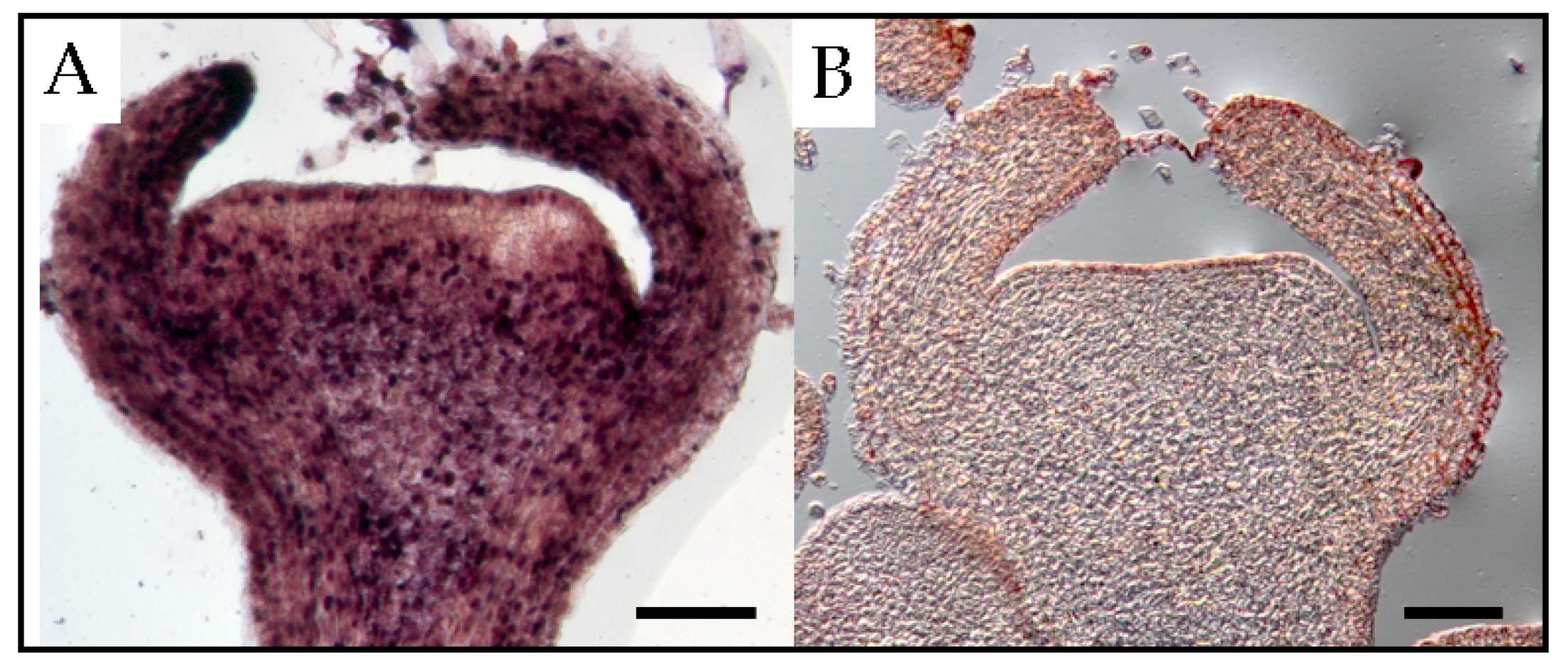
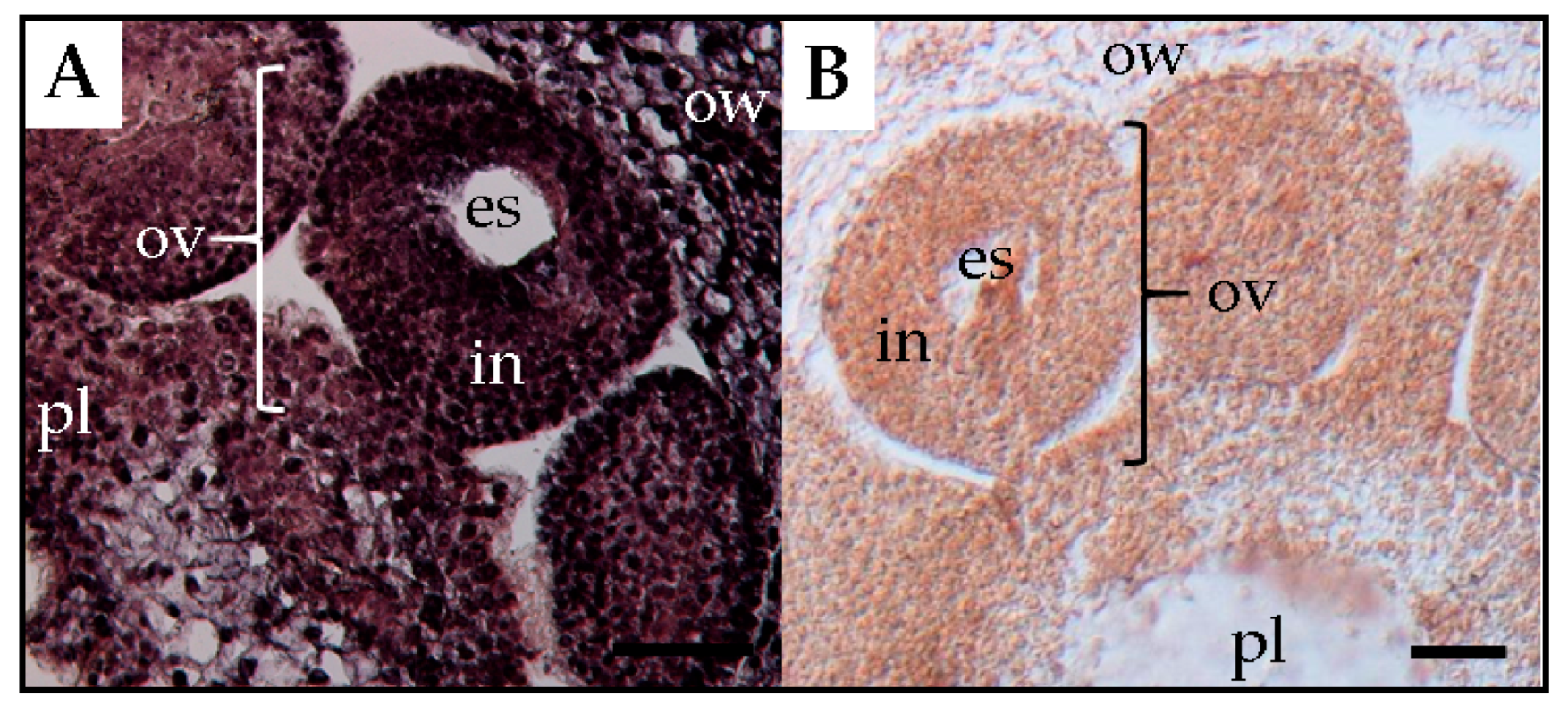
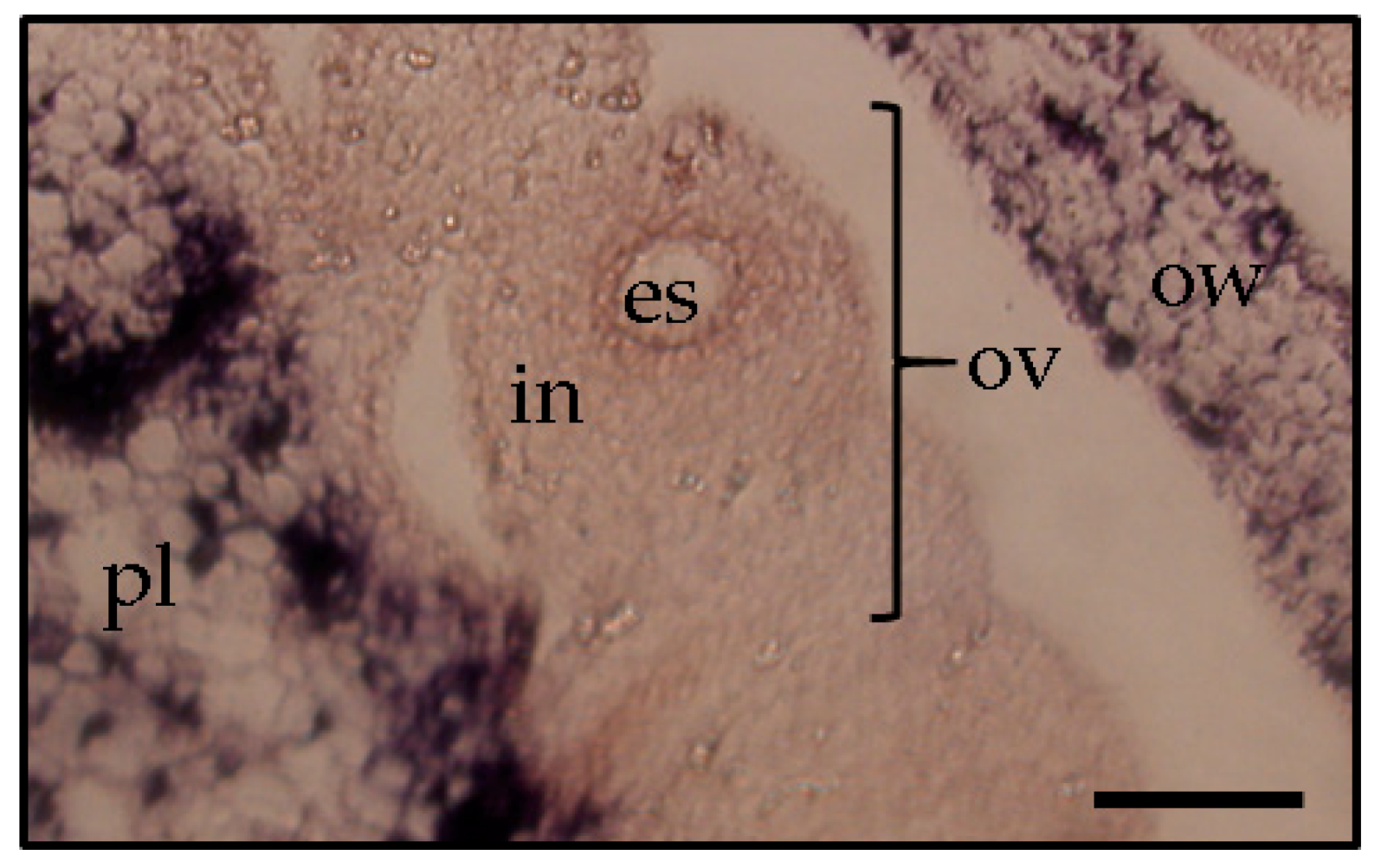
3. Trafficking of Viroids in Floral Organs and Pollen Tubes after Pollination with Infected Pollen
4. Mechanism of Seed Transmission
5. Mechanism of Horizontal Transmission
6. Viroid Nucleotide Sequences Affecting the Efficiency of Horizontal and Vertical Transmission
7. Further Prospects
Funding
Conflicts of Interest
References
- Hadidi, A.; Vidalakis, G.; Sano, T. Economic significance of fruit tree and grapivne viroids. In Viroids and Satellites; Hadidi, A., Flores, R., Palukaitis, P., Randles, J., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 15–26. ISBN 978-0-12-801498-1. [Google Scholar]
- Hammond, W.R. Economic significance of viroids in vegetable and filed crops. In Viroids and Satellites; Hadidi, A., Flores, R., Palukaitis, P., Randles, J., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 5–14. ISBN 978-0-12-801498-1. [Google Scholar]
- Di Serio, F.; Li, S.F.; Pallás, V.; Owens, R.A.; Randles, J.W.; Sano, T.; Verhoeven, J.T.; Vidalakis, G.; Flores, R. Viroid taxonomy. In Viroids and Satellites; Hadidi, A., Flores, R., Palukaitis, P., Randles, J., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 135–146. ISBN 978-0-12-801498-1. [Google Scholar]
- Flores, R.; Gago-Zachert, S.; Serra, P.; Sanjuan, R.; Elena, S.F. Viroids: Survivors from the RNA world? Annu. Rev. Microbiol. 2014, 68, 395–414. [Google Scholar] [CrossRef] [PubMed]
- Kovalskaya, N.; Hammond, R.W. Molecular biology of viroid–host interactions and disease control strategies. Plant Sci. 2014, 228, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.K.; Sinclair, B.V. Principles of Seed Pathology Volume I; CRC Press: Boca Raton, FL, USA, 1996; pp. 1–264. [Google Scholar]
- Sastry, K.S. Seed-Borne Plant Virus Diseases; Springer: New Delhi, India, 2013; pp. 1–53. ISBN 978-81-322-0812-9. [Google Scholar]
