A Novel Deltaflexivirus that Infects the Plant Fungal Pathogen, Sclerotinia sclerotiorum, Can Be Transmitted Among Host Vegetative Incompatible Strains


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture
2.2. Analysis of the Biological Properties of Strain 228
2.3. Extraction and Purification of dsRNA
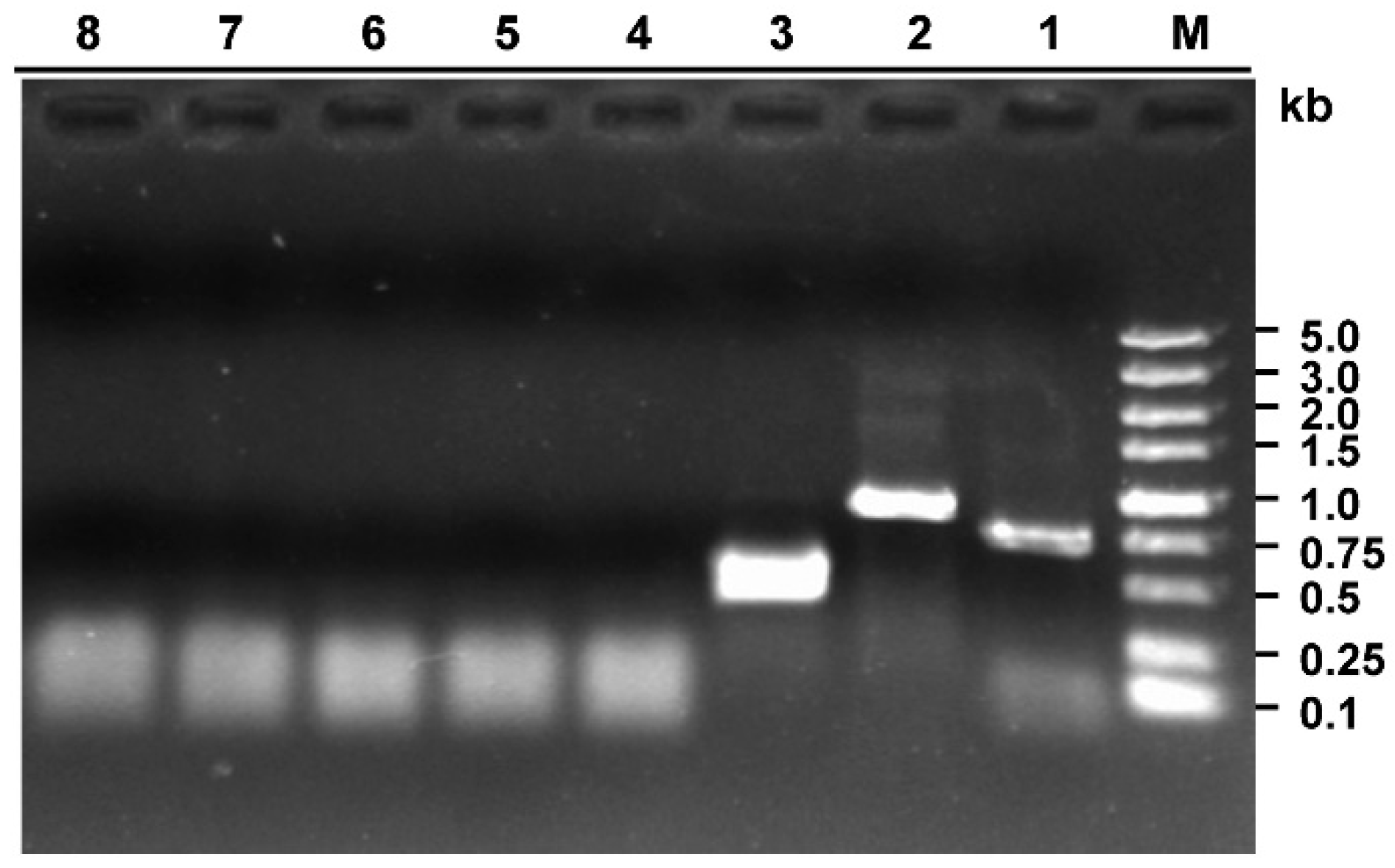
2.4. Confirmation of Putative Mycoviruses
2.5. Rapid Amplification of cDNA Ends (RACE) and Sequencing
2.6. Bioinformatics and Sequence Analysis
2.7. Viral Transmission Assay
3. Results
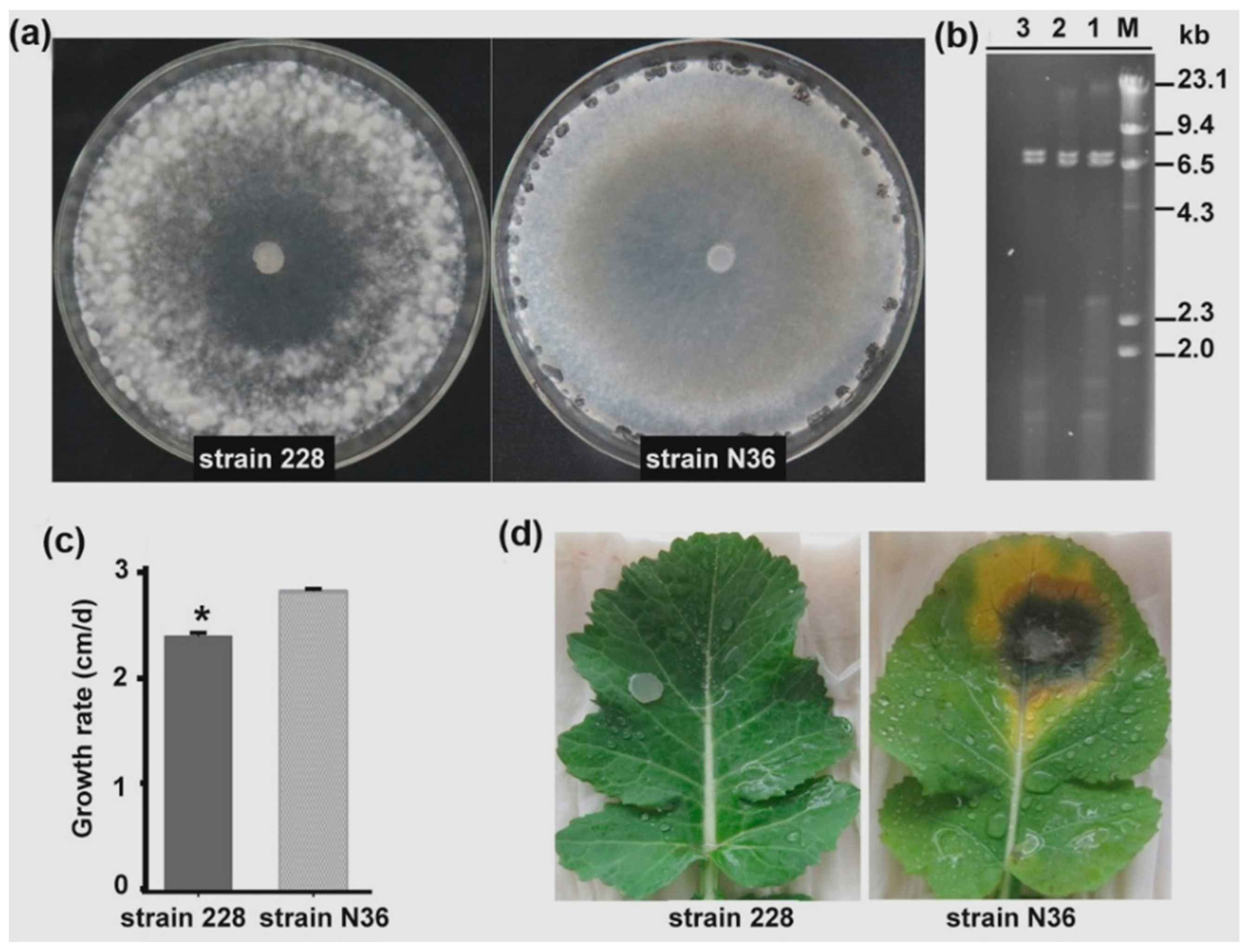
3.1. Strain 228 is a Hypovirulent Strain of S. sclerotiorum Co-Infected by Two Viruses
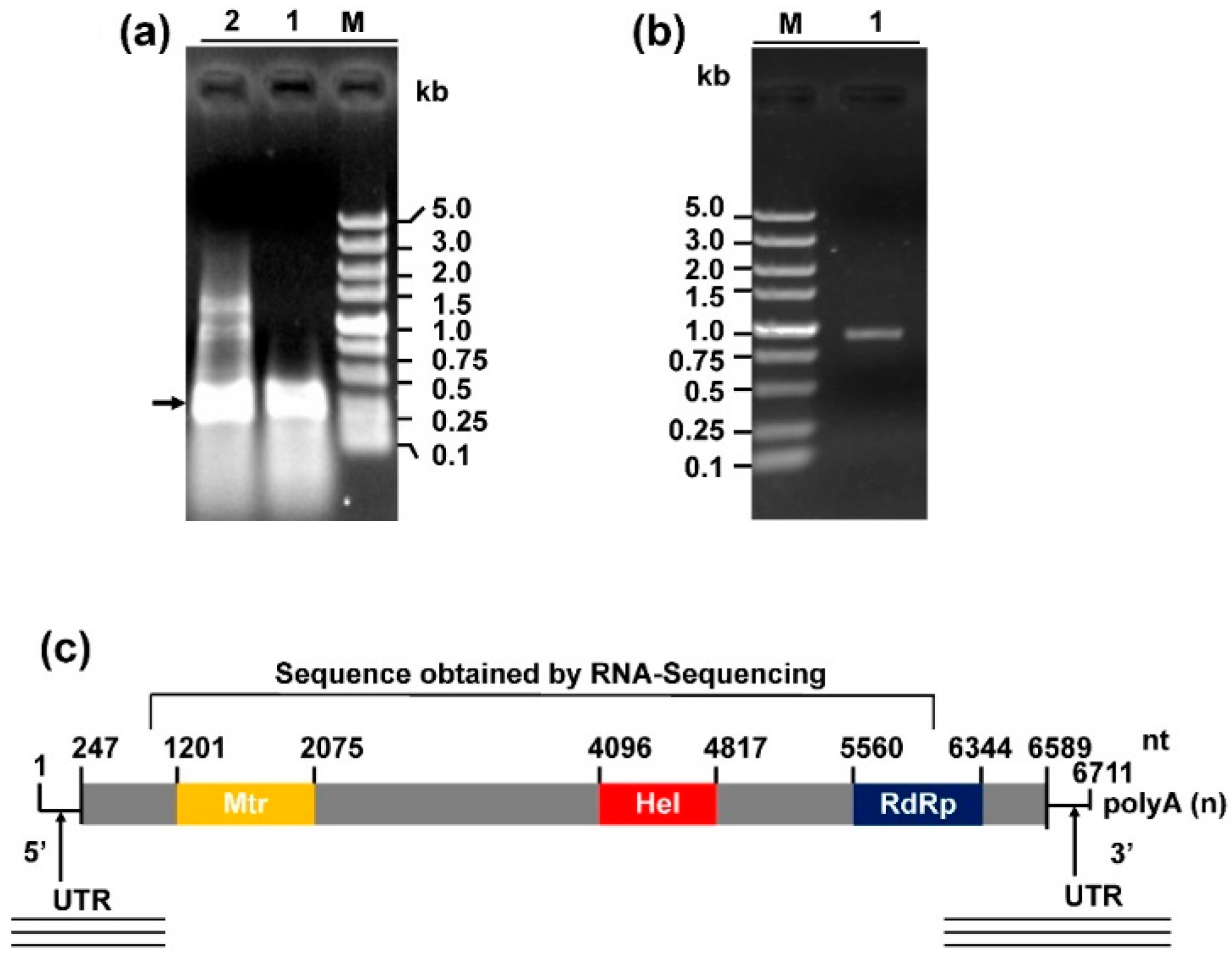
3.2. Genome Organization of Sclerotinia sclerotiorum Deltaflexivirus 2
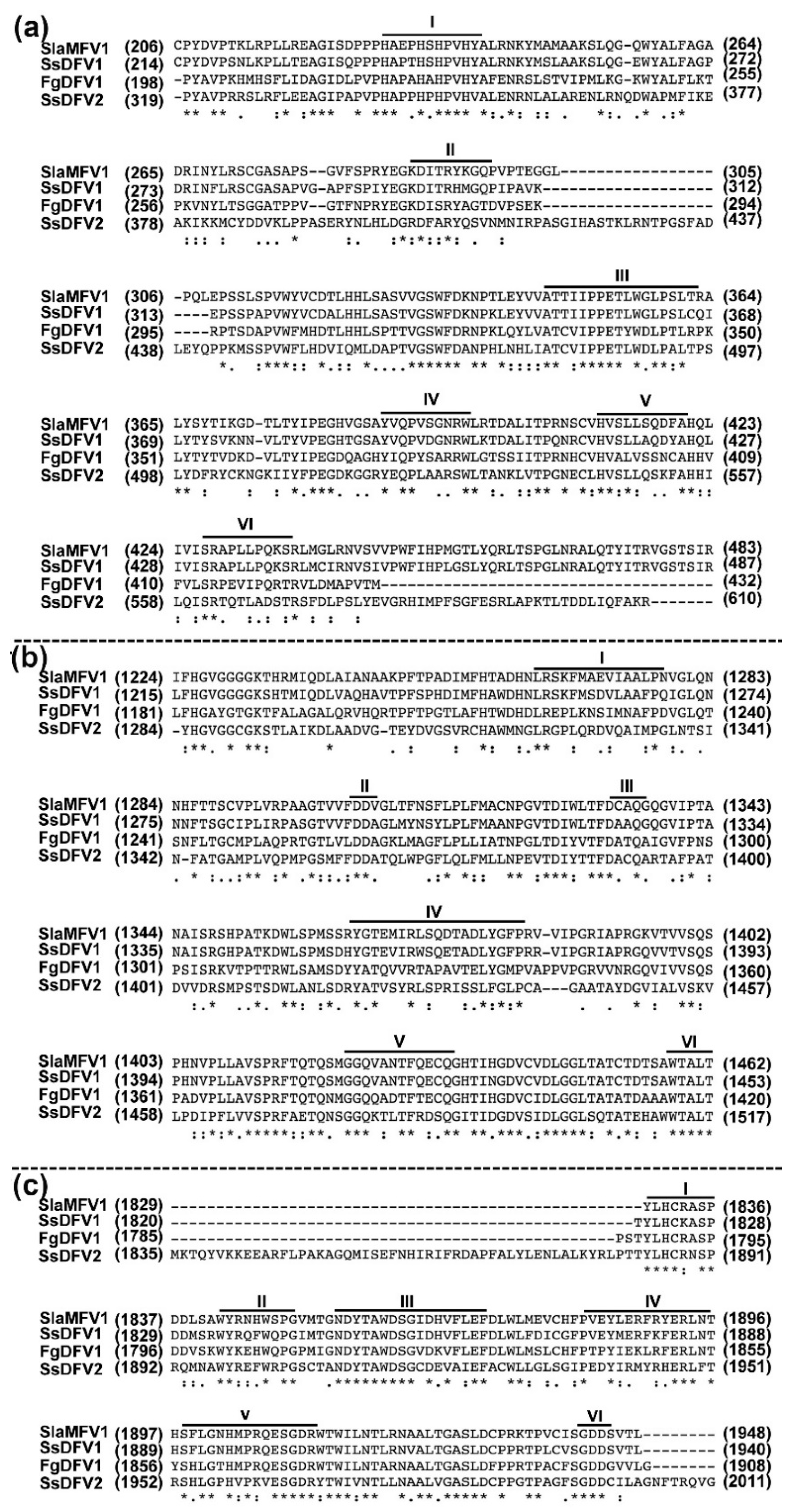
3.3. SsDFV2 is Closely Phylogenetically Related to Flexiviruses
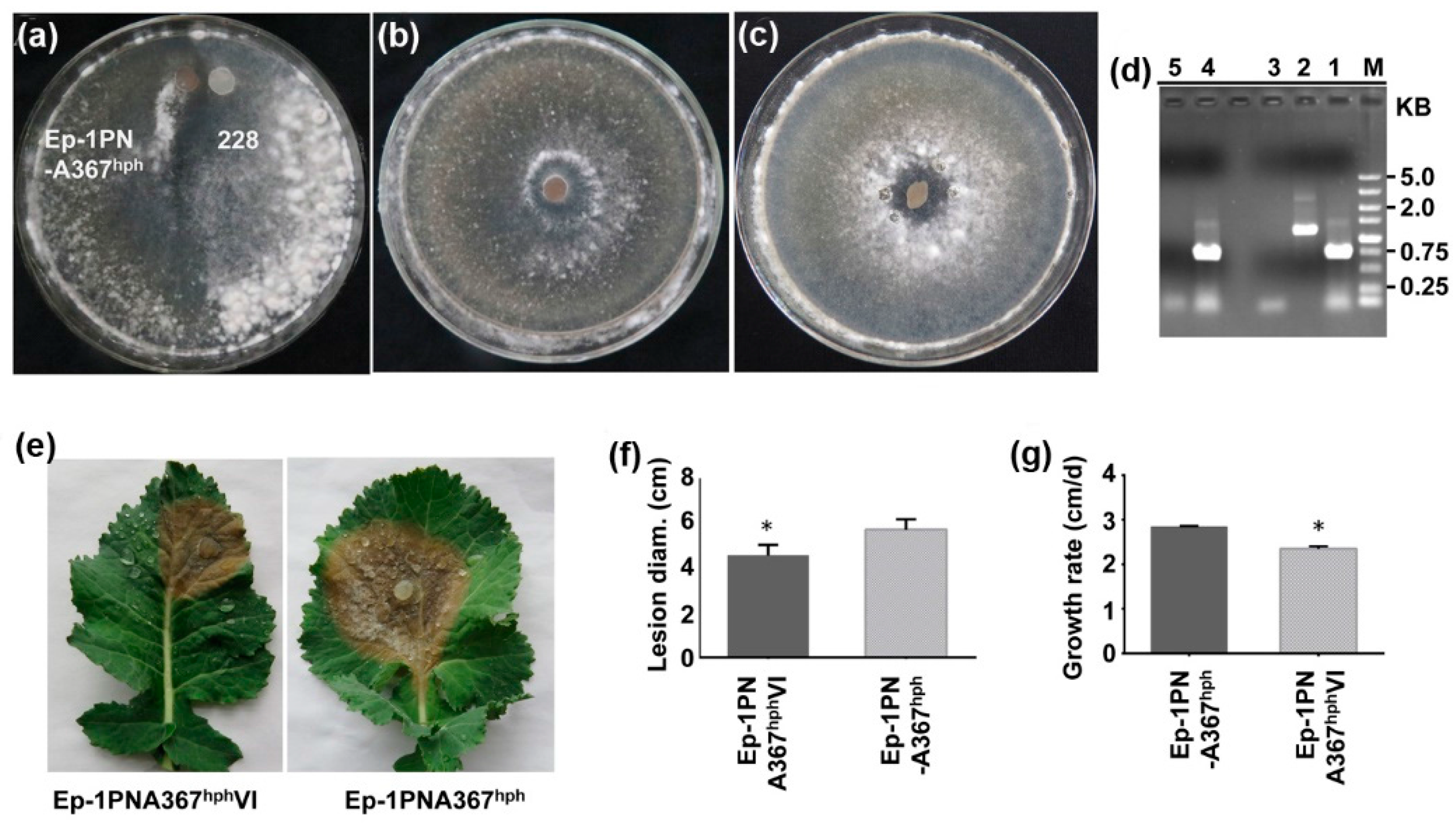
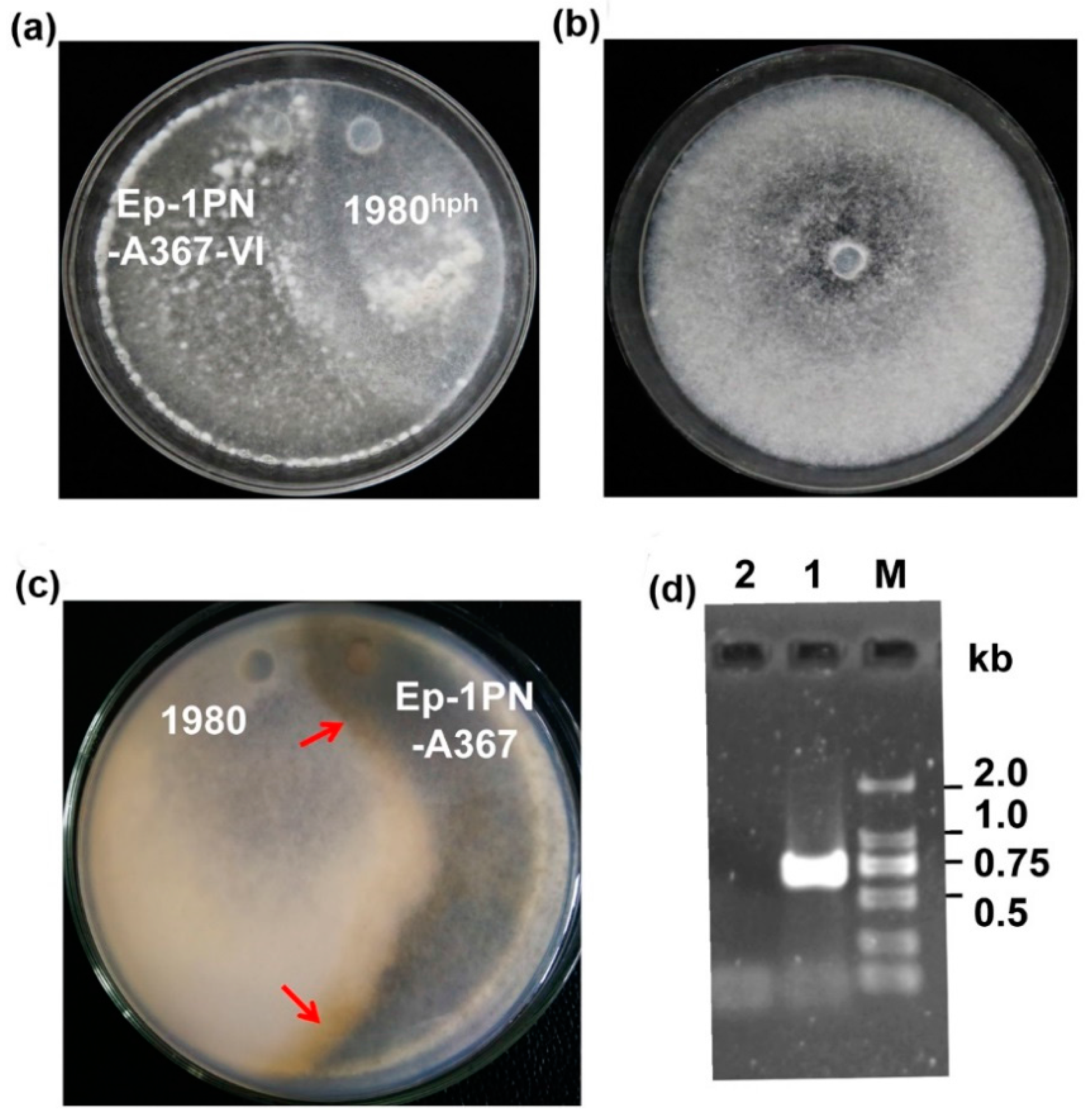
3.4. SsDFV2 can be Transmitted Among Individuals that Exhibit Vegetative Incompatibility with the Host
3.5. SsDFV2 Had a Minor Influence on the S. sclerotiorum Phenotype
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bolton, M.D.; Thomma, B.P.; Nelson, B.D. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant. Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal–plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [PubMed]
- Cortesi, P.; Mcculloch, C.E.; Song, H.; Lin, H.; Milgroom, M.G. Genetic control of horizontal virus transmission in the chestnut blight fungus, Cryphonectria parasitica. Genetics 2001, 159, 107–118. [Google Scholar] [PubMed]
- Milgroom, M.G.; Cortesi, P. Biological control of chestnut blight with hypovirulence: A critical analysis. Annu. Rev. Phytopathol. 2004, 42, 311–338. [Google Scholar] [CrossRef] [PubMed]
- Brusini, J.; Robin, C. Mycovirus transmission revisited by in situ pairings of vegetatively incompatible isolates of Cryphonectria parasitica. J. Virol. Methods 2013, 187, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.X.; Nuss, D.L. Engineering super mycovirus donor strains of chestnut blight fungus by systematic disruption of multilocus vic genes. Proc. Natl. Acad. Sci. USA 2016, 113, 2062–2067. [Google Scholar] [CrossRef] [PubMed]
