Intracellular Localization of Blattella germanica Densovirus (BgDV1) Capsid Proteins

 and
and
Abstract
1. Introduction
2. Materials and Methods
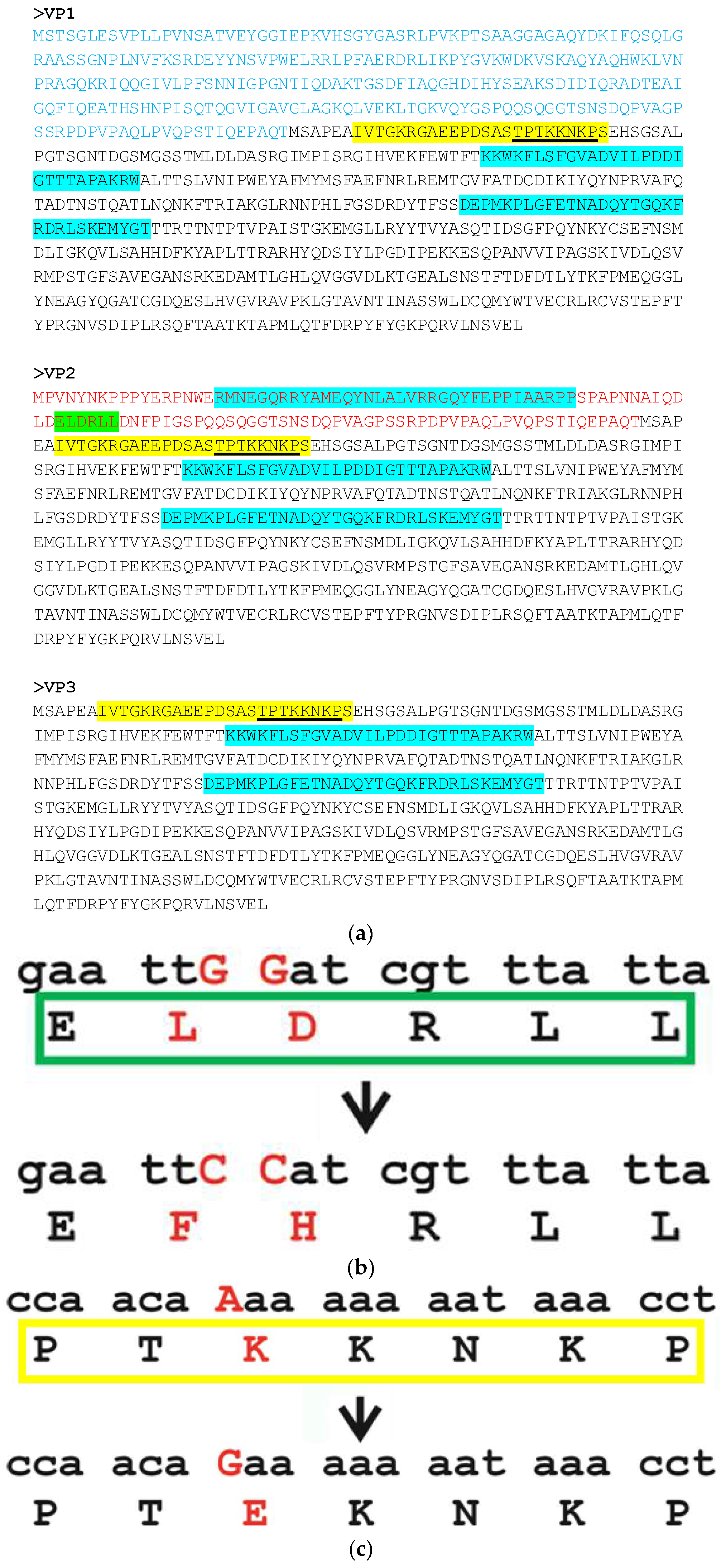
2.1. In Silico Analysis
2.2. Plasmid Construction
2.3. Site-Directed Mutagenesis
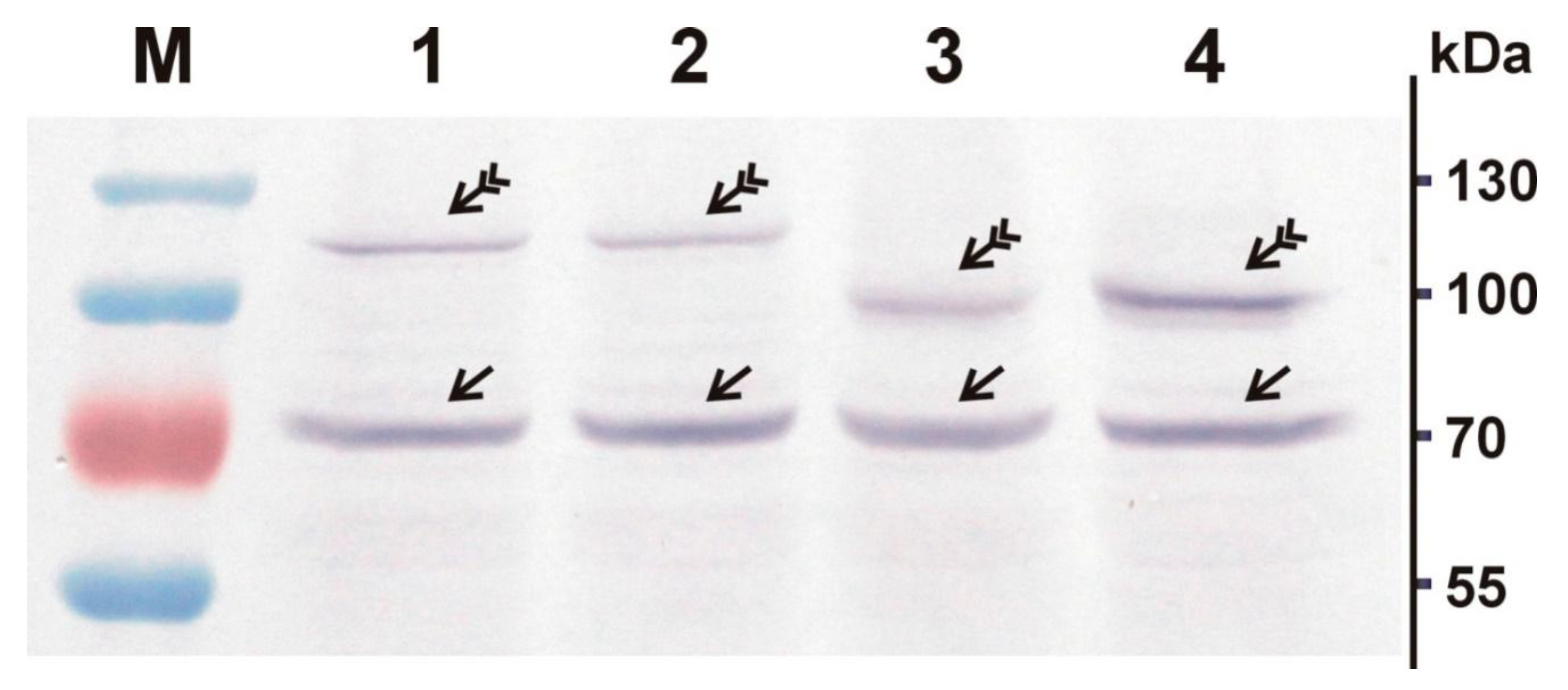
2.4. In Vitro Transcription and Translation
2.5. Protein Electrophoresis and Western Blotting
2.6. Cell Culture
2.7. Transfection
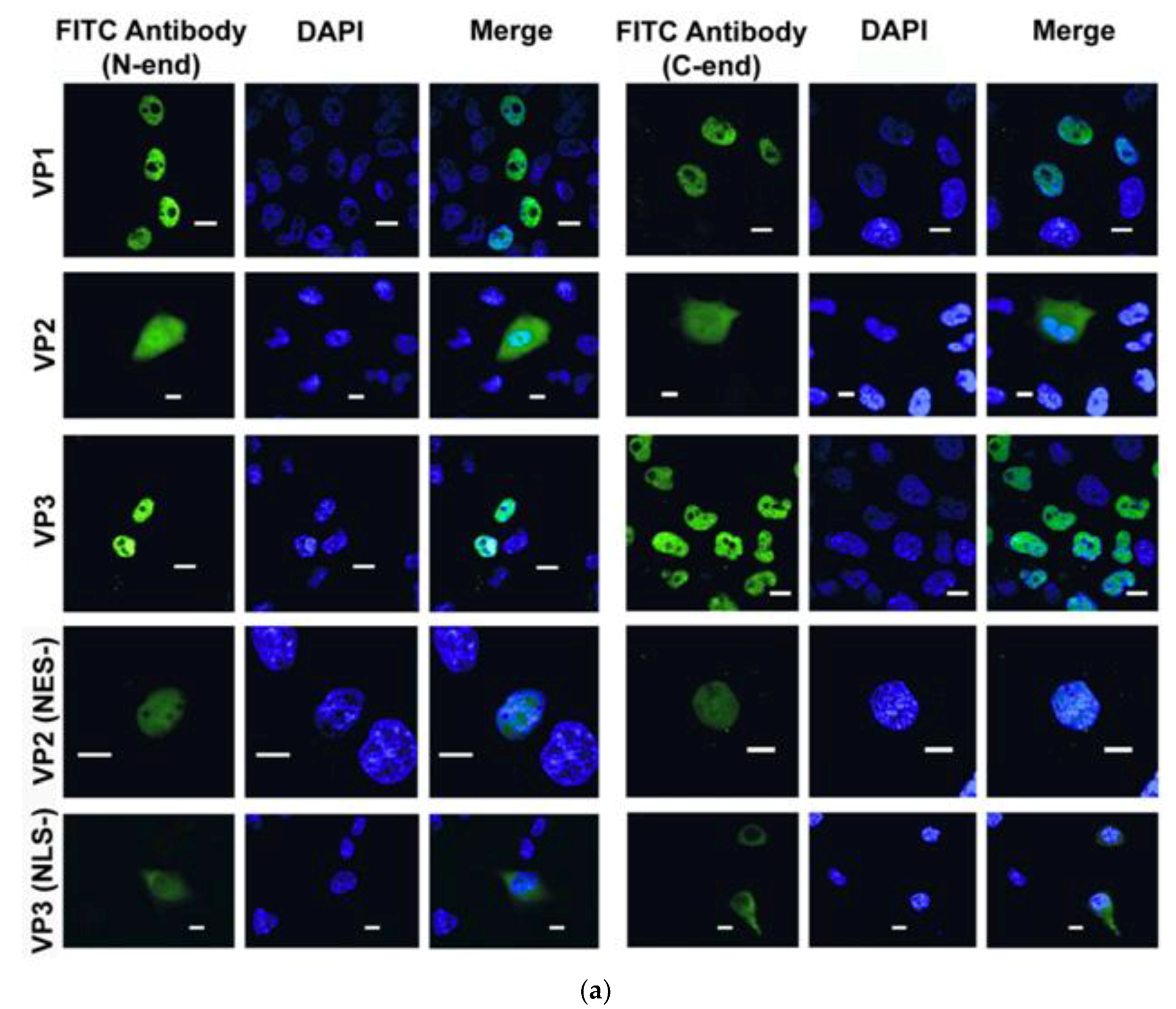
2.8. Immunohistochemistry
3. Results and Discussion
3.1. BgDV1 Nuclear Transport Signal Prediction
3.2. Nuclear Transport Signal Prediction for Other Densovirus Species
3.3. Intracellular Localization of BgDV1 Capsid Proteins
3.4. Functional Analysis of the Nuclear Localization and Nuclear Export Signals in BgDV1 Capsid Proteins
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cotmore, S.F.; Agbandje-McKenna, M.; Chiorini, J.A.; Mukha, D.V.; Pintel, D.J.; Qiu, J.; Soderlund-Venermo, M.; Tattersall, P.; Tijssen, P.; Gatherer, D.; et al. The family Parvoviridae. Arch. Virol. 2014, 159, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Bergoin, M.; Tijssen, P. Molecular Biology of Densovirinae. Contrib. Microbiol. 2000, 4, 12–32. [Google Scholar] [PubMed]
- Tijssen, P.; Penzes, J.J.; Yu, Q.; Pham, H.T.; Bergoin, M. Diversity of small, single-stranded DNA viruses of invertebrates and their chaotic evolutionary past. J. Invertebr. Pathol. 2016, 140, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Agbandje, M.; Chapman, M.S. Correlating structure with function in the viral capsid. In Parvoviruses; Kerr, J.R., Cotmore, S.F., Bloom, M.E., Linden, R.M., Parrish, C.R., Eds.; Hodder Arnold: London, UK, 2006; pp. 125–139. ISBN 0-340-81198-6. [Google Scholar]
- Tijssen, P.; Li, Y.; El-Far, M.; Szelei, J.; Letarte, M.; Zádori, Z. Organization and expression strategy of the ambisense genome of densonucleosis virus of Galleria mellonella. J. Virol. 2003, 77, 10357–10365. [Google Scholar] [CrossRef] [PubMed]
- Bruemmer, A.; Scholari, F.; Lopez-Ferber, M.; Conway, J.F.; Hewat, E.A. Structure of an insect parvovirus (Junonia coenia Densovirus) determined by cryo-electron microscopy. J. Mol. Biol. 2005, 347, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Fédière, G.; El-Far, M.; Li, Y.; Bergoin, M.; Tijssen, P. Expression strategy of densonucleosis virus from Mythimna loreyi. Virology 2004, 320, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Meng, G.; Zhang, X.; Plevka, P.; Yu, Q.; Tijssen, P.; Rossmann, M.G. The structure and host entry of an invertebrate parvovirus. J. Virol. 