Yeast Derived LysA2 Can Control Bacterial Contamination in Ethanol Fermentation



Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial and Yeast Strains
2.2. Construction of a LysA2 Bacterial and Yeast Expression Vectors
2.3. Bacterial and Yeast Transformation with the LysA2 Bacterial and Yeast Expression Vectors
2.4. Protein Expression and Purification
2.5. Turbidity Reduction Assay
2.6. Fermentation with Simulated Contamination
3. Results
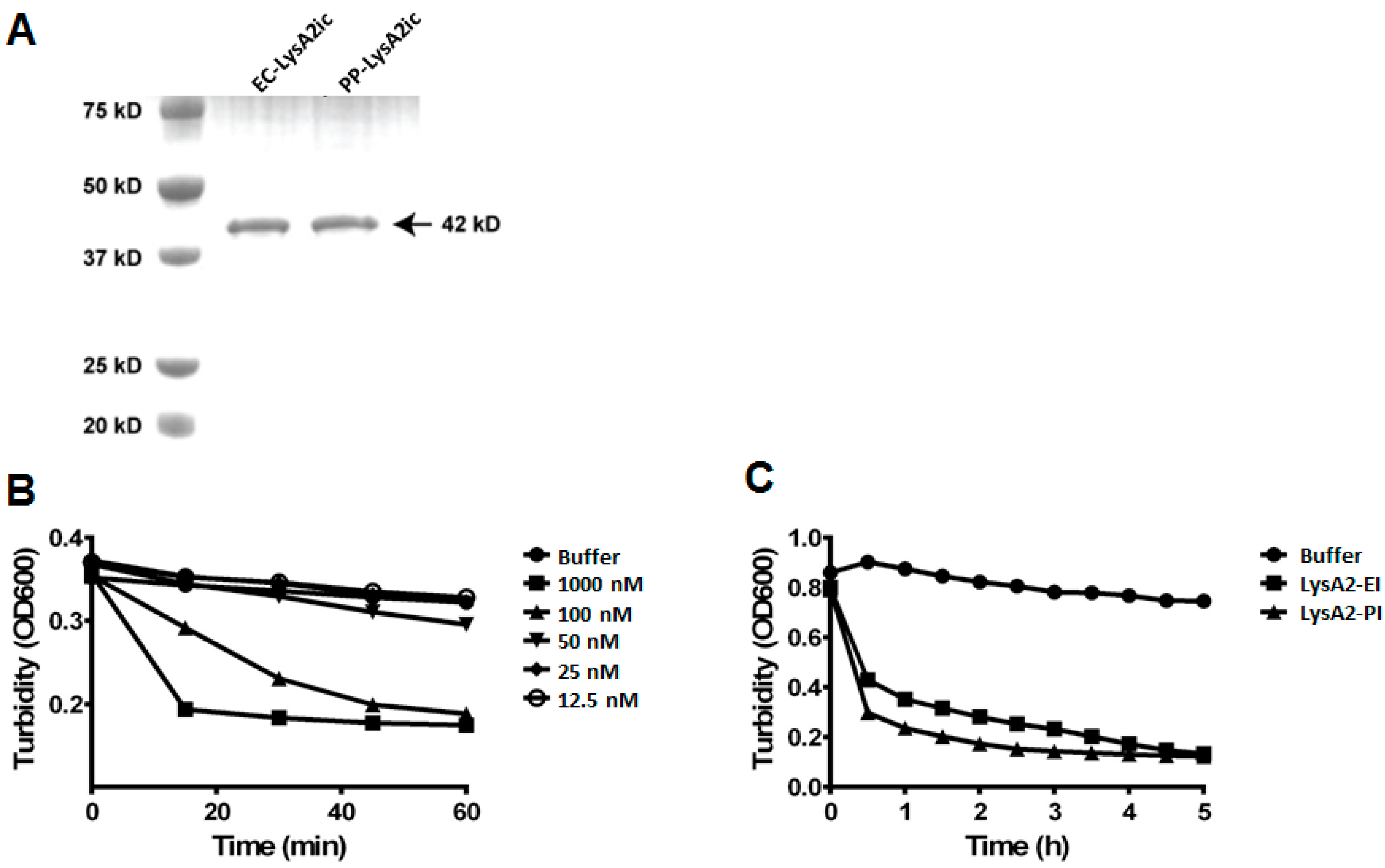
3.1. Comparison of Lytic Activity of LysA2 Expressed by Bacteria and Yeast
3.2. Lytic Activity of Intracellularly Produced LysA2 from P. pastoris on Common Biofuel Contaminants
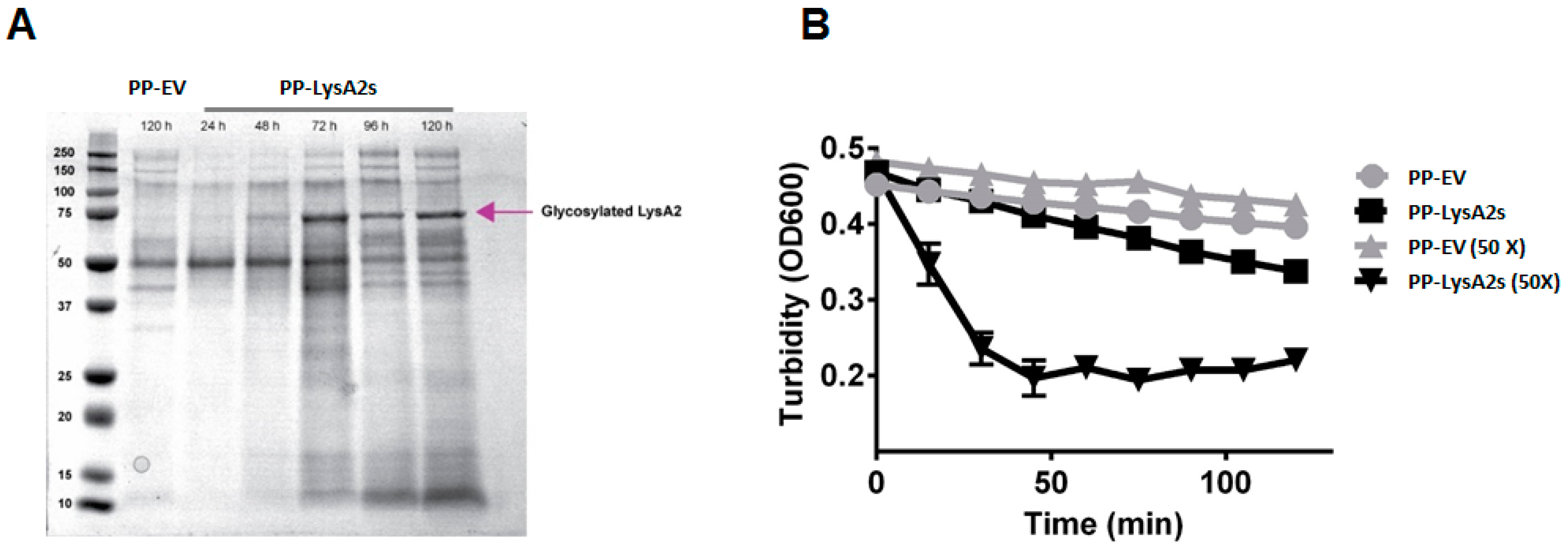
3.3. Secreted LysA2 and Its Lytic Activity
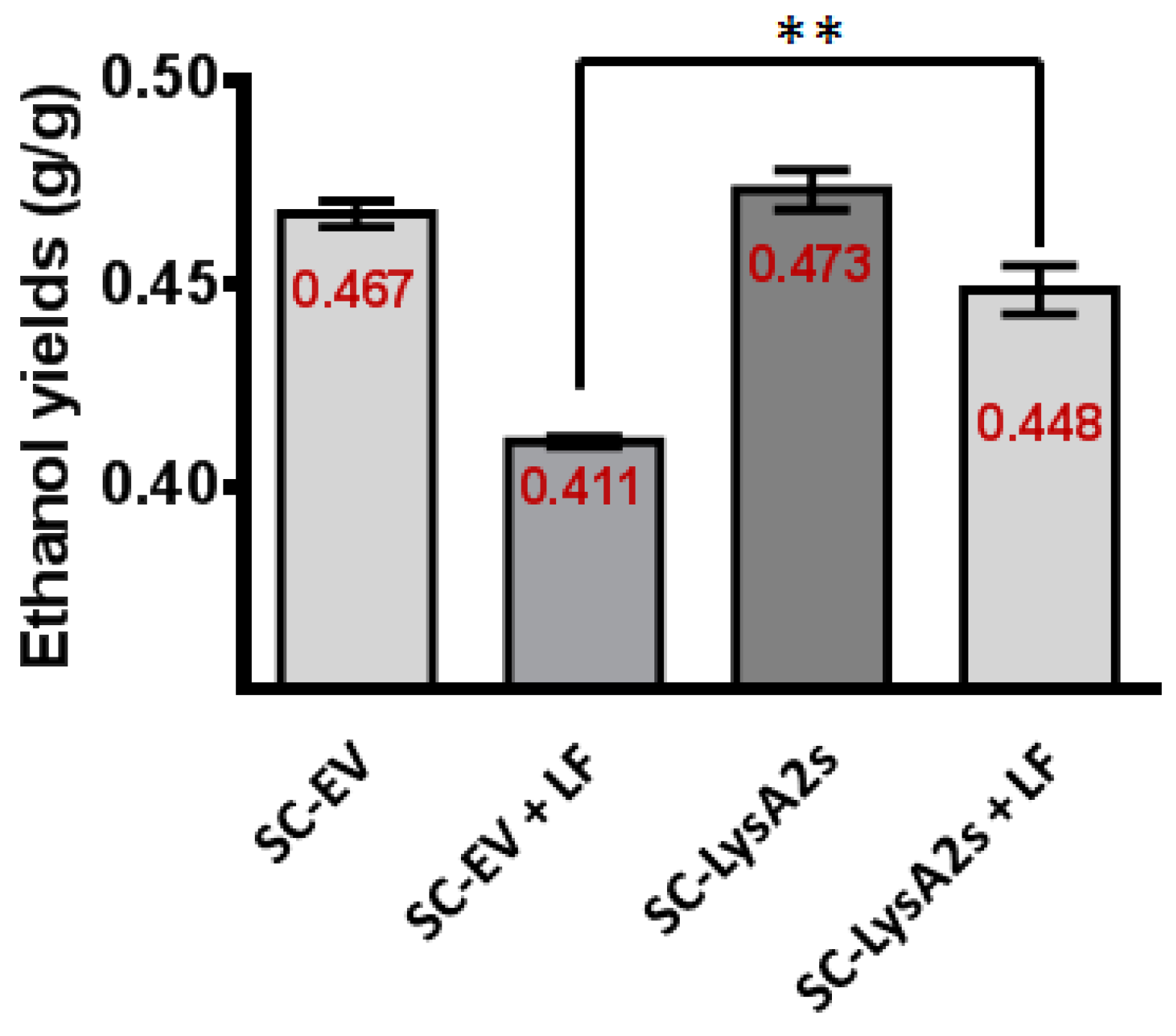
3.4. S. cerevisiae Secreting LysA2 Restores Ethanol Yield in Mock Contamination of Fermentation
3.5. Mock Contamination with Sugarcane Juice
4. Discussion