- Hamelin, F.M.; Allen, L.J.; Prendeville, H.R.; Hajimorad, M.R.; Jeger, M.J. The evolution of plant virus transmission pathways. J. Theor. Biol. 2016, 396, 75–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isogai, M.; Kamata, Y.; Ando, S.; Kamata, M.; Shirakawa, A.; Sekine, K.T.; Yoshikawa, N. Horizontal pollen transmission of Gentian ovary ring-spot virus is initiated during penetration of the stigma and style by infected pollen tubes. Virology 2017, 503, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Benson, A.P.; Singh, R.P. Seed transmission of potato spindle tuber virus in tomato. Am. Potato J. 1964, 41, 294. [Google Scholar]
- Singh, R.P. Seed transmission of potato spindle tuber virus in tomato and potato. Am. Potato J. 1970, 47, 225–227. [Google Scholar] [CrossRef]
- Tsuda, S.; Sano, T. Threats to Japanese agriculture from newly emerged plant viruses and viroids. J. Gen. Plant Pathol. 2014, 80, 2–14. [Google Scholar] [CrossRef]
- Card, S.D.; Person, M.N.; Clover, G.R.G. Plant pathogens transmitted by pollen. Australas. Plant Pathol. 2007, 36, 455–461. [Google Scholar] [CrossRef]
- Mink, G.I. Pollen and seed-transmitted viruses and viroids. Annu. Rev. Phytopathol. 1993, 31, 375–402. [Google Scholar] [CrossRef]
- Matsushita, Y.; Tsuda, S. Seed transmission of potato spindle tuber viroid, tomato chlorotic dwarf viroid, tomato apical stunt viroid, and Columnea latent viroid in horticultural plants. Eur. J. Plant Pathol. 2016, 145, 1007–1011. [Google Scholar] [CrossRef]
- Chung, B.N.; Choi, G.S. Incidence of coleus blumei viroid 1 in seeds of commercial coleus in Korea. Plant Pathol. J. 2008, 24, 305–308. [Google Scholar] [CrossRef]
- Singh, R.P.; Dilworth, D.A. Tomato chlorotic dwarf viroid in the ornamental plant Vinca minor and its transmission through tomato seed. Eur. J. Plant Pathol. 2009, 123, 111–116. [Google Scholar] [CrossRef]
- Matsushita, Y.; Usugi, T.; Tsuda, S. Distribution of tomato chlorotic dwarf viroid in floral organs of tomato. Eur. J. Plant Pathol. 2011, 130, 441–447. [Google Scholar] [CrossRef]