2013, 87, 12523–12530. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Li, Y.; Jousset, F.X.; Zadori, Z.; Szelei, J.; Yu, Q.; Pham, H.T.; Lépine, F.; Bergoin, M.; Tijssen, P. The Acheta domesticus densovirus, isolated from the European house cricket, has evolved an expression strategy unique among parvoviruses. J. Virol. 2011, 84, 10069–10078. [Google Scholar] [CrossRef] [PubMed]
- Baquerizo-Audiot, E.; Abd-Alla, A.; Jousset, F.; Cousserans, F.; Tijssen, P.; Bergoin, M. Structure and expression strategy of the genome of Culex pipiens densovirus, a mosquito densovirus with an ambisense organization. J. Virol. 2009, 83, 6863–6873. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zheng, J.; Iizuka, T.; Bando, H. A densovirus newly isolated from the smoky-brown cockroach Periplaneta fuliginosa. Arch. Virol. 1994, 138, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, J.; Hu, Y.; Zheng, J.; Bando, H. Genome organization and mRNA structure of Periplaneta fuliginosa densovirus imply alternative splicing involvement in viral gene expression. Arch. Virol. 1999, 144, 2111–2124. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; He, J.; Huang, X.; Dai, A.; Cheng, L.; Shao, D.; Zhang, J. The truncated virus-like particles of C6/36 cell densovirus: Implications for the assembly mechanism of brevidensovirus. Virus Res. 2008, 132, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Shike, H.; Dhar, A.K.; Burns, J.C.; Shimizu, C.; Jousset, F.X.; Klimpel, K.R.; Bergoin, M. Infectious hypodermal and hematopoetic necrosis virus of shrimp is related to mosquito Brevidensoviruses. Virology 2000, 277, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zadori, Z.; Bando, H.; Dubuc, R.; Fediere, G.; Szelei, J.; Tijssen, P. Genome organization of the densovirus from Bombyx mori (BmDNV-1) and enzyme activity of its capsid. J. Gen. Virol. 2001, 82, 2821–2825. [Google Scholar] [CrossRef] [PubMed]
- Nakagaki, M.; Kawase, S. Structural proteins of densonucleosis virus isolated from the silkworm, Bombyx mori, infected with the flacherie virus. J. Invertebr. Pathol. 1980, 36, 166–171. [Google Scholar] [CrossRef]