- Rigling, D.; Prospero, S. Cryphonectria parasitica, the causal agent of chestnut blight: Invasion history, population biology and disease control. Mol. Plant. Pathol. 2018, 19, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, B.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Extracellular transmission of a DNA mycovirus and its use as a natural fungicide. Proc. Natl. Acad. Sci. USA 2013, 110, 1452–1457. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xie, J.; Cheng, J.; Li, B.; Chen, T.; Fu, Y.; Li, G.; Wang, M.; Jin, H.; Wan, H.; et al. Fungal DNA virus infects a mycophagous insect and utilizes it as a transmission vector. Proc. Natl. Acad. Sci. USA 2016, 113, 12803–12808. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Ghabrial, S.A.; Xie, J. Virus-mediated suppression of host non-self recognition facilitates horizontal transmission of heterologous viruses. PLoS Pathogens 2017, 13, e1006234. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Xiao, X.; Fu, Y.; Liu, H.; Cheng, J.; Ghabrial, S.A.; Li, G.; Jiang, D. A novel mycovirus closely related to hypoviruses that infects the plant pathogenic fungus Sclerotinia sclerotiorum. Virology 2011, 418, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, M.E.; Pearson, M.N. Molecular characterisation of novel mitoviruses associated with Sclerotinia sclerotiorum. Arch. Virol. 2014, 159, 3157–3160. [Google Scholar] [CrossRef] [PubMed]
- Marzano, S.Y.L.; Nelson, B.D.; Ajayi-Oyetunde, O.; Bradley, C.A.; Hughes, T.J.; Hartman, G.L.; Eastburn, D.M.; Domier, L.L. Identification of diverse mycoviruses through metatranscriptomics characterization of the viromes of five major fungal plant pathogens. J. Virol. 2016, 90, 6846–6863. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wu, S.; Liu, L.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. A mitovirus related to plant mitochondrial gene confers hypovirulence on the phytopathogenic fungus Sclerotinia sclerotiorum. Virus Res. 2015, 197, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, Q.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. Molecular characterization of a bipartite double-stranded RNA virus and its satellite-like RNA co-infecting the phytopathogenic fungus Sclerotinia sclerotiorum. Front. Microbiol. 