- Tsushima, T.; Sano, T. A point-mutation of Coleus blumei viroid 1 switches the potential to transmit through seed. J. Gen. Virol. 2018, 99, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Maula, A.J. Early embryo invasion as a determinant in pea of the seed transmission of pea seed-borne mosaic virus. J. Gen. Virol. 1992, 73, 1615–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Serio, F.; Martínez de Alba, A.E.; Navarro, B.A.; Gisel, A.; Flores, R. RNA-dependent RNA polymerase 6 delays accumulation and precludes meristem invasion of a viroid that replicates in the nucleus. J. Virol. 2010, 84, 2477–2489. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, Y.; Tsuda, S. Distribution of potato spindle tuber viroid in reproductive organs of petunia during its developmental stages. Phytopathology 2014, 104, 964–969. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, H.; Matsushita, Y. Host ranges and seed transmission of Tomato planta macho viroid and Pepper chat fruit viroid. Eur. J. Plant Pathol. 2017, 149, 211–217. [Google Scholar] [CrossRef]
- Yanagisawa, H.; Sano, T.; Hase, S.; Matsushita, Y. Influence of the terminal left domain on horizontal and vertical transmissions of tomato planta macho viroid and potato spindle tuber viroid through pollen. Virology 2019, 526, 22–31. [Google Scholar] [CrossRef]
- Fernow, K.H.; Peterson, L.C.; Plaisted, R.L. Spindle tuber virus in seeds and pollen of infected potato plants. Am. Potato J. 1970, 47, 75–80. [Google Scholar] [CrossRef]
- Hunter, D.E.; Darling, H.M.; Beale, W.L. Seed transmission of potato spindle tuber virus. Am. Potato J. 1969, 46, 247–250. [Google Scholar] [CrossRef]
- Kryczyński, S.; Paduch-Cichal, E.; Skrzeczkowski, L.J. Transmission of three viroids through seed and pollen of tomato plants. J. Phytopathol. 1988, 121, 51–57. [Google Scholar] [CrossRef]
- Verhoeven, J.T.J.; Jansen, C.C.C.; Roenhorst, J.W.; Flores, R.; de la Pena, M. Pepper chat fruit viroid: Biological and molecular properties of a proposed new species of the genus Pospiviroid. Virus Res. 2009, 144, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Semancik, J.S. Citrus exocortis viroid. CMI/AAB Descriptions Plant Viruses 1980, 226, 4. [Google Scholar]
- Singh, R.P. Detection of potato spindle tuber viroid in the pollen and various parts of potato plant pollinated with viroid-infected pollen. Plant Dis. 1992, 76, 951–953. [Google Scholar] [CrossRef]