- Fediere, G.; Li, Y.; Zadori, Z.; Szelei, J.; Tijssen, P. Genome organization of Casphalia extranea densovirus, a new Iteravirus. Virology 2002, 292, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Sukhumsirichart, W.; Attasart, P.; Boonsaeng, V.; Panyim, S. Complete nucleotide sequence and genomic organization of hepatopancreatic parvovirus (HPV) of Penaeus monodon. Virology 2006, 346, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Zadori, Z.; Szelei, J.; Lacoste, M.-C.; Li, Y.; Gariepy, S.; Raymond, P.; Allaire, M.; Nabi, I.R.; Tijssen, P. A viral phospholipase A2 is required for parvoviral infectivity. Dev. Cell 2001, 1, 291–302. [Google Scholar] [CrossRef]
- Kalderon, D.; Roberts, B.L.; Richardson, W.D.; Smith, A.E. A short amino acid sequence able to specify nuclear location. Cell 1984, 39, 499–509. [Google Scholar] [CrossRef]
- Dang, C.V.; Lee, W.M. Identification of the human c-myc protein nuclear translocation signal. Mol. Cell. Biol. 1988, 8, 4048–4054. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Hasebe, M.; Matsumura, N.; Takashima, H.; Miyamoto-Sato, E.; Tomita, M.; Yanagawa, H. Six classes of nuclear localization signals specific to different binding grooves of importin α. J. Biol. Chem. 2009, 284, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Jans, D.A.; Xiao, C.-Y.; Lam, M.H.C. Nuclear targeting signal recognition: A key control point in nuclear transport? BioEssays 2000, 22, 532–544. [Google Scholar] [CrossRef]
- Dingwall, C.; Robbins, J.; Dilworth, S.M.; Roberts, B.; Richardson, W.D. The nucleoplasmin nuclear location sequence is larger and more complex than that of SV-40 large T antigen. J. Cell Biol. 1988, 107, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Robbins, J.; Dilworth, S.M.; Laskey, R.A.; Dingwall, C. Two interdependent basic domains in nucleoplasmin nuclear targeting sequence: Identification of a class of bipartite nuclear targeting sequence. Cell 1991, 64, 615–623. [Google Scholar] [CrossRef]
- Freitas, N.; Cunha, C. Mechanisms and signals for the nuclear import of proteins. Curr. Genom. 2009, 10, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Owens, L. Bioinformatical analysis of nuclear localization sequences in penaeid densoviruses. Mar. Genom. 2013, 12, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Boisvert, M.; Bouchard-Lévesque, V.; Fernandes, S.; Tijssen, P. Classic nuclear localization signals and a novel nuclear localization motif are required for nuclear transport of Porcine Parvovirus capsid proteins. J. Virol. 2014, 88, 11748–11759. [Google Scholar] [CrossRef] [PubMed]
- Chelsky, D.; Ralph, R.; Jonak, G. Sequence requirements for synthetic peptide mediated translocation to the nucleus. Mol. Cell. Biol. 1989, 9, 2487–2492. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Farmer, A.; Collett, G.; Crishin, N.V.; Chook, Yu.M. Sequence and structural analyses of nuclear export signals in the NESdb database. Mol. Biol. Cell 2012, 23, 3677–3693. [Google Scholar] [CrossRef] [PubMed]
