Applying Unique Molecular Identifiers in Next Generation Sequencing Reveals a Constrained Viral Quasispecies Evolution under Cross-Reactive Antibody Pressure Targeting Long Alpha Helix of Hemagglutinin

 , , , ,
, , , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
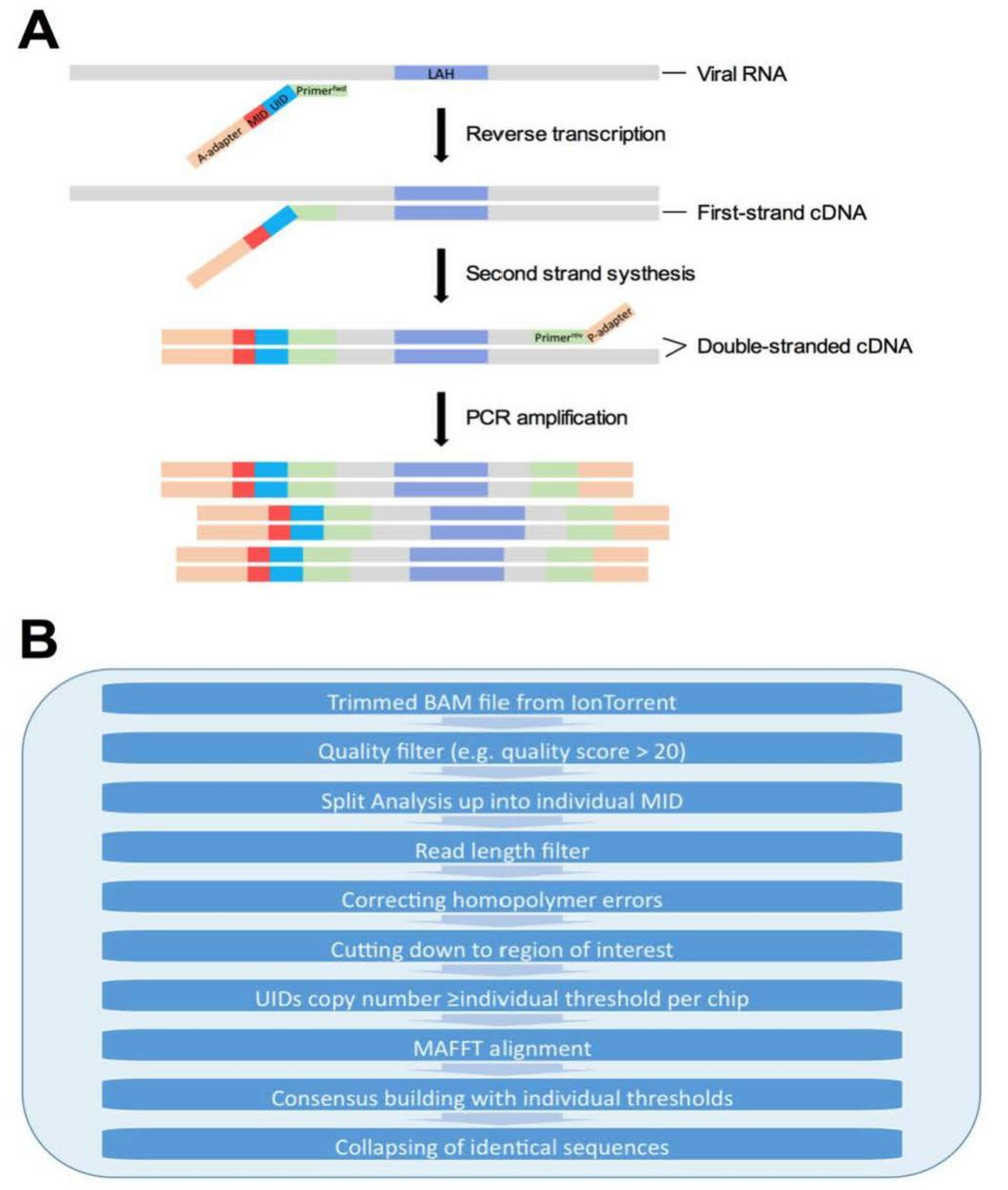{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mice and Viral Infection
2.2. Ethics Statement
2.3. ELISA
2.4. Virus Culture, Titrations and Lung Titer
2.5. RNA Extraction and Library Preparation
2.6. High-Throughput Next Generation Sequencing
2.7. NGS Data Analysis
2.8. Statistical Analysis
3. Results
3.1. Induction of Broadly Reactive Anti-LAH Antibodies by Immunization
3.2. Reduced Lung Virus Titers in Immunized Mice
3.3. Decreased Diversity of LAH Epitopes
3.4. Absence of Escape Mutants Following Vaccination
3.5. Identification of Diversified Positions on LAH Epitope
3.6. Analysis of Amino Acid Substitutions
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Funding
References
- Krammer, F.; Palese, P. Advances in the development of influenza virus vaccines. Nat. Rev. Drug Discov. 2015, 14, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Korownyk, C.; Garrison, S.; Kolber, M.R. Antiviral medications for influenza. Can. Fam. Physician 2015, 61, 351. [Google Scholar] [PubMed]
- Gerdil, C. The annual production cycle for influenza vaccine. Vaccine 2003, 21, 1776–1779. [Google Scholar] [CrossRef]
- Palese, P. Influenza: Old and new threats. Nat. Med. 2004, 10, S82–S87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, L.; Compans, R.W.; Wang, B.Z. Universal influenza vaccines, a dream to be realized soon. Viruses 2014, 6, 1974–1991. [Google Scholar] [CrossRef] [PubMed]
- Steel, J.; Lowen, A.C.; Wang, T.T.; Yondola, M.; Gao, Q.; Haye, K.; Garcia-Sastre, A.; Palese, P. Influenza virus vaccine based on the conserved hemagglutinin stalk domain. mBio 2010, 1, e00018-10. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Tan, G.S.; Hai, R.; Pica, N.; Ngai, L.; Ekiert, D.C.; Wilson, I.A.; Garcia-Sastre, A.; Moran, T.M.; Palese, P. Vaccination with a synthetic peptide from the influenza virus hemagglutinin provides protection against distinct viral subtypes. Proc. Natl. Acad. Sci. USA 2010, 107, 18979–18984. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Chen, S.; Qu, D.; Chen, J.; Wang, F.; Zhang, R.; Chen, Z. Influenza H7N9 LAH-HBc virus-like particle vaccine with adjuvant protects mice against homologous and heterologous influenza viruses. Vaccine 2016, 34, 6464–6471. [Google Scholar] [CrossRef] [PubMed]
- Kazaks, A.; Lu, I.N.; Farinelle, S.; Ramirez, A.; Crescente, V.; Blaha, B.; Ogonah, O.; Mukhopadhyay, T.; de Obanos, M.P.; Krimer, A.; et al. Production and purification of chimeric HBc virus-like particles carrying influenza virus LAH domain as vaccine candidates. BMC Biotechnol. 2017, 17, 79. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.J.; Das, S.R.; Wang, W.; Fitzgerald, T.; Pickett, B.E.; Aevermann, B.D.; Topham, D.J.; Falsey, A.R.; Scheuermann, R.H. Diversifying selection analysis predicts antigenic evolution of 2009 pandemic H1N1 influenza A virus in humans. J. Virol. 2015, 89, 5427–5440. [Google Scholar] [CrossRef] [PubMed]