- Singh, R.P.; Boucher, A.; Wang, R.G. Detection, distribution and long-term persistence of potato spindle tuber viroid in true potato seed from Heilongjiang China. Am. Potato J. 1991, 68, 65–74. [Google Scholar] [CrossRef]
- Yanagisawa, H.; Matsushita, Y. Differences in dynamics of horizontal transmission of tomato planta macho viroid and potato spindle tuber viroid after pollination with viroid-infected pollen. Virology 2018, 516, 258–264. [Google Scholar] [CrossRef]
- Matsushita, Y.; Yanagisawa, H. Distribution of tomato planta macho viroid in germinating pollen and transmitting tract. Virus Genes 2018, 54, 124–129. [Google Scholar] [CrossRef]
- Hanamata, S.; Kurusu, T.; Kuchitsu, K. Roles of autophagy in male reproductive development in plants. Front. Plant Sci. 2014, 5, 457. [Google Scholar] [CrossRef]
- Johansen, E.; Edwards, M.C.; Hampton, R.O. Seed transmission of viruses: Current perspectives. Annu. Rev. Phytopathol. 1994, 32, 363–386. [Google Scholar] [CrossRef]
- Ding, B.; Kwon, M.O.; Hammond, R.; Owens, R. Cell-to-cell movement of potato spindle tuber viroid. Plant J. 1997, 12, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Carroll, T.W.; Mayhew, D.E. Occurrence of virions in developing ovules and embryo sacs of barley in relation to the seed transmissibility of barley stripe mosaic virus. Can. J. Bot. 1976, 54, 2497–2512. [Google Scholar] [CrossRef]
- Gaspar, J.O.; Vega, J.; Camargo, I.J.B.; Costa, A.S. An ultrastructural study of particle distribution during microsporogenesis in tomato plants infected with the Brazilian tobacco rattle virus. Can. J. Bot. 1984, 62, 372–378. [Google Scholar] [CrossRef]
- Wilcoxson, R.D.; Johnson, L.E.B.; Frosheiser, F.I. Variation in the aggregation forms of alfalfa mosaic virus strains in different alfalfa organs. Phytopathology 1975, 65, 1249–1254. [Google Scholar] [CrossRef]