- Vendeville, A.; Ravallec, M.; Jousset, F.X.; Devise, M.; Mutuel, D.; Lopez-Ferber, M.; Fournier, P.; Dupressoir, T.; Oqliastro, M. Densovirus infectious pathway requires clathrin-mediated endocytosis followed by trafficking to the nucleus. J. Virol. 2009, 83, 4678–4689. [Google Scholar] [CrossRef] [PubMed]
- Briassouli, A.; Kompatsiaris, I. Nuclear transport of the major capsid proteins is essential for adeno-associated virus capsid formation. J. Virol. 1999, 73, 7912–7915. [Google Scholar]
- Grieger, J.C.; Snowdy, S.; Samulski, R.J. Separate basic region motifs within the adeno-associated virus capsid proteins are essential for infectivity and assembly. J. Virol. 2006, 80, 5199–5210. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, E.; Ramírez, J.C.; Garcia, J.; Almendral, J.M. Complementary roles of multiple nuclear targeting signals in the capsid proteins of the parvovirus minute virus of mice during assembly and onset of infection. J. Virol. 2002, 76, 7049–7059. [Google Scholar] [CrossRef] [PubMed]
- Pillet, S.; Annan, Z.; Fichelson, S.; Morinet, F.R. Identification of a nonconventional motif necessary for the nuclear import of the human parvovirus B19 major capsid protein (VP2). Virology 2003, 306, 25–32. [Google Scholar] [CrossRef]
- Vihinen-Ranta, M.; Kakkola, L.; Kalela, A.; Vilja, P.; Vuento, M. Characterization of a nuclear localization signal of canine parvovirus capsid proteins. Eur. J. Biochem. 1997, 250, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Vihinen-Ranta, M.; Wang, D.; Weichert, W.; Parrish, C.R. The VP1 N-terminal sequence of canine parvovirus affects nuclear transport of capsids and efficient cell infection. J. Virol. 2002, 76, 1884–1891. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Chen, Sh.; Wang, M.; Cheng, A. The role of nuclear localization signal in parvovirus life cycle. Virol. J. 2017, 14, 80. [Google Scholar] [CrossRef] [PubMed]
- Afanasiev, B.N.; Gaylov, E.E.; Buchatsky, L.P.; Kozlov, Y.V. Nucleotide sequence and genomic organization of Aedes densonucleosis virus. Virology 1991, 185, 323–336. [Google Scholar] [CrossRef]