2015, 6, 406. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zheng, D.; Cheng, J.; Chen, T.; Fu, Y.; Jiang, D.; Xie, J. Characterization of a novel Sclerotinia sclerotiorum RNA virus as the prototype of a new proposed family within the order Tymovirales. Virus Res. 2016, 219, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wan, Y.; Sun, X.; Cheng, J.; Fu, Y.; Liu, H.; Jiang, D.; Ghabrial, S.A.; Xie, J. Characterization of a novel megabirnavirus from Sclerotinia sclerotiorum reveals horizontal gene transfer from single-stranded RNA virus to double-stranded RNA virus. J. Virol. 2015, 89, 8567–8579. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, M.E.; Pearson, M.N. Characterisation of a novel hypovirus from Sclerotinia sclerotiorum potentially representing a new genus within the Hypoviridae. Virology 2014, 464, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Marzano, S.Y.; Hobbs, H.A.; Nelson, B.D.; Hartman, G.L.; Eastburn, D.M.; Mccoppin, N.K.; Domier, L.L. Transfection of Sclerotinia sclerotiorum with in vitro transcripts of a naturally occurring interspecific recombinant of Sclerotinia sclerotiorum hypovirus 2 significantly reduces virulence of the fungus. J. Virol. 2015, 89, 5060–5071. [Google Scholar] [CrossRef] [PubMed]
- Mu, F.; Xie, J.; Cheng, S.; You, M.; Barbetti, M.J.; Jia, J.; Wang, Q.; Cheng, J.; Fu, Y.; Chen, T. Virome Characterization of a Collection of S. sclerotiorum from Australia. Front. Microbiol. 2017, 8, 2540. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; He, H.; Yang, X.; Zeng, H.; Qiu, D.; Guo, L. The complete genome sequence of a novel Fusarium graminearum RNA virus in a new proposed family within the order Tymovirales. Arch. Virol. 2016, 161, 2899–2903. [Google Scholar] [CrossRef] [PubMed]
- Marzano, S.Y.L.; Domier, L.L. Novel mycoviruses discovered from metatranscriptomics survey of soybean phyllosphere phytobiomes. Virus Res. 2016, 213, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Fu, Y.; Xie, J.; Jiang, D.; Li, G.; Yi, X. A novel virus that infecting hypovirulent strain XG36-1 of plant fungal pathogen Sclerotinia sclerotiorum. Virol. J. 2009, 6, 96. [Google Scholar] [CrossRef] [PubMed]