4.1. Bacteria and Yeast-Based LysA2 Showed the Same Lytic Activity and Spectrum
4.2. Secreted Lysa2 from P. Pastoris Successfully Restores Ethanol Productivity during Bacterial Contamination
4.3. S. cerevisiae Secreting LysA2 is Effective Against Bacterial Contamination in YPD Medium
4.4. S. cerevisiae Secreting LysA2 Fails to Protect Against Bacterial Contamination in Sugarcane Juice
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Renewable Fuels Association (RFA). Going Global—2015 Ethanol Industry Outlook; RFA: Washington, DC, USA, 2015; p. 4. [Google Scholar]
- Amorim, H.V.; Lopes, M.L.; de Castro Oliveira, J.V.; Buckeridge, M.S.; Goldman, G.H. Scientific challenges of bioethanol production in Brazil. Appl. Microbiol. Biotechnol. 2011, 91, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, K.M.; Liu, S.; Leathers, T.D.; Worthington, R.E.; Rich, J.O. Modeling bacterial contamination of fuel ethanol fermentation. Biotechnol. Bioeng. 2009, 103, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Bischoff, K.M.; Gill, J.J.; Mire-Criscione, M.D.; Berry, J.D.; Young, R.; Summer, E.J. Bacteriophage application restores ethanol fermentation characteristics disrupted by Lactobacillus fermentum. Biotechnol. Biofuels 2015, 8, 132. [Google Scholar] [CrossRef] [PubMed]
- Lucena, B.T.L.; dos Santos, B.M.; Moreira, J.L.S.; Moreira, A.P.B.; Nunes, A.C.; Azevedo, V.; Miyoshi, A.; Thompson, F.L.; de Morais, M. Diversity of lactic acid bacteria of the bioethanol process. BMC Microbiol. 2010, 10, 298. [Google Scholar] [CrossRef] [PubMed]
- Jacques, K.A.; Lyons, T.P.; Kelsall, D.R. The Alcohol Textbook: A Reference for the Beverage, Fuel and Industrial Alcohol Industries; Nottingham University Press: Nottingham, UK, 2003. [Google Scholar]
- Scott, Lewis. Options Expand for Effective Bacterial Control in Ethanol Production. Available online: http://ethanolproducer.com/articles/13871/options-expand-for-effective-bacterial-control-in-ethanol-production (accessed on 10 November 2016).