- Chen, S.; Cheng, L.; Zhang, Q.; Lin, W.; Lu, X.; Brannan, J.; Zhou, Z.H.; Zhang, J. Genetic, biochemical, and structural characterization of a new densovirus isolated from a chronically infected Aedes albopictus C6/36 cell line. Virology 2004, 318, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Zhang, J.; Yang, B.; Zhou, L.; Hu, Y. The nuclear localization signal of the NS1 protein is essential for Periplaneta fuliginosa densovirus infection. Virus Res. 2009, 145, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Valle, N.; Riolobos, L.; Almendral, J.M. Synthesis, post-translational modification and trafficking of the parvovirus structural polypeptides. In Parvoviruses; Kerr, J.R., Cotmore, S.F., Bloom, M.E., Linden, R.M., Parrish, C.R., Eds.; Hodder Arnold: London, UK, 2006; pp. 291–304. ISBN 0-340-81198-6. [Google Scholar]
- Wolfsberg, R.; Kempf, C.; Ros., C. Late maturation steps preceding selective nuclear export and egress of progeny parvovirus. J. Virol. 2016, 90, 5462–5474. [Google Scholar] [CrossRef] [PubMed]
- Maroto, B.; Valle, N.; Saffrich, R.; Almendral, J.M. Nuclear export of the nonenveloped parvovirus virion is directed by an unordered protein signal exposed on the capsid surface. J. Virol. 2004, 78, 10685–10694. [Google Scholar] [CrossRef] [PubMed]
- Hutten, S.; Kehlenbach, R.H. CRM1-mediated nuclear export to the pore and beyond. Trends Cell Biol. 2007, 17, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Eichwald, V.; Daeffler, L.; Klein, M.; Rommelaere, J.; Salome, N. The NS2 proteins of parvovirus minute virus of mice are required for efficient nuclear egress of progeny virions in mouse cells. J. Virol. 2002, 76, 10307–10319. [Google Scholar] [CrossRef] [PubMed]
- Engelsma, D.; Valle, N.; Fish, A.; Salomé, N.; Almendral, J.M.; Fornerod, M. A supraphysiological nuclear export signal is required for parvovirus nuclear export. Mol. Biol. Cell 2008, 19, 2544–2552. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.L.; Pintel, D.J. Interaction between parvovirus NS2 protein and nuclear export factor Crm1 is important for viral egress from the nucleus of murine cells. J. Virol. 2002, 76, 3257–3266. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, T.; Nakajima, T.; Oishi, T.; Imamoto, N.; Yoneda, Y.; Fukamizu, A.; Yagami, K. CRM1 mediates nuclear export of nonstructural protein 2 from parvovirus minute virus of mice. Biochem. Biophys. Res. Commun. 1999, 264, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Mukha, D.; Chumachenko, A.; Dykstra, M.; Kurtti, T.; Schal, C. Characterization of a new densovirus infecting the German cockroach, Blattella germanica. J. Gen. Virol. 2006, 87, 1567–1575. [Google Scholar] [CrossRef] [PubMed]