- Rong, L.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. Molecular characterization of a novel positive-sense, single-stranded RNA mycovirus infecting the plant pathogenic fungus Sclerotinia sclerotiorum. Viruses 2015, 7, 2470–2484. [Google Scholar]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Darissa, O.; Willingmann, P.; Adam, G. Optimized approaches for the sequence determination of double-stranded RNA templates. J. Virol. Methods 2010, 169, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acid Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acid Res. 2018, 46, D708. [Google Scholar] [CrossRef] [PubMed]
- Howitt, R.L.J.; Beever, R.E.; Pearson, M.N.; Forster, R.L.S. Genome characterization of a flexuous rod-shaped mycovirus, Botrytis virus X, reveals high amino acid identity to genes from plant ‘potex-like’ viruses. Arch. Virol. 2006, 151, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Howitt, R.L.; Beever, R.E.; Pearson, M.N.; Forster, R.L. Genome characterization of Botrytis virus F, a flexuous rod-shaped mycovirus resembling plant ‘potex-like’ viruses. J. Gen. Virol. 2001, 82, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wei, D.; Jiang, D.; Fu, Y.; Li, G.; Ghabrial, S.A.; Peng, Y. Characterization of debilitation-associated mycovirus infecting the plant-pathogen fungus Sclerotinia sclerotiorum. J. Gen. Virol. 2006, 87, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Mascia, T.; Nigro, F.; Abdallah, A.; Ferrara, M.; De Stradis, A.; Faedda, R.; Palukaitis, P.; Gallitelli, D. Gene silencing and gene expression in phytopathogenic fungi using a plant virus vector. Proc. Natl. Acad. Sci. USA 2014, 111, 4291–4296. [Google Scholar] [CrossRef] [PubMed]
- Nerva, L.; Varese, G.C.; Falk, B.W.; Turina, M. Mycoviruses of an endophytic fungus can replicate in plant cells: Evolutionary implications. Sci. Rep. 2017, 7, 1908. [Google Scholar] [CrossRef] [PubMed]