- Skinner, K.A.; Leathers, T.D. Bacterial contaminants of fuel ethanol production. J. Ind. Microbiol. Biotechnol. 2004, 31, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Guzzo, F.; Cappello, M.S.; Azzolini, M.; Tosi, E.; Zapparoli, G. The inhibitory effects of wine phenolics on lysozyme activity against lactic acid bacteria. Int. J. Food Microbiol. 2011, 148, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.; O’Sullivan, O.; Mills, S.; McAuliffe, O.; Ross, R.P.; Neve, H.; Coffey, A. Genome sequence of the phage clP1, which infects the beer spoilage bacterium Pediococcus damnosus. Gene 2012, 504, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.L.; Pettersson, B.; Priest, F.G. Characterization of lactobacilli from Scotch malt whisky distilleries and description of Lactobacillus ferintoshensis sp. nov., a new species isolated from malt whisky fermentations. Microbiology 2001, 147, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Costa, O.Y.A.; Souto, B.M.; Tupinambá, D.D.; Bergmann, J.C.; Kyaw, C.M.; Kruger, R.H.; Barreto, C.C.; Quirino, B.F. Microbial diversity in sugarcane ethanol production in a Brazilian distillery using a culture-independent method. J. Ind. Microbiol. Biotechnol. 2015, 42, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Schell, D.J.; Dowe, N.; Ibsen, K.N.; Riley, C.J.; Ruth, M.F.; Lumpkin, R.E. Contaminant occurrence, identification and control in a pilot-scale corn fiber to ethanol conversion process. Bioresour. Technol. 2007, 98, 2942–2948. [Google Scholar] [CrossRef] [PubMed]
- Basso, T.O.; de Kok, S.; Dario, M.; do Espirito-Santo, J.C.A.; Müller, G.; Schlölg, P.S.; Silva, C.P.; Tonso, A.; Daran, J.-M.; Gombert, A.K.; et al. Engineering topology and kinetics of sucrose metabolism in Saccharomyces cerevisiae for improved ethanol yield. Metab. Eng. 2011, 13, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, K.M.; Skinner-Nemec, K.A.; Leathers, T.D. Antimicrobial susceptibility of Lactobacillus species isolated from commercial ethanol plants. J. Ind. Microbiol. Biotechnol. 2007, 34, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Rich, J.O.; Leathers, T.D.; Nunnally, M.S.; Bischoff, K.M. Rapid evaluation of the antibiotic susceptibility of fuel ethanol contaminant biofilms. Bioresour. Technol. 2011, 102, 1124–1130. [Google Scholar] [CrossRef] [PubMed]
- Oliva Neto, P.D.; Lima, F.A.D.; Silva, K.C.D.; Silva, D.F.D.; Carvalho, A.F.A.; Santos, C.D. Chemical inhibition of the contaminant Lactobacillus fermentum from distilleries producing fuel bioethanol. Braz. Arch. Biol. Technol. 2014, 57, 441–447. [Google Scholar] [CrossRef]
- Stroppa, C.T.; Andrietta, M.G.S.; Andrietta, S.R.; Steckelberg, C.; Serra, G. Use of Penicillin and Monensin to Control Bacterial Contamination of Brazilian Alcohol Fermentations. Int. Sugar J. 2000, 102, 78–82. [Google Scholar]
- Murphree, C.A.; Heist, E.P.; Moe, L.A. Antibiotic Resistance Among Cultured Bacterial Isolates from Bioethanol Fermentation Facilities Across the United States. Curr. Microbiol. 2014, 69, 277–285. [Google Scholar] [CrossRef] [PubMed]