- Yang, A.F.; Hamilton, R.I. The mechanism of seed transmission of tobacco ringspot virus in soybean. Virology 1974, 62, 26–37. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P. Reassessment of the presence of viroid species of the genus Pospiviroid in infected floral parts, using reverse transcription—Polymerase chain reaction and infectivity assays. Can J. Plant Pathol. 2006, 28, 242–249. [Google Scholar] [CrossRef]
- Singh, D.; Mathur, S.B. Histopathology of Seed-borne Infections; CRC Press: Boca Raton, IL, USA, 2004; 282p, ISBN 0849328233. [Google Scholar]
- Isogai, M.; Suzuki, Y.; Matsudaira, T.; Yoshikawa, N. Genomic RNA accumulation of gentian ovary ring-spot virus and raspberry bushy dwarf virus in pollen tubes. J. Gen. Plant. Pathol. 2018, 84, 376–380. [Google Scholar] [CrossRef]
- Baek, Y.S.; Covey, P.A.; Petersen, J.J.; Chetelat, R.T.; McClure, B.; Bedinger, P.A. Testing the SI × SC rule: Pollen-pistil interactions in interspecific crosses between members of the tomato clade (Solanum section Lycopersicon, Solanaceae). Am. J. Bot. 2015, 102, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Škarić, D. Viroid Biology. In Viroids and Satellites; Hadidi, A., Flores, R., Palukaitis, P., Randles, J., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 53–61. ISBN 978-0-12-801498-1. [Google Scholar]
- Zhong, X.; Archual, A.J.; Amin, A.A.; Ding, B. A genomic map of viroid RNA motifs critical for replication and systemic trafficking. Plant Cell 2008, 20, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Steger, G.; Riesner, D.; Maurel, M.C.; Perreault, J.P. Viroid structure. In Viroids and Satellites; Hadidi, A., Flores, R., Palukaitis, P., Randles, J., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 63–70. ISBN 978-0-12-801498-1. [Google Scholar]
- Itaya, A.; Folimonov, A.; Matsuda, Y.; Nelson, R.S.; Ding, B. Potato spindle tuber viroid as inducer of RNA silencing in infected tomato. Mol. Plant Microbe Interact. 2001, 14, 1332–1334. [Google Scholar] [CrossRef] [PubMed]
