Characterisation of the Virome of Tonsils from Conventional Pigs and from Specific Pathogen-Free Pigs



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Nucleic Acid Extraction
2.3. Nucleic Acid Amplification and Sequencing
2.4. High-Throughput Sequencing (HTS) Data Annotation
2.5. Viral Genetic Analyses
3. Results
3.1. Sequencing Data and Annotation
3.2. Adenovirus
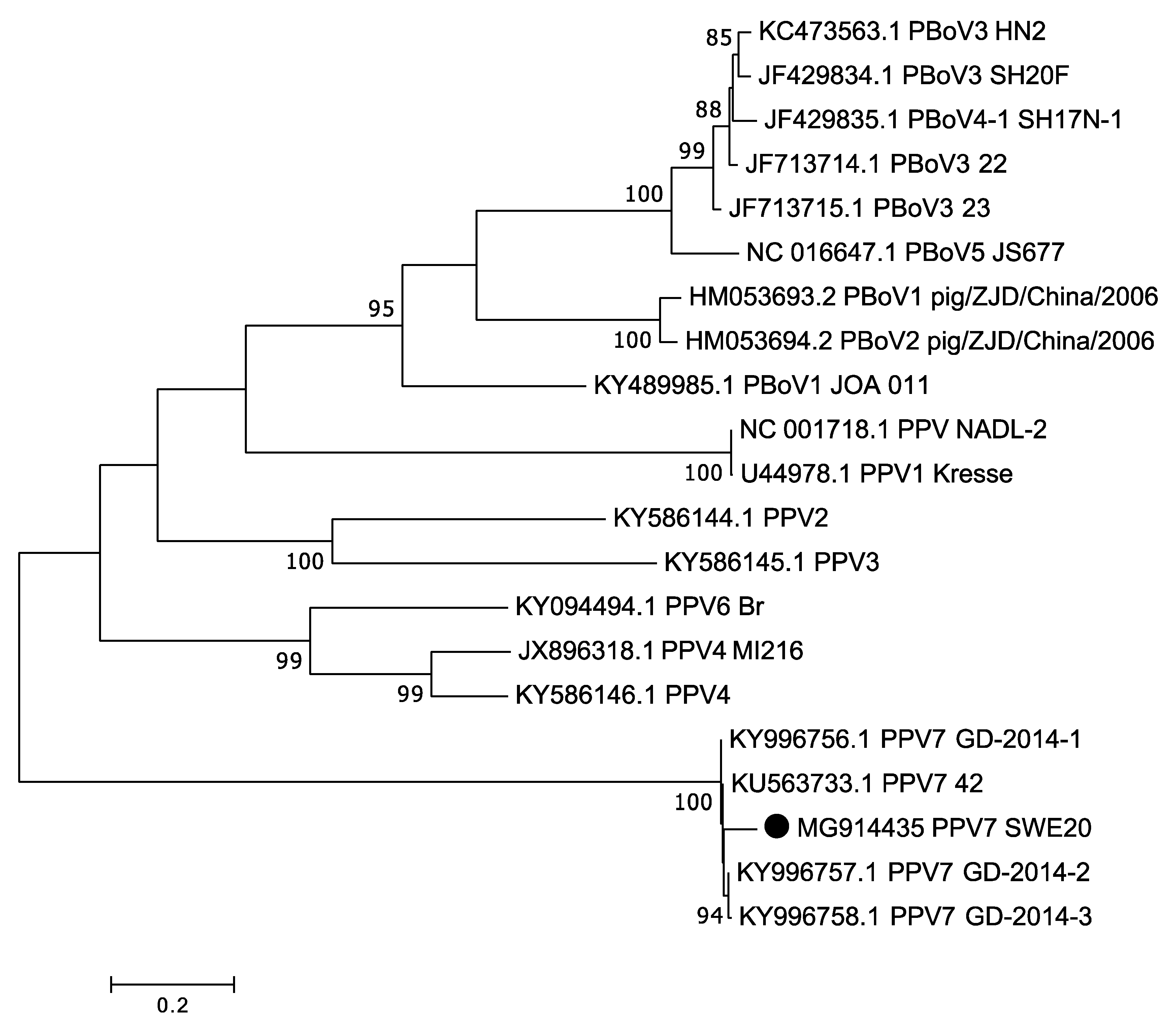
3.3. Porcine Parvovirus 7
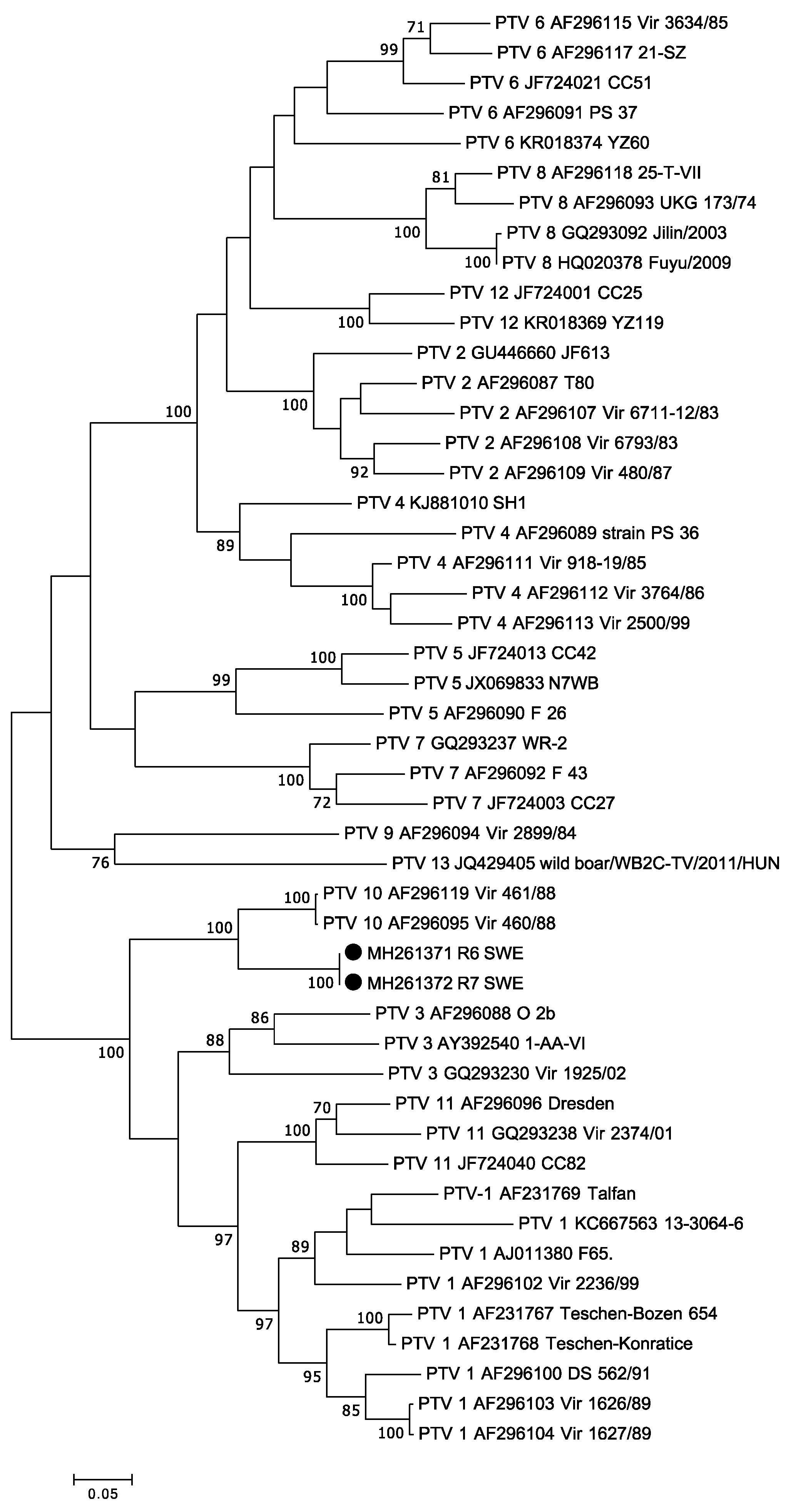
3.4. Teschovirus
3.5. Other Viruses
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brockmeier, S.L.; Halbur, P.G.; Thacker, E.-L. Porcine respiratory disease complex. In Polymicrobial Diseases; Brogden, K.A., Guthmiller, J.M., Eds.; ASM Press: Washington, DC, USA, 2012; pp. 231–258. [Google Scholar]
- Opriessnig, T.; Gimenez-Lirola, L.G.; Halbur, P.G. Polymicrobial respiratory disease in pigs. Anim. Health Res. Rev. 2011, 12, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Neumann, E.J.; Kliebenstein, J.B.; Johnson, C.D.; Mabry, J.W.; Bush, E.J.; Seitzinger, A.H.; Green, A.L.; Zimmerman, J.J. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J. Am. Vet. Med. Assoc. 2005, 227, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuis, N.; Duinhof, T.F.; van Nes, A. Economic analysis of outbreaks of porcine reproductive and respiratory syndrome virus in nine sow herds. Vet. Rec. 2012, 170, 225. [Google Scholar] [CrossRef] [PubMed]
- Blomström, A.L. Viral metagenomics as an emerging and powerful tool in veterinary medicine. Vet. Quart. 2011, 31, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Delwart, E.L. Viral metagenomics. Rev. Med. Virol. 2007, 17, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Roossinck, M.J.; Martin, D.P.; Roumagnac, P. Plant virus metagenomics: Advances in virus discovery. Phytopathology 2015, 105, 716–727. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Giannitti, F.; Rossow, S.; Marthaler, D.; Knutson, T.P.; Li, L.; Deng, X.; Resende, T.; Vannucci, F.; Delwart, E. Detection of a novel circovirus PCV3 in pigs with cardiac and multi-systemic inflammation. Virol. J. 2016, 13, 184. [Google Scholar] [CrossRef] [PubMed]