- Kapelinskaya, T.; Martynova, E.; Schal, C.; Mukha, D. Expression strategy of densonucleosis virus from the German cockroach, Blattella germanica. J. Virol. 2011, 85, 11855–11870. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, E.N.; Mukha, D.V. Mammalian cell culture as a model for studying the intracellular traffic of densovirus proteins. Russ. J. Genet. 2015, 51, 218–222. [Google Scholar] [CrossRef]
- Martynova, E.; Kapelinskaya, T.; Schal, C.; Mukha, D. Intracellular localization of regulatory proteins of the German cockroach, Blattella germanica, densovirus. Mol. Biol. (Mosc.) 2014, 48, 301–304. [Google Scholar] [CrossRef]
- Kosugi, S.; Hasebe, M.; Tomita, M.; Yanagawa, H. Systematic identification of yeast cell cycle-dependent nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc. Natl. Acad. Sci. USA 2009, 106, 10171–10176. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed]
- La Cour, T.; Kiemer, L.; Mølgaard, A.; Gupta, R.; Skriver, K.; Brunak, S. Analysis and prediction of leucine-rich nuclear export signals. Protein Eng. Des. Sel. 2004, 17, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.; Hunt, H.; Horton, R.; Pullen, J.; Pease, L. Site-directed mutagenesis by overlap extension using the polymerase chain reaction. Gene 1989, 77, 51–59. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Tijssen, P. Gene expression of five different Iteradensoviruses: Bombyx mori densovirus, Casphalia extranea densovirus, Papilio polyxenes densovirus, Sibine fusca densovirus, and Danaus plexippus densovirus. J. Virol. 2014, 88, 12152–12157. [Google Scholar] [CrossRef] [PubMed]
- Martynova, E.U.; Schal, C.; Mukha, D.V. Effects of recombination on densovirus phylogeny. Arch. Virol. 2016, 161, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, B.; Simpson, A.A.; Rossmann, M.G. The structure of human parvovirus B19. Proc. Natl. Acad. Sci. USA 2004, 101, 11628–11633. [Google Scholar] [CrossRef] [PubMed]
- Simpson, A.A.; Paul R Chipman, P.R.; Baker, T.S.; Tijssen, P.; Rossmann, M.G. The structure of an insect parvovirus (Galleria mellonella densovirus) at 3.7 Å resolution. Structure 1998, 6, 1355–1367. [Google Scholar] [CrossRef]
- Kaufmann, B.; El-Far, M.; Plevka, P.; Bowman, V.D.; Li, Y.; Tijssen, P.; Rossmann, M.G. Structure of Bombyx mori densovirus 1, a silkworm pathogen. J. Virol. 2011, 85, 4691–4697. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, B; Bowman, V.D.; Li, Y.; Szelei, J.; Waddell, P.J.; Tijssen, P.; Rossmann, M.G. The structure of Penaeus stylirostris densovirus, a shrimp pathogen. J. Virol. 2010, 84, 11289–11296. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Chen, S.; Brannan, J.M.; Jakana, J.; Zhang, Q.; Zhou, Z.H.; Zhang, J. Three-dimensional structure determination of capsid of Aedes albopictus C6/36 cell densovirus. Sci. China Ser. C Life Sci. 2004, 47, 224–228. [Google Scholar] [CrossRef]