- De Alwis, H.; Heller, D.N. Multiclass, multiresidue method for the detection of antibiotic residues in distillers grains by liquid chromatography and ion trap tandem mass spectrometry. J. Chromatogr. A 2010, 1217, 3076–3084. [Google Scholar] [CrossRef] [PubMed]
- Summer, E.J.; Liu, M. Application of Bacteriophages for the Control of Unwanted Bacteria in Biofuel Production Mediated by Bacterial Reactive Agents. U.S. Patents US13,423,010, 2012. [Google Scholar]
- Becker, S.C.; Foster-Frey, J.; Donovan, D.M. The phage K lytic enzyme LysK and lysostaphin act synergistically to kill MRSA. FEMS Microbiol. Lett. 2008, 287, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Khatibi, P.A.; Roach, D.R.; Donovan, D.M.; Hughes, S.R.; Bischoff, K.M. Saccharomyces cerevisiae expressing bacteriophage endolysins reduce Lactobacillus contamination during fermentation. Biotechnol. Biofuels 2014, 7, 104. [Google Scholar] [CrossRef]
- Van Tassell, M.L.; Angela Daum, M.; Kim, J.-S.; Miller, M.J. Creative lysins: Listeria and the engineering of antimicrobial enzymes. Curr. Opin. Biotechnol. 2016, 37, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Ribelles, P.; Rodríguez, I.; Suárez, J.E. LysA2, the Lactobacillus casei bacteriophage A2 lysin is an endopeptidase active on a wide spectrum of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2012, 94, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Roach, D.R.; Khatibi, P.A.; Bischoff, K.M.; Hughes, S.R.; Donovan, D.M. Bacteriophage-encoded lytic enzymes control growth of contaminating Lactobacillus found in fuel ethanol fermentations. Biotechnol. Biofuels 2013, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.C.; Schmelcher, M.; Rodriguez-Rubio, L.; Klumpp, J.; Pritchard, D.G.; Dong, S.; Donovan, D.M. Endolysins as antimicrobials. Adv. Virus Res. 2012, 83, 299–365. [Google Scholar] [PubMed]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.; de los Reyes-Gavilan, C.G.; Caso, J.L.; Suarez, J.E. Characterization of o393-A2, a bacteriophage that infects Lactobacillus casei. Microbiology 1994, 140, 2585–2590. [Google Scholar] [CrossRef]
- Parekh, R.N.; Shaw, M.R.; Wittrup, K.D. An Integrating Vector for Tunable, High Copy, Stable Integration into the Dispersed Ty δ Sites of Saccharomyces cerevisiae. Biotechnol. Prog. 1996, 12, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Gietz, R.D.; Schiestl, R.H. Frozen competent yeast cells that can be transformed with high efficiency using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.S.; Alper, H.; Yang, Y.T.; Stephanopoulos, G. Improvement of Xylose Uptake and Ethanol Production in Recombinant Saccharomyces cerevisiae through an Inverse Metabolic Engineering Approach. Appl. Environ. Microbiol. 2005, 71, 8249–8256. [Google Scholar] [CrossRef] [PubMed]