- Blomström, A.L.; Belak, S.; Fossum, C.; McKillen, J.; Allan, G.; Wallgren, P.; Berg, M. Detection of a novel porcine boca-like virus in the background of porcine circovirus type 2 induced postweaning multisystemic wasting syndrome. Virus Res. 2009, 146, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Hause, B.M.; Collin, E.A.; Peddireddi, L.; Yuan, F.; Chen, Z.; Hesse, R.A.; Gauger, P.C.; Clement, T.; Fang, Y.; Anderson, G. Discovery of a novel putative atypical porcine pestivirus in pigs in the USA. J. Gen. Virol. 2015, 96, 2994–2998. [Google Scholar] [CrossRef] [PubMed]
- Delwart, E. Animal virus discovery: Improving animal health, understanding zoonoses, and opportunities for vaccine development. Curr. Opin. Virol. 2012, 2, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Ahlberg, V.; Hjertner, B.; Wallgren, P.; Hellman, S.; Lovgren Bengtsson, K.; Fossum, C. Innate immune responses induced by the saponin adjuvant Matrix-M in specific pathogen free pigs. Vet. Res. 2017, 48, 30. [Google Scholar] [CrossRef] [PubMed]
- Wallgren, P.; Segall, T.; Pedersen Morner, A.; Gunnarsson, A. Experimental infections with Actinobacillus pleuropneumoniae in pigs-I. Comparison of five different parenteral antibiotic treatments. Zentralbl. Veterinarmed. B 1999, 46, 249–260. [Google Scholar] [PubMed]
- Blomstrom, A.L.; Fossum, C.; Wallgren, P.; Berg, M. Viral metagenomic analysis displays the co-infection situation in healthy and PMWS affected pigs. PLoS ONE 2016, 11, e0166863. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Blomström, A.L.; Belak, S.; Fossum, C.; Fuxler, L.; Wallgren, P.; Berg, M. Studies of porcine circovirus type 2, porcine boca-like virus and torque teno virus indicate the presence of multiple viral infections in postweaning multisystemic wasting syndrome pigs. Virus Res. 2010, 152, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Holtkamp, D.J.; Kliebenstein, J.B.; Neumann, E.J.; Zimmerman, J.J.; Rotto, H.F.; Yoder, T.K.; Wang, C.; Yeske, P.E.; Mowrer, C.L.; Haley, C.A. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J. Swine Health Prod. 2013, 21, 72–84. [Google Scholar]