- Marais, A.; Nivault, A.; Faure, C.; Comont, G.; Theil, S.; Candresse, T.; Corio-Costet, M.-F. Molecular characterization of a novel fusarivirus infecting the plant-pathogenic fungus Neofusicoccum luteum. Arch. Virol. 2018, 163, 559–562. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Shang, H.H.; Zhu, C.X.; Zhu, J.Z.; Zhu, H.J.; Hu, Y.; Gao, B.D. Characterization of a novel single-stranded RNA virus, closely related to fusariviruses, infecting the plant pathogenic fungus Alternaria brassicicola. Virus Res. 2016, 217, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Zhao, S.Q.; Li, G.F.; Pang, X.D.; Deng, X.J.; Zhu, H.J.; Gao, B.D.; Zhou, Q. A novel fusarivirus isolated from the phytopathogenic fungus Nigrospora oryzae. Virus Genes 2016, 52, 891–895. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.J.; Lim, W.S.; Park, S.H.; Park, M.R.; Kim, K.H. Molecular characterization of a dsRNA mycovirus, Fusarium graminearum virus-DK21, which is phylogenetically related to hypoviruses but has a genome organization and gene expression strategy resembling those of plant potex-like viruses. Mol. Cells 2009, 28, 304–315. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Suzuki, N. Fungal Viruses. In Encyclopedia of Virology, 3rd ed.; Elsevier: Oxford, UK, 2008; pp. 284–291. [Google Scholar]
- Sasaki, A.; Miyanishi, M.; Ozaki, K.; Onoue, M.; Yoshida, K. Molecular characterization of a partitivirus from the plant pathogenic ascomycete Rosellinia necatrix. Arch. Virol. 2005, 150, 1069–1083. [Google Scholar] [CrossRef] [PubMed]
- Cole, T.; Mcller, B.; Hong, Y.; Brasier, C.; Buck, K. Complexity of virus-like double-stranded RNA elements in a diseased isolate of the Dutch elm disease fungus, Ophiostoma novo-ulmi. J. Phytopathol. 1998, 146, 593–598. [Google Scholar] [CrossRef]
- Hong, Y.; Cole, T.E.; Brasier, C.M.; Buck, K.W. Evolutionary relationships among putative RNA-dependent RNA polymerases encoded by a mitochondrial virus-like RNA in the Dutch elm disease fungus, Ophiostoma novo-ulmi, by other viruses and virus-like RNAs and by the Arabidopsis mitochondrial genome. Virology 1998, 246, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, M.E.; Pearson, M.N. Molecular characterization of three mitoviruses co-infecting a hypovirulent isolate of Sclerotinia sclerotiorum fungus. Virology 2013, 441, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Fu, Y.; Li, G.; Ghabrial, S.A. Viruses of the plant pathogenic fungus Sclerotinia sclerotiorum. Adv. Virus Res. 2013, 86, 215–248. [Google Scholar] [PubMed]