- Della-Bianca, B.E.; Gombert, A.K. Stress tolerance and growth physiology of yeast strains from the Brazilian fuel ethanol industry. Antonie Leeuwenhoek 2013, 104, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Gombert, A.K.; van Maris, A.J.A. Improving conversion yield of fermentable sugars into fuel ethanol in 1st generation yeast-based production processes. Curr. Opin. Biotechnol. 2015, 33, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.L.; Duarte, W.F.; Freire, A.L.; Dias, D.R.; Eleutherio, E.C.A.; Schwan, R.F. Evaluation of stress tolerance and fermentative behavior of indigenous Saccharomyces cerevisiae. Braz. J. Microbiol. 2013, 44, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Turchi, B.; Mok, K.C.; Taga, M.E.; Miller, M.J. HM2-phage resistant solventogenic Clostridium saccharoperbutylacetonicum N1-4 shows increased exopolysaccharide production. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed]
- Gaeng, S.; Scherer, S.; Neve, H.; Loessner, M.J. Gene cloning and expression and secretion of Listeria monocytogenes bacteriophage-lytic enzymes in Lactococcus lactis. Appl. Environ. Microbiol. 2000, 66, 2951–2958. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, T.; Lo Curto, R.; Minniti, E.; Narbad, A.; Mayer, M.J. Application of Lactobacillus johnsonii expressing phage endolysin for control of Clostridium perfringens. Lett. Appl. Microbiol. 2014, 59, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Briers, Y.; Lavigne, R. Volckaert G Antimicrobial Agents. U.S. Patents US13061053, 2009. [Google Scholar]
- Damasceno, L.M.; Huang, C., Jr.; Batt, C.A. Protein secretion in Pichia pastoris and advances in protein production. Appl. Microbiol. Biotechnol. 2012, 93, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Schleifer, K.H.; Kandler, O. Peptidoglycan types of bacterial cell walls and their taxonomic implications. Bacteriol. Rev. 1972, 36, 407–477. [Google Scholar] [PubMed]
- Sleytr, U. Bacterial S-layers. Trends Microbiol. 1999, 7, 253–260. [Google Scholar] [CrossRef]
- Hynönen, U.; Palva, A. Lactobacillus surface layer proteins: Structure, function and applications. Appl. Microbiol. Biotechnol. 2013, 97, 5225–5243. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.C.; Swift, S.; Korobova, O.; Schischkova, N.; Kopylov, P.; Donovan, D.M.; Abaev, I. Lytic activity of the staphylolytic Twort phage endolysin CHAP domain is enhanced by the SH3b cell wall binding domain. FEMS Microbiol. Lett. 2015, 362, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Eugster, M.R.; Haug, M.C.; Huwiler, S.G.; Loessner, M.J. The cell wall binding domain of Listeria bacteriophage endolysin PlyP35 recognizes terminal GlcNAc residues in cell wall teichoic acid: GlcNAc is the binding ligand for Listeria phage endolysin PlyP35. Mol. Microbiol. 2011, 81, 1419–1432. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Dorado, I.; Campillo, N.E.; Monterroso, B.; Hesek, D.; Lee, M.; Páez, J.A.; García, P.; Martínez-Ripoll, M.; García, J.L.; Mobashery, S.; et al. Elucidation of the Molecular Recognition of Bacterial Cell Wall by Modular Pneumococcal Phage Endolysin CPL-1. J. Biol. Chem. 2007, 282, 24990–24999. [Google Scholar] [CrossRef] [PubMed]
- Cereghino, G.P.L.; Cereghino, J.L.; Ilgen, C.; Cregg, J.M. Production of recombinant proteins in fermenter cultures of the yeast Pichia pastoris. Curr. Opin. Biotechnol. 2002, 13, 329–332. [Google Scholar] [CrossRef]