- Shan, T.; Li, L.; Simmonds, P.; Wang, C.; Moeser, A.; Delwart, E. The fecal virome of pigs on a high-density farm. J. Virol. 2011, 85, 11697–11708. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, O.E.; Larsson, J.; Hayer, J.; Berg, M.; Jacobson, M. The intestinal eukaryotic virome in healthy and diarrhoeic neonatal piglets. PLoS ONE 2016, 11, e0151481. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Renukaradhya, G.J.; Alekseev, K.P.; Fang, Y.; Tang, Y.; Saif, L.J. Porcine reproductive and respiratory syndrome virus modifies innate immunity and alters disease outcome in pigs subsequently infected with porcine respiratory coronavirus: Implications for respiratory viral co-infections. J. Gen. Virol. 2009, 90, 2713–2723. [Google Scholar] [CrossRef] [PubMed]
- Van Reeth, K.; Nauwynck, H.; Pensaert, M. Dual infections of feeder pigs with porcine reproductive and respiratory syndrome virus followed by porcine respiratory coronavirus or swine influenza virus: A clinical and virological study. Vet. Microbiol. 1996, 48, 325–335. [Google Scholar] [CrossRef]
- Harms, P.A.; Sorden, S.D.; Halbur, P.G.; Bolin, S.R.; Lager, K.M.; Morozov, I.; Paul, P.S. Experimental reproduction of severe disease in CD/CD pigs concurrently infected with type 2 porcine circovirus and porcine reproductive and respiratory syndrome virus. Vet. Pathol. 2001, 38, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Rovira, A.; Balasch, M.; Segales, J.; Garcia, L.; Plana-Duran, J.; Rosell, C.; Ellerbrok, H.; Mankertz, A.; Domingo, M. Experimental inoculation of conventional pigs with porcine reproductive and respiratory syndrome virus and porcine circovirus 2. J. Virol. 2002, 76, 3232–3239. [Google Scholar] [CrossRef] [PubMed]
- Dobrescu, I.; Levast, B.; Lai, K.; Delgado-Ortega, M.; Walker, S.; Banman, S.; Townsend, H.; Simon, G.; Zhou, Y.; Gerdts, V.; et al. In vitro and ex vivo analyses of co-infections with swine influenza and porcine reproductive and respiratory syndrome viruses. Vet. Microbiol. 2014, 169, 18–32. [Google Scholar] [CrossRef] [PubMed]