- Potgieter, C.A.; Castillo, A.; Castro, M.; Cottet, L.; Morales, A. A wild-type Botrytis cinerea strain co-infected by double-stranded RNA mycoviruses presents hypovirulence-associated traits. Virol. J. 2013, 10, 220. [Google Scholar] [CrossRef] [PubMed]
- Zanzinger, D.H.; Bandy, B.P.; Tavantzis, S.M. High frequency of finding double-stranded RNA in naturally occurring isolates of Rhizoctonia solani. J. Gen. Virol. 1984, 65, 1601–1605. [Google Scholar] [CrossRef]
- Das, S. Rhizoctonia solani on Potato in New Zealand: Pathogen Characterisation and Identification of Double-Stranded RNA Viruses that May Affect Their Virulence; Lincoln University: Canterbury, New Zealand, 2013. [Google Scholar]
- Bartholomäus, A.; Wibberg, D.; Winkler, A.; Pühler, A.; Schlüter, A.; Varrelmann, M. Deep sequencing analysis reveals the mycoviral diversity of the virome of an avirulent isolate of Rhizoctonia solani AG-2-2 IV. PLoS ONE 2016, 11, e0165965. [Google Scholar] [CrossRef] [PubMed]
- Lyu, R.; Yi, Z.; Tang, Q.; Li, Y.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. Two alphapartitiviruses co-infecting a single isolate of the plant pathogenic fungus Rhizoctonia solani. Arch. Virol. 2017, 163, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Choi, G.H.; Nuss, D.L. A single argonaute gene is required for induction of RNA silencing antiviral defense and promotes viral RNA recombination. Proc. Natl. Acad. Sci. USA 2009, 106, 17927–17932. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Nuss, D.L. A host dicer is required for defective viral RNA production and recombinant virus vector RNA instability for a positive sense RNA virus. Proc. Natl. Acad. Sci. USA 2008, 105, 16749–16754. [Google Scholar] [CrossRef] [PubMed]
- Segers, G.C.; Zhang, X.; Deng, F.; Sun, Q.; Nuss, D.L. Evidence that RNA silencing functions as an antiviral defense mechanism in fungi. Proc. Natl. Acad. Sci. USA 2007, 104, 12902–12906. [Google Scholar] [CrossRef] [PubMed]
- Andika, I.B.; Jamal, A.; Kondo, H.; Suzuki, N. SAGA complex mediates the transcriptional up-regulation of antiviral RNA silencing. Proc. Natl. Acad. Sci. USA 2017, 114, E3499. [Google Scholar] [CrossRef] [PubMed]