- Van den Hoecke, S.; Verhelst, J.; Vuylsteke, M.; Saelens, X. Analysis of the genetic diversity of influenza A viruses using next-generation DNA sequencing. BMC Genom. 2015, 16, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zharikova, D.; Mozdzanowska, K.; Feng, J.; Zhang, M.; Gerhard, W. Influenza type a virus escape mutants emerge in vivo in the presence of antibodies to the ectodomain of matrix protein 2. J. Virol. 2005, 79, 6644–6654. [Google Scholar] [CrossRef] [PubMed]
- Gerhard, W.; Mozdzanowska, K.; Zharikova, D. Prospects for universal influenza virus vaccine. Emerg. Infect. Dis. 2006, 12, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Kaverin, N.V.; Rudneva, I.A.; Ilyushina, N.A.; Varich, N.L.; Lipatov, A.S.; Smirnov, Y.A.; Govorkova, E.A.; Gitelman, A.K.; Lvov, D.K.; Webster, R.G. Structure of antigenic sites on the haemagglutinin molecule of H5 avian influenza virus and phenotypic variation of escape mutants. J. Gen. Virol. 2002, 83, 2497–2505. [Google Scholar] [CrossRef] [PubMed]
- Eisfeld, A.J.; Neumann, G.; Kawaoka, Y. Influenza A virus isolation, culture and identification. Nat. Protoc. 2014, 9, 2663–2681. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E.; Sheldon, J.; Perales, C. Viral quasispecies evolution. Microbiol. Mol. Biol. Rev. 2012, 76, 159–216. [Google Scholar] [CrossRef] [PubMed]
- Kinde, I.; Wu, J.; Papadopoulos, N.; Kinzler, K.W.; Vogelstein, B. Detection and quantification of rare mutations with massively parallel sequencing. Proc. Natl. Acad. Sci. USA 2011, 108, 9530–9535. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Vijaya Satya, R.; Lewis, M.; Randad, P.; Wang, Y. Reducing amplification artifacts in high multiplex amplicon sequencing by using molecular barcodes. BMC Genom. 2015, 16, 589. [Google Scholar] [CrossRef] [PubMed]
- Peyret, H.; Gehin, A.; Thuenemann, E.C.; Blond, D.; El Turabi, A.; Beales, L.; Clarke, D.; Gilbert, R.J.; Fry, E.E.; Stuart, D.I.; et al. Tandem fusion of hepatitis B core antigen allows assembly of virus-like particles in bacteria and plants with enhanced capacity to accommodate foreign proteins. PLoS ONE 2015, 10, e0120751. [Google Scholar] [CrossRef] [PubMed]
- Lu, I.N.; Farinelle, S.; Sausy, A.; Muller, C.P. Identification of a CD4 T-cell epitope in the hemagglutinin stalk domain of pandemic H1N1 influenza virus and its antigen-driven TCR usage signature in BALB/c mice. Cell. Mol. Immunol. 2017, 14, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Theisen, L.L.; Muller, C.P. EPs® 7630 (umckaloabo®), an extract from Pelargonium sidoides roots, exerts anti-influenza virus activity in vitro and in vivo. Antivir. Res. 2012, 94, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Vollmers, C.; Sit, R.V.; Weinstein, J.A.; Dekker, C.L.; Quake, S.R. Genetic measurement of memory B-cell recall using antibody repertoire sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 13463–13468. [Google Scholar] [CrossRef] [PubMed]