- Guan, Y.; Zu, Q.; Huang, D.; Zhao, S.; Lo, L.-J.; Peng, J. An equation to estimate the difference between theoretically predicted and SDS PAGE-displayed molecular weights for an acidic peptide. Sci. Rep. 2015, 5, 13370. [Google Scholar] [CrossRef] [PubMed]
- Rath, A.; Glibowicka, M.; Nadeau, V.G.; Chen, G.; Deber, C.M. Detergent binding explains anomalous SDS-PAGE migration of membrane proteins. Proc. Natl. Acad. Sci. USA 2009, 106, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-S.; Jan, M.-S.; Chou, C.-K.; Chen, P.-H.; Ke, N.-J. Is green fluorescent protein toxic to the living cells? Biochem. Biophys. Res. Commun. 1999, 260, 712–717. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Virus | NLS * | Score | VP Protein (VP-Coding ORF) Containing NLS Motif ** | NLS Amino Acid Positions *** | NES | VP Protein (VP Coding ORF) Cjntaining NES Motif ** | NES Amino Acid Positions *** |
|---|---|---|---|---|---|---|---|---|
| Ambidensovirus | Lepidopteran densoviruses | Consensus motif:ExTKRKA(D/G)(S/T)(S/P)(A/V)xE(T/G)P(A/S)KKG(A/T)(T/H), “x” is any amino acid | VP1 | 326–346 | No | |||
| VP2 | 50–70 | |||||||
| VP3 | 3–23 | |||||||
| GmDV | EATKRKADSPAVETPAKKGTT | 6.0 | ||||||
| MlDV | EPTKRKAGSSAAETPAKKGAT | 6.5 | ||||||
| JcDV | EGTKRKADTPVEEGPSKKGAH | 6.5 | ||||||
| HaDV1 | EATKRKADTPAEEGPSKKGAH | 7.0 | ||||||
| PiDV | EGTKRKADSPVEEGPSKKGAH | 6.5 | ||||||
| DsDV | ESTKRKADTPAEETPSKKGAH | 7.5 | ||||||
| CpDV | LNKTGYGSNVSFTEFTAGKPMIPSKIAKP | 7.3 | VP ORF1 | 399–427 | Yes | VP ORF1 | 97–107 | |
| PfDV | GEPPNKKAKTG | 8.0 | VP ORF6 | 126–136 | Yes | VP ORF5 | 67–79 | |
| AdDV | DKGKGKRGGGGRPPKSSGGKRSR (low score) | 4.3 | VP1 | 365–387 | Yes | VP1 | 8–15 | |
| VP2 | 146–168 | |||||||
| VP3 | 16–138 | |||||||
| BgDV1 | TPTKKNKP(probably part of IVTGKRGAEEPDSASTPTKKNKPS) | 8.6 # | VP1 | 291–297(275–298) | Yes | |||
| VP2 | 141–147(125–148) | VP2 | 64–69 | |||||
| VP3 | 23–29(7–30) | |||||||
| BgDV2 | TYRKKWRFL | 6.5 | VP ORF1 | 182–190 | Yes | VP ORF1 | 59–66 | |
| VP ORF2 | 286–299 | |||||||
| PcDV | PRVSKKPRIS(probably part of WYRKYQFVNNLNDKGQPRVSKKPRIS) | 9.5(6.3) | smaller VP coding ORF | 74–83(58–83) | No | |||
| MpDV | NERKRIKLG | 7.0 | smaller VP coding ORF | 16–24 | Yes | smaller VP coding ORF | 89–100 | |