- Chiba, S.; Suzuki, N. Highly activated RNA silencing via strong induction of dicer by one virus can interfere with the replication of an unrelated virus. Proc. Natl. Acad. Sci. USA 2015, 112, 4911–4918. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Hisano, S.; Tani, A.; Kondo, H.; Kanematsu, S.; Suzuki, N. A capsidless ssRNA virus hosted by an unrelated dsRNA virus. Nat. Microbiol. 2016, 1, 15001. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Cheng, J.; Tang, J.; Fu, Y.; Jiang, D.; Baker, T.S.; Ghabrial, S.A.; Xie, J. A novel partitivirus that confers hypovirulence on plant pathogenic fungi. J. Virol. 2014, 88, 10120–10133. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence |
|---|---|
| SsDFV2-R1 | 5′-GGGAGGCGATTGACAGG-3′ |
| SsDFV2-F1 | 5′-TGTCTTTGGCAGTACCCTCC-3′ |
| BpRV1-dsRNA1-R1 | 5′-AGCCAGGTATGCGTGAATG-3′ |
| BpRV1-dsRNA1-F1 | 5′-CAAGGGTAAAGGAAGCGACT-3′ |
| BpRV1-dsRNA2-R1 | 5′-GGCCTCATCCCAGATTAAG-3′ |
| BpRV1-dsRNA2-F1 | 5′-CTGTCTACTCGGGCTCAAGG-3′ |
| PC3-T7 Loop adapter | 5′-p-GGATCCCGGGAATTCGGTAATACGACTCACTATATTTT TATAGTGAGTCGTATTA-OH-3′ |
| PC2 primer | 5′-p-CCGAATTCCCGGGATCC-3′ |
| Seq. specific primer—5end SsDFV2-5end-R | 5′-CGGGATCATCAATGCGAC-3′ |
| Seq specific primer—3 end SsDFV2-3end-F | 5′-TTCAACCACATCCGAATCTT-3′ |
| Family | Virus Name | Abbr. | NCBI Accession Number | Mtr (Identity %) | Hel (Identity %) | RdRp (Identity %) |
|---|---|---|---|---|---|---|
| Alphaflexiviridae | Potato Virus X | PVX | NC_011620.1 | 34/205 (17%) | 59/238 (25%) | 54/237 (23%) |
| Yam Virus X | YVX | NC_025252.1 | 54/263 (20%) | 60/235 (26%) | 53/225 (24%) | |
| Cassava virus X | CVX | NC_034375.1 | 75/296 (25%) | 25/156 (16%) | 37/225 (16%) | |
| Sclerotinia sclerotiorum debilitation-associated RNA virus | SsDRV | NC_007415.1 | 72/263 (27%) | 25/108 (23%) | 46/201 (23%) | |
| Tymoviridae | Okra Mosaic Virus | OMV | NC_009532.1 | 75/244 (31%) | - | 29/129 (22%) |
| Grapevine rupestris vein feathering virus | GRVFV | KY513702.1 | 87/294 (30%) | 32/198 (16%) | 45/202 (22%) | |
| Grapevine Syrah Virus 1 | GSV | JX513896.1 | 82/294 (28%) | 27/191 (14%) | 26/146 (18%) | |
| Gammaflexi-viridae | Botrytis Virus F | BVF | NC_002604.1 | 60/177 (34%) | 40/235 (17%) | 61/218 (28%) |