- Narendranath, N.V.; Hynes, S.H.; Thomas, K.C.; Ingledew, W.M. Effects of lactobacilli on yeast-catalyzed ethanol fermentations. Appl. Environ. Microbiol. 1997, 63, 4158–4163. [Google Scholar] [PubMed]
- De Oliva-Neto, P.; Yokoya, F. Evaluation of bacterial contamination in a fed-batch alcoholic fermentation process. World J. Microbiol. Biotechnol. 1994, 10, 697–699. [Google Scholar] [CrossRef] [PubMed]
- Gellissen, G.; Strasser, A.W.M.; Suckow, M. Key and Criteria to the Selection of An Expression Platform. In Production of Recombinant Proteins; Gellissen, G., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2005; pp. 1–5. [Google Scholar]
- Idiris, A.; Tohda, H.; Kumagai, H.; Takegawa, K. Engineering of protein secretion in yeast: Strategies and impact on protein production. Appl. Microbiol. Biotechnol. 2010, 86, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Abreu-Cavalheiro, A.; Monteiro, G. Solving ethanol production problems with genetically modified yeast strains. Braz. J. Microbiol. 2013, 44, 665–671. [Google Scholar] [CrossRef] [PubMed]
- De Souza, R.B.; de Menezes, J.A.S.; de Souza, R.D.F.R.; Dutra, E.D.; de Morais, M.A. Mineral composition of the sugarcane juice and its influence on the ethanol fermentation. Appl. Biochem. Biotechnol. 2015, 175, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Argueso, J.L.; Carazzolle, M.F.; Mieczkowski, P.A.; Duarte, F.M.; Netto, O.V.C.; Missawa, S.K.; Galzerani, F.; Costa, G.G.L.; Vidal, R.O.; Noronha, M.F.; et al. Genome structure of a Saccharomyces cerevisiae strain widely used in bioethanol production. Genome Res. 2009, 19, 2258–2270. [Google Scholar] [CrossRef] [PubMed]
- Basso, L.C.; de Amorim, H.V.; de Oliveira, A.J.; Lopes, M.L. Yeast selection for fuel ethanol production in Brazil. FEMS Yeast Res. 2008, 8, 1155–1163. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Description |
|---|---|
| Escherichia coli | BL21 (DE3) |
| Escherichia coli | BL21 (DE3) containing pRSETA_LysA2 |
| Pichia pastoris | GS115 |
| Pichia pastoris | GS115 containing pPICZAa |
| Pichia pastoris | GS115 containing pPICZA_LysA2 |
| Pichia pastoris | GS115 containing pPICZAa_LysA2 |
| Saccharomyces cerevisiae | D452-2 |
| Saccharomyces cerevisiae | D452-2 containing pRS423_LysA2 |
| Saccharomyces cerevisiae | D452-2 containing pITY3_LysA2 1 |
| Saccharomyces cerevisiae | D452-2 empty vector |
| Bacterial Contaminants | ATCC # | Peptidoglycan Chemotype | Relative Activity 1 |
|---|---|---|---|
| Aerococcus viridans | 11563 | A1α direct | − |
| Lactobacillus plantarum | 14917 | A1γ mesoDpm | − |
| Lactobacillus casei | 393 | A4α L-Lys-D-Asp | Positive control 2 |
| Enterococcus faecium | 6057 | A4α L-Lys-D-Asp | ++ |
| Enterococcus gallinarum | 49573 | A4α L-Lys-D-Asp | ++ |
| Lactobacillus brevis | 14869 | A4α L-Lys-D-Asp | − |
| Lactobacillus delbrueckii | 9649 | A4α L-Lys-D-Asp | +++ |
| Lactobacillus paracasei | 25598 | A4α L-Lys-D-Asp | ++++ |
| Lactobacillus rhamnosus | 53103 | A4α L-Lys-D-Asp | + |
| Lactococcus lactis | 19257 | A4α L-Lys-D-Asp | + |
| Pediococcus acidilactici | NA | A4α L-Lys-D-Asp | − |
| Pediococcus damnosus | 29358 | A4α L-Lys-D-Asp | − |