| SKRPADSSGSEPAPKRAGGT | 9.0 | larger VP coding ORF | 105–124 | |||||
| Unassigned | AdMADV | NRLKRLRLAAE | 8.0 | VP (ORF VP1) | 127–137 | No | ||
| Iteradensovirus | BmDV | GMGKRKSTEKDWAKIKRINR | 6.5 | N-terminal part of VP ORF (most probably VP1 and VP2 proteins) | 94–113 | Only CeDV | ||
| CeDV | GMGKRKSTEKDWAKIKRINT | 6.0 | 94–113 | VP ORF (most probably all VPs) | 283–295 | |||
| DpDV | GMGKRKNTEKDWATIKRINR | 5.5 | 92–111 | |||||
| PpDV | GMGKRKSNEKDWATIKRINR | 6.0 | 94–113 | |||||
| DpplDV | GMGKRKSTEKDWAKIKRINR | 6.0 | 94–113 | |||||
| SfDV | GMGKRKPNEDNIEKDWAKIKRINR | 6.0 | 94–117 | |||||
| CeDV | PVTRSKK | -- ## | C-terminus of VP ORF (most probably common C-terminus of all capsid proteins) | 668–674 | ||||
| PpDV | PVTRSKK | -- | 663–669 | |||||
| SfDV | PVTRSKK | -- | 672–678 | |||||
| DpplDV | PVTRSKK | -- | 667–673 | |||||
| HaDV2 | KNFTEEKIVGRPLYGMPTKDWGRIKKI (low score) | 2.9 | N-terminal part of VP ORF (VP1 and VP2) | 79–105 | No | |||
| Brevidensovirus | Consensus motif:RGTKRKRx(A/T)x | N-terminus of VP ORF (most probably VP1 and VP2) | ||||||
| AalDV3, HeDV, AalDV2 | RGTKRKREAD | 11.0 | 12–21 | Yes | All VP proteins | 108–117 | ||
| AalDV1 | RGTKRKRETG | 10.0 | 14–23 | Yes | 111–120 | |||
| AaeDV1 | RGTKRKRDAG | 10.0 | 14–23 | No | ||||
| AaeDV2 | RGTKRKRGAG | 10.0 | 14–23 | Yes | 111–120 | |||
| AgDV, CppDV | RGTKRKRGAE | 11.0 | 14–23 | Yes | 109–118 | |||
| Penstyldensovirus | Described in Owens et al., 2013 [27]. | No | ||||||
| Hepandensovirus | Described in Owens et al., 2013 [27]. | Yes | VP ORF | 174–181 | ||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozlov, E.N.; Martynova, E.U.; Popenko, V.I.; Schal, C.; Mukha, D.V. Intracellular Localization of Blattella germanica Densovirus (BgDV1) Capsid Proteins. Viruses 2018, 10, 370. https://doi.org/10.3390/v10070370
Kozlov EN, Martynova EU, Popenko VI, Schal C, Mukha DV. Intracellular Localization of Blattella germanica Densovirus (BgDV1) Capsid Proteins. Viruses. 2018; 10(7):370. https://doi.org/10.3390/v10070370
Chicago/Turabian StyleKozlov, Evgeny N., Elena U. Martynova, Vladimir I. Popenko, Coby Schal, and Dmitry V. Mukha. 2018. "Intracellular Localization of Blattella germanica Densovirus (BgDV1) Capsid Proteins" Viruses 10, no. 7: 370. https://doi.org/10.3390/v10070370
APA StyleKozlov, E. N., Martynova, E. U., Popenko, V. I., Schal, C., & Mukha, D. V. (2018). Intracellular Localization of Blattella germanica Densovirus (BgDV1) Capsid Proteins. Viruses, 10(7), 370. https://doi.org/10.3390/v10070370