- De Baere, M.I.; Van Gorp, H.; Delputte, P.L.; Nauwynck, H.J. Interaction of the European genotype porcine reproductive and respiratory syndrome virus (PRRSV) with sialoadhesin (CD169/Siglec-1) inhibits alveolar macrophage phagocytosis. Vet. Res. 2012, 43, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiou, M.T.; Jeng, C.R.; Chueh, L.L.; Cheng, C.H.; Pang, V.F. Effects of porcine reproductive and respiratory syndrome virus (isolate tw91) on porcine alveolar macrophages in vitro. Vet. Microbiol. 2000, 71, 9–25. [Google Scholar] [CrossRef]
- Benfield, D.A.; Hesse, R.A. Porcine adenoviruses. In Diseases of Swine; Zimmerman, J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp. 392–395. [Google Scholar]
- International Committee on Taxonomy of Viruses; King, A.M.Q. Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; Academic Press: London, UK; Waltham, MA, USA, 2012; p. 1327. [Google Scholar]
- Zhou, F.; Sun, H.; Wang, Y. Porcine bocavirus: Achievements in the past five years. Viruses 2014, 6, 4946–4960. [Google Scholar] [CrossRef] [PubMed]
- Palinski, R.M.; Mitra, N.; Hause, B.M. Discovery of a novel Parvovirinae virus, porcine parvovirus 7, by metagenomic sequencing of porcine rectal swabs. Virus Genes 2016, 52, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.; Zhou, H.; Tong, L.; Chen, Y.; Sun, Y.; Wang, H.; Zhang, G. First identification of porcine parvovirus 7 in China. Arch. Virol. 2018, 163, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Alexandersen, S.; Knowles, N.J.; Dekker, A.; Belsham, G.J.; Zhang, Z.; Koenen, F. Picornaviruses. In Disease of Swine, 10th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2012; pp. 587–620. [Google Scholar]



{kind=link}
{kind=link}
| S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | R1 | R2 | R3 | R4 | R5 | R6 | R7 | R8 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Adenoviridae | Porcine mastadenovirus A | 24 | 8 | 66 | 50 | 112 | 4 | 945 | 245 | 2 | 124 | 13 | 14 | 98 | 69 | 38 | |
| Porcine mastadenovirus B | 376 | 2147 | 6619 | 1361 | 14666 | 7 | 3615 | 1191 | 230 | 9584 | |||||||
| Other mastadenovirus | 609 | 24,010 | 3167 | 3 | 6561 | 1073 | 19478 | 5 | 7901 | 2255 | 206 | 8138 | |||||
| Herpesviridae | Suid herpesvirus 2 | 4 | 16 | 4 | 50 | 752 | |||||||||||
