Antiviral Effect of Resveratrol in Piglets Infected with Virulent Pseudorabies Virus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Compounds
2.2. Virus and Piglets
2.3. Ethics Statement
2.4. Experimental Design
2.5. Analysis of Viral Load by Real-Time PCR
2.6. Histopathological Analysis
2.7. Serum Cytokines Assay
2.8. Statistical Analysis
3. Results
3.1. Resveratrol Reduced Mortality and Increased Body Weight Gained by Piglets Infected with Virulent PRV
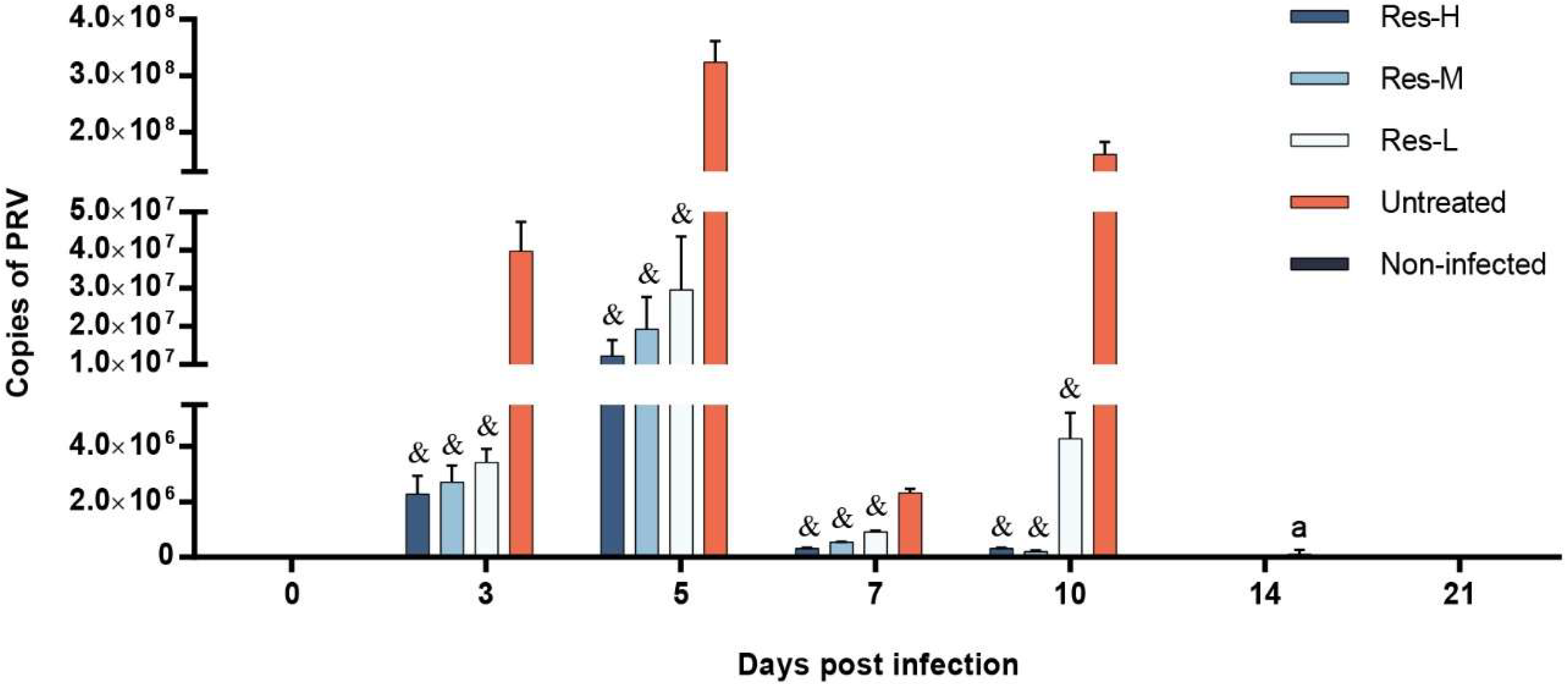
3.2. The Viral Load of Nasal Swab and Brain Were Depressed by Res
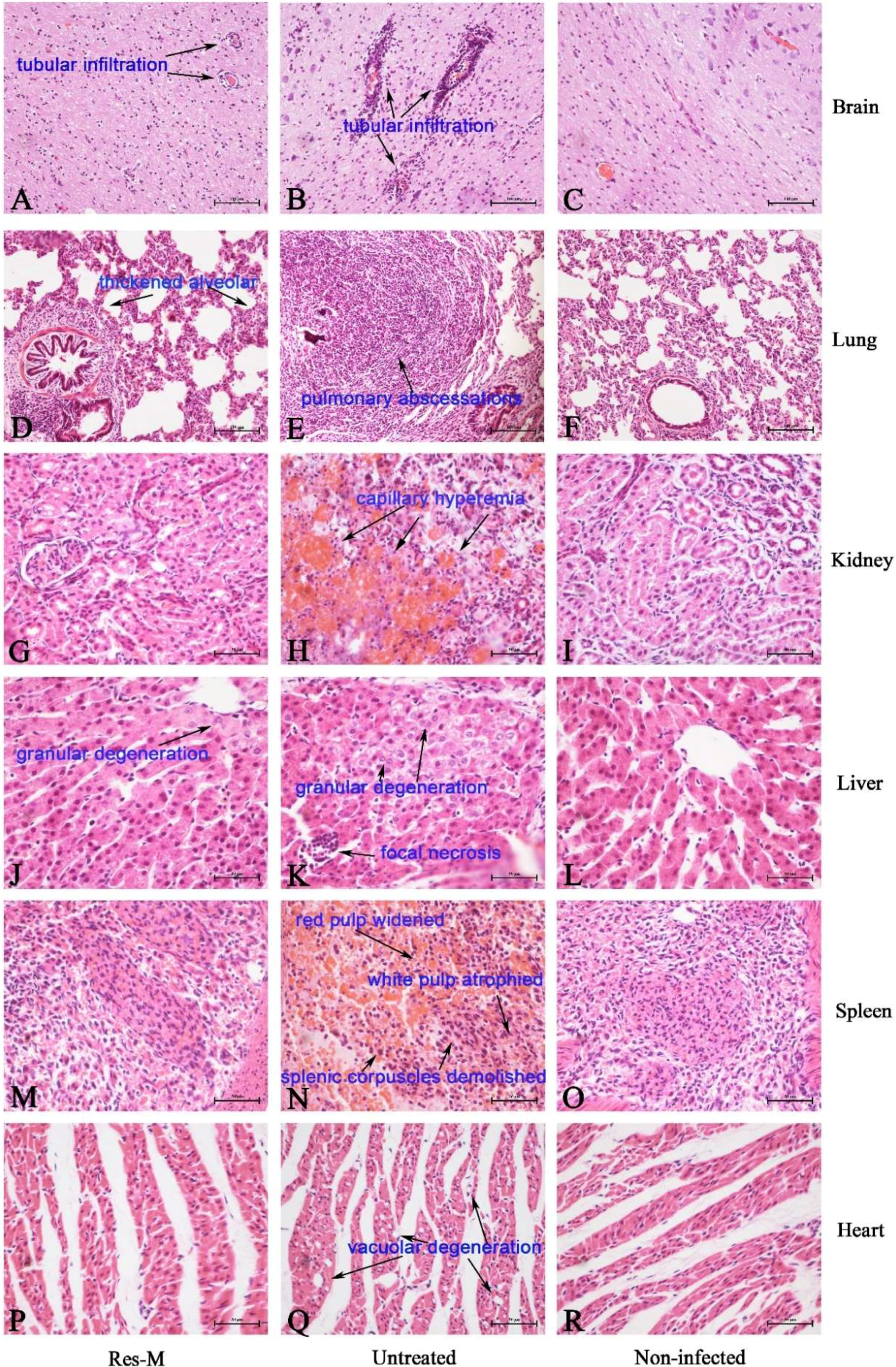
3.3. Res Reduced the Pathological Lesions of PRV-Infected Piglets