| Lactobacillus fermentum | 9338 | A4β L-Orn-D-Asp | ++ |
| Contaminants | Yeast Strain | n | Glucose Consumption (g/L ± SEM) | Lactic Acid (g/L ± SEM) | Ethanol (g/L ± SEM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 6 h | 12 h | 24 h | 0 h | 6 h | 12 h | 24 h | 0 h | 6 h | 12 h | 24 h | |||
| None | SC-EV 1 | 3 | 0.0 ± 0.0 | 2.6 ± 0.3 | 13.7 ± 0.3 | 62 ± 0.4 | ND | ND | ND | ND | 0.0 ± 0.0 | 0.6 ± 0.1 | 6.1 ± 0.1 | 26.8 ± 1.8 |
| SC-LysA2s 2 | 3 | 0.0 ± 0.0 | 3.2 ± 0.0 | 15.1 ± 0.5 | 62 ± 0.0 | ND | ND | ND | ND | 0.0 ± 0.0 | 0.8 ± 0.3 | 8.0 ± 0.2 | 27.3 ± 0.2 | |
| L. fermentum | SC-EV 1 | 3 | 0.0 ± 0.0 | 4.7 ± 0.0 | 17.1 ± 0.3 | 50.6 ± 0.2 | 0.0 ± 0.0 | 0.7 ± 0.0 | 1.8 ± 0.0 | 3.4 ± 0.0 | 0.0 ± 0.0 | 0.8 ± 0.0 | 6.2 ± 0.3 | 20.8 ± 0.1 |
| SC-LysA2s 2 | 3 | 0.0 ± 0.0 | 5.1 ± 0.8 | 19.3 ± 0.4 | 62.0 ± 0.0 | 0.0 ± 0.0 | 0.6 ± 0.0 | 1.2 ± 0.0 | 2.5 ± 0.0 | 0.0 ± 0.0 | 1.1 ± 0.3 | 7.1 ± 0.2 | 26.8 ± 0.2 | |
| Contaminant | Yeast Strain | n | Ethanol Production (g/L ± SEM) | Lactic Acid (g/L ± SEM) | Ethanol yield (g/g ± SEM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 12 h | 24 h | 48 h | 0 h | 12 h | 24 h | 48 h | 0 h | 12 h | 24 h | 48 h | |||
| None | SC-EV 1 | 3 | 0.0 ± 0.0 | 6.8 ± 0.5 | 15.5 ± 0.5 | 28.4 ± 0.1 | ND | ND | ND | ND | 0 | 0.46 | 0.48 | 0.48 |
| SC-LysA2s 2 | 3 | 0.0 ± 0.0 | 6.9 ± 0.1 | 15.4 ± 0.2 | 27.8 ± 0.0 | ND | ND | ND | ND | 0 | 0.45 | 0.48 | 0.48 | |
| SC-LysA2s* 3 | 3 | 0.0 ± 0.0 | 6.9 ± 0.1 | 16.4 ± 0.3 | 28.8 ± 0.2 | ND | ND | ND | ND | 0 | 0.45 | 0.48 | 0.48 | |
| SC-EV/LysA2-PI 4 | 3 | 0.0 ± 0.0 | 6.7 ± 0.3 | 16.2 ± 0.2 | 27.8 ± 0.2 | ND | ND | ND | ND | 0 | 0.46 | 0.48 | 0.48 | |
| L. fermentum | SC-EV 1 | 3 | 0.0 ± 0.0 | 6.4 ± 0.1 | 13.4 ± 0.4 | 18.6 ± 0.4 | 0.0 ± 0.0 | 1.1 ± 0.0 | 2.8 ± 0.0 | 3.5 ± 0.0 | 0 | 0.25 | 0.27 | 0.31 |
| SC-LysA2s 2 | 3 | 0.0 ± 0.0 | 6.6 ± 0.2 | 13.2 ± 0.2 | 18.4 ± 0.0 | 0.0 ± 0.0 | 1.3 ± 0.0 | 2.6 ± 0.0 | 3.4 ± 0.0 | 0 | 0.24 | 0.27 | 0.32 | |
| SC-LysA2s* 3 | 3 | 0.0 ± 0.0 | 6.2 ± 0.2 | 15.5 ± 0.2 | 20.3 ± 0.0 | 0.0 ± 0.0 | 0.6 ± 0.0 | 2.2 ± 0.0 | 3.1 ± 0.0 | 0 | 0.24 | 0.30 | 0.35 | |
| SC-EV/LysA2-PI 4 | 3 | 0.0 ± 0.0 | 6.0 ± 0.2 | 18.7 ± 0.2 | 23.6 ± 0.3 | 0.0 ± 0.0 | 0.4 ± 0.0 | 1.2 ± 0.0 | 2.2 ± 0.0 | 0 | 0.41 | 0.39 | 0.4 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-S.; Daum, M.A.; Jin, Y.-S.; Miller, M.J. Yeast Derived LysA2 Can Control Bacterial Contamination in Ethanol Fermentation. Viruses 2018, 10, 281. https://doi.org/10.3390/v10060281
Kim J-S, Daum MA, Jin Y-S, Miller MJ. Yeast Derived LysA2 Can Control Bacterial Contamination in Ethanol Fermentation. Viruses. 2018; 10(6):281. https://doi.org/10.3390/v10060281
Chicago/Turabian StyleKim, Jun-Seob, M. Angela Daum, Yong-Su Jin, and Michael J. Miller. 2018. "Yeast Derived LysA2 Can Control Bacterial Contamination in Ethanol Fermentation" Viruses 10, no. 6: 281. https://doi.org/10.3390/v10060281
APA StyleKim, J.-S., Daum, M. A., Jin, Y.-S., & Miller, M. J. (2018). Yeast Derived LysA2 Can Control Bacterial Contamination in Ethanol Fermentation. Viruses, 10(6), 281. https://doi.org/10.3390/v10060281