| Porcine lymphotrophic herpesvirus 1 | 30 | ||||||||||||||||
| Parvoviridae | Porcine bocavirus 1 | 4 | 2 | 2 | 2 | 88 | 2 | 27 | 4 | 31 | 1 | 4 | |||||
| Porcine bocavirus 3 | 1 | 1 | 3 | ||||||||||||||
| Porcine parvovirus 7 | 1 | 2 | 14131 | 14 | |||||||||||||
| Adeno-associated virus | 2 | 4 | 2 | 16 | |||||||||||||
| Uncharacterised parvovirus | 20 | 3 | 3 | 1 | 1 | ||||||||||||
| Circoviridae | Porcine circovirus 2 | 1 | 1 | ||||||||||||||
| Unclassified circovirus | 2 | ||||||||||||||||
| Anelloviridae | Torque teno sus virus 1 | 4 | 1 | 106 | 8 | 16 | 5 | ||||||||||
| Torque teno sus virus 2 | 1 | ||||||||||||||||
| Picornaviridae | Pasivirus A | 4 | 30 | 29 | 31 | 8 | 30 | 46 | 34 | 4 | 1 | 16 | 125 | 5 | 8 | ||
| Teschovirus | 1 | 8 | 6 | 2 | 67 | 5939 | 3385 | 85 | |||||||||
| Porcine sapelovirus A | 31 | 56 | |||||||||||||||
| Unassigned * | Posavirus 1 | 1 | 2 | 17 | 2 | 3 | |||||||||||
| Caliciviridae | Sapovirus | 1 | |||||||||||||||
| Astroviridae | Astrovirus | 2 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||
| Retroviridae | Porcine endogenous retrovirus | 80 | 27 | 47 | 87 | 120 | 50 | 80 | 102 | 128 | 93 | 59 | 64 | 88 | 173 | 133 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blomström, A.-L.; Ye, X.; Fossum, C.; Wallgren, P.; Berg, M. Characterisation of the Virome of Tonsils from Conventional Pigs and from Specific Pathogen-Free Pigs. Viruses 2018, 10, 382. https://doi.org/10.3390/v10070382
Blomström A-L, Ye X, Fossum C, Wallgren P, Berg M. Characterisation of the Virome of Tonsils from Conventional Pigs and from Specific Pathogen-Free Pigs. Viruses. 2018; 10(7):382. https://doi.org/10.3390/v10070382
Chicago/Turabian StyleBlomström, Anne-Lie, Xingyu Ye, Caroline Fossum, Per Wallgren, and Mikael Berg. 2018. "Characterisation of the Virome of Tonsils from Conventional Pigs and from Specific Pathogen-Free Pigs" Viruses 10, no. 7: 382. https://doi.org/10.3390/v10070382
APA StyleBlomström, A. -L., Ye, X., Fossum, C., Wallgren, P., & Berg, M. (2018). Characterisation of the Virome of Tonsils from Conventional Pigs and from Specific Pathogen-Free Pigs. Viruses, 10(7), 382. https://doi.org/10.3390/v10070382