- Castellano, M.; Martinez, G.; Marques, M.C.; Moreno-Romero, J.; Köhler, C.; Pallas, V.; Gomez, G. Changes in the DNA methylation pattern of the host male gametophyte of viroid-infected cucumber plants. J. Exp. Bot. 2016, 67, 5857–5868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, K.; Kusakari, S.; Kawaratani, M.; Negoro, J.; Ohki, S.; Osaki, T. Tobacco mosaic virus is transmissible from tomato to tomato by pollinating bumblebees. J. Gen. Plant Pathol. 2010, 66, 71–74. [Google Scholar] [CrossRef]
- Shipp, J.L.; Buitenhuis, R.; Stobbs, L.; Wang, K.; Ferguson, G. Vectoring of Pepino mosaic virus by bumble-bees in tomato greenhouses. Ann. Appl. Biol. 2008, 153, 149–155. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viroid | Host Plant Species | Seed-Transmitted | Pollen-Transmitted | Reference |
|---|---|---|---|---|
| Potato spindle tuber viroid | Potato | + | + | [11,25,26] |
| Tomato | + | + | [10,11,15,27] | |
| Capsicum annum | + | [15] | ||
| Glebionis coronaria | + | [15] | ||
| Petunia hybrida | + | + | [22,23] | |
| Tomato chlorotic dwarf viroid | Tomato | + | [17] | |
| Petunia hybrida | + | [15] | ||
| Tomato apical stunt viroid | Tomato | + | [15] | |
| Tomato planta macho viroid | Tomato | + | [23] | |
| Petunia hybrida | + | + | [23] | |
| Pepper chat fruit viroid | Tomato | + | [23] | |
| Capsicum annum | + | [28] | ||
| Petunia hybrida | + | + | [23] | |
| Columunea latent viroid | Tomato | + | [15] | |
| Chrysanthemum stunt viroid | Tomato | + | + | [27] |
| Chrysahtmemum | + | + | [16] | |
| Citrus exocortis viroid | Tomato | + | [29] | |
| Impatiens | + | [29] | ||
| Verbena | + | [29] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsushita, Y.; Yanagisawa, H.; Sano, T. Vertical and Horizontal Transmission of Pospiviroids. Viruses 2018, 10, 706. https://doi.org/10.3390/v10120706
Matsushita Y, Yanagisawa H, Sano T. Vertical and Horizontal Transmission of Pospiviroids. Viruses. 2018; 10(12):706. https://doi.org/10.3390/v10120706
Chicago/Turabian StyleMatsushita, Yosuke, Hironobu Yanagisawa, and Teruo Sano. 2018. "Vertical and Horizontal Transmission of Pospiviroids" Viruses 10, no. 12: 706. https://doi.org/10.3390/v10120706
APA StyleMatsushita, Y., Yanagisawa, H., & Sano, T. (2018). Vertical and Horizontal Transmission of Pospiviroids. Viruses, 10(12), 706. https://doi.org/10.3390/v10120706