- Loman, N.J.; Misra, R.V.; Dallman, T.J.; Constantinidou, C.; Gharbia, S.E.; Wain, J.; Pallen, M.J. Performance comparison of benchtop high-throughput sequencing platforms. Nat. Biotechnol. 2012, 30, 434–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, C.D.; Vogel, L.; Wright, A.; Das, S.R.; Wrammert, J.; Li, G.M.; McCausland, M.; Zheng, N.Y.; Yewdell, J.W.; Ahmed, R.; et al. Antibody pressure by a human monoclonal antibody targeting the 2009 pandemic H1N1 virus hemagglutinin drives the emergence of a virus with increased virulence in mice. mBio 2012, 3, e00120-12. [Google Scholar] [CrossRef]
- Luo, C.; Hirsch, H.H.; Kant, J.; Randhawa, P. VP-1 quasispecies in human infection with polyomavirus BK. J. Med. Virol. 2012, 84, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Huang, W.; Kang, J.; Huang, Y.; Ren, H.; Ding, K. Comparison of error correction algorithms for ion torrent PGM data: Application to hepatitis B virus. Sci. Rep. 2017, 7, 8106. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, S.; McPherson, J.D.; McCombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef] [PubMed]
- Rapaport, F.; Khanin, R.; Liang, Y.; Pirun, M.; Krek, A.; Zumbo, P.; Mason, C.E.; Socci, N.D.; Betel, D. Comprehensive evaluation of differential gene expression analysis methods for RNA-seq data. Genome Biol. 2013, 14, 3158. [Google Scholar] [CrossRef] [PubMed]
- Pawelek, K.A.; Huynh, G.T.; Quinlivan, M.; Cullinane, A.; Rong, L.; Perelson, A.S. Modeling within-host dynamics of influenza virus infection including immune responses. PLoS Comput. Biol. 2012, 8, e1002588. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Vargas, E.A.; Wilk, E.; Canini, L.; Toapanta, F.R.; Binder, S.C.; Uvarovskii, A.; Ross, T.M.; Guzman, C.A.; Perelson, A.S.; Meyer-Hermann, M. Effects of aging on influenza virus infection dynamics. J. Virol. 2014, 88, 4123–4131. [Google Scholar] [CrossRef] [PubMed]
- Barouch, D.H.; Ghneim, K.; Bosche, W.J.; Li, Y.; Berkemeier, B.; Hull, M.; Bhattacharyya, S.; Cameron, M.; Liu, J.; Smith, K.; et al. Rapid inflammasome activation following mucosal SIV infection of rhesus monkeys. Cell 2016, 165, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Bull, R.A.; Luciani, F.; McElroy, K.; Gaudieri, S.; Pham, S.T.; Chopra, A.; Cameron, B.; Maher, L.; Dore, G.J.; White, P.A.; et al. Sequential bottlenecks drive viral evolution in early acute hepatitis C virus infection. PLoS Pathog. 2011, 7, e1002243. [Google Scholar] [CrossRef] [PubMed]
- Laskus, T.; Wilkinson, J.; Gallegos-Orozco, J.F.; Radkowski, M.; Adair, D.M.; Nowicki, M.; Operskalski, E.; Buskell, Z.; Seeff, L.B.; Vargas, H.; et al. Analysis of hepatitis C virus quasispecies transmission and evolution in patients infected through blood transfusion. Gastroenterology 2004, 127, 764–776. [Google Scholar] [CrossRef] [PubMed]
- McMichael, A.J.; Borrow, P.; Tomaras, G.D.; Goonetilleke, N.; Haynes, B.F. The immune response during acute hiv-1 infection: Clues for vaccine development. Nat. Rev. Immunol. 2010, 10, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E.; Holland, J.J. RNA virus mutations and fitness for survival. Annu. Rev. Microbiol. 1997, 51, 151–178. [Google Scholar] [CrossRef] [PubMed]