| Betaflexiviridae | Citrus leaf blotch virus | CLBV | KR023647.1 | 44/291 (15%) | 24/132 (18%) | 75/251 (30%) |
| Cherry rusty mottle associated virus | CRMAV | KP258176.1 | 32/243 (13%) | 34/238 (14%) | 73/250 (29%) | |
| Shallot latent virus | SLV | LC279526.1 | 9/264 (30%) | 20/144 (14%) | 53/191 (28%) | |
| Garlic common latent virus | GCLV | KX255694.1 | 49/291 (17%) | 81/257 (32%) | 64/262 (24%) | |
| Blueberry scorch virus | BSV | AY941199.1 | 75/258 (29%) | 30/224 (13%) | 77/254 (30%) | |
| Cowpea mild mottle virus | CMMV | NC_014730.1 | 73/249 (29%) | - | 75/254 (30%) | |
| Deltaflexiviridae | Soybean leaf-associated mycoflexivirus 1 | SlaMFV1 | KT598226.1 | 110/292 (38%) | 101/243 (42%) | 122/262 (47%) |
| Fusarium graminearum deltaflexivirus 1 | FgDFV1 | NC_030654.1 | 111/292 (38%) | 113/247 (46%) | 134/262 (51%) | |
| Sclerotinia sclerotiorum deltaflexivirus 1 | SsDFV1 | KT581451.1 | 101/292 (35%) | 105/244 (43%) | 132/262 (50%) | |
| Unlcassified Flexivirus | Rhizoctonia solani flexivirus 1 | RsFV1 | NC_030655.1 | 87/234 (37%) | 71/157 (45%) | 58/196 (29%) |
| Rhizoctonia solani flexivirus 2 | RsFV2 | KX349069.1 | - | - | 97/262 (37%) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamid, M.R.; Xie, J.; Wu, S.; Maria, S.K.; Zheng, D.; Assane Hamidou, A.; Wang, Q.; Cheng, J.; Fu, Y.; Jiang, D. A Novel Deltaflexivirus that Infects the Plant Fungal Pathogen, Sclerotinia sclerotiorum, Can Be Transmitted Among Host Vegetative Incompatible Strains. Viruses 2018, 10, 295. https://doi.org/10.3390/v10060295
Hamid MR, Xie J, Wu S, Maria SK, Zheng D, Assane Hamidou A, Wang Q, Cheng J, Fu Y, Jiang D. A Novel Deltaflexivirus that Infects the Plant Fungal Pathogen, Sclerotinia sclerotiorum, Can Be Transmitted Among Host Vegetative Incompatible Strains. Viruses. 2018; 10(6):295. https://doi.org/10.3390/v10060295
Chicago/Turabian StyleHamid, Muhammad Rizwan, Jiatao Xie, Songsong Wu, Shahzeen Kanwal Maria, Dan Zheng, Abdoulaye Assane Hamidou, Qihua Wang, Jiasen Cheng, Yanping Fu, and Daohong Jiang. 2018. "A Novel Deltaflexivirus that Infects the Plant Fungal Pathogen, Sclerotinia sclerotiorum, Can Be Transmitted Among Host Vegetative Incompatible Strains" Viruses 10, no. 6: 295. https://doi.org/10.3390/v10060295