- Sriwilaijaroen, N.; Suzuki, Y. Molecular basis of the structure and function of H1 hemagglutinin of influenza virus. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2012, 88, 226–249. [Google Scholar] [CrossRef] [PubMed]
- Perdue, M.L. Naturally occurring NS gene variants in an avian influenza virus isolate. Virus Res. 1992, 23, 223–240. [Google Scholar] [CrossRef]
- Wang, L.; Suarez, D.L.; Pantin-Jackwood, M.; Mibayashi, M.; Garcia-Sastre, A.; Saif, Y.M.; Lee, C.W. Characterization of influenza virus variants with different sizes of the non-structural (NS) genes and their potential as a live influenza vaccine in poultry. Vaccine 2008, 26, 3580–3586. [Google Scholar] [CrossRef] [PubMed]
- Dundon, W.G. Variability among the neuraminidase, non-structural 1 and PB1-F2 proteins in the influenza A virus genome. Virus Genes 2012, 44, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Chai, N.; Swem, L.R.; Reichelt, M.; Chen-Harris, H.; Luis, E.; Park, S.; Fouts, A.; Lupardus, P.; Wu, T.D.; Li, O.; et al. Two escape mechanisms of influenza a virus to a broadly neutralizing stalk-binding antibody. PLoS Pathog. 2016, 12, e1005702. [Google Scholar] [CrossRef] [PubMed]
- Valkenburg, S.A.; Rutigliano, J.A.; Ellebedy, A.H.; Doherty, P.C.; Thomas, P.G.; Kedzierska, K. Immunity to seasonal and pandemic influenza A viruses. Microbes Infect. 2011, 13, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Lucas, M.; Karrer, U.; Lucas, A.; Klenerman, P. Viral escape mechanisms—Escapology taught by viruses. Int. J. Exp. Pathol. 2001, 82, 269–286. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hauck, N.C.; Kirpach, J.; Kiefer, C.; Farinelle, S.; Maucourant, S.; Morris, S.A.; Rosenberg, W.; He, F.Q.; Muller, C.P.; Lu, I.-N. Applying Unique Molecular Identifiers in Next Generation Sequencing Reveals a Constrained Viral Quasispecies Evolution under Cross-Reactive Antibody Pressure Targeting Long Alpha Helix of Hemagglutinin. Viruses 2018, 10, 148. https://doi.org/10.3390/v10040148
Hauck NC, Kirpach J, Kiefer C, Farinelle S, Maucourant S, Morris SA, Rosenberg W, He FQ, Muller CP, Lu I-N. Applying Unique Molecular Identifiers in Next Generation Sequencing Reveals a Constrained Viral Quasispecies Evolution under Cross-Reactive Antibody Pressure Targeting Long Alpha Helix of Hemagglutinin. Viruses. 2018; 10(4):148. https://doi.org/10.3390/v10040148
Chicago/Turabian StyleHauck, Nastasja C., Josiane Kirpach, Christina Kiefer, Sophie Farinelle, Sophie Maucourant, Stephen A. Morris, William Rosenberg, Feng Q. He, Claude P. Muller, and I-Na Lu. 2018. "Applying Unique Molecular Identifiers in Next Generation Sequencing Reveals a Constrained Viral Quasispecies Evolution under Cross-Reactive Antibody Pressure Targeting Long Alpha Helix of Hemagglutinin" Viruses 10, no. 4: 148. https://doi.org/10.3390/v10040148
APA StyleHauck, N. C., Kirpach, J., Kiefer, C., Farinelle, S., Maucourant, S., Morris, S. A., Rosenberg, W., He, F. Q., Muller, C. P., & Lu, I.-N. (2018). Applying Unique Molecular Identifiers in Next Generation Sequencing Reveals a Constrained Viral Quasispecies Evolution under Cross-Reactive Antibody Pressure Targeting Long Alpha Helix of Hemagglutinin. Viruses, 10(4), 148. https://doi.org/10.3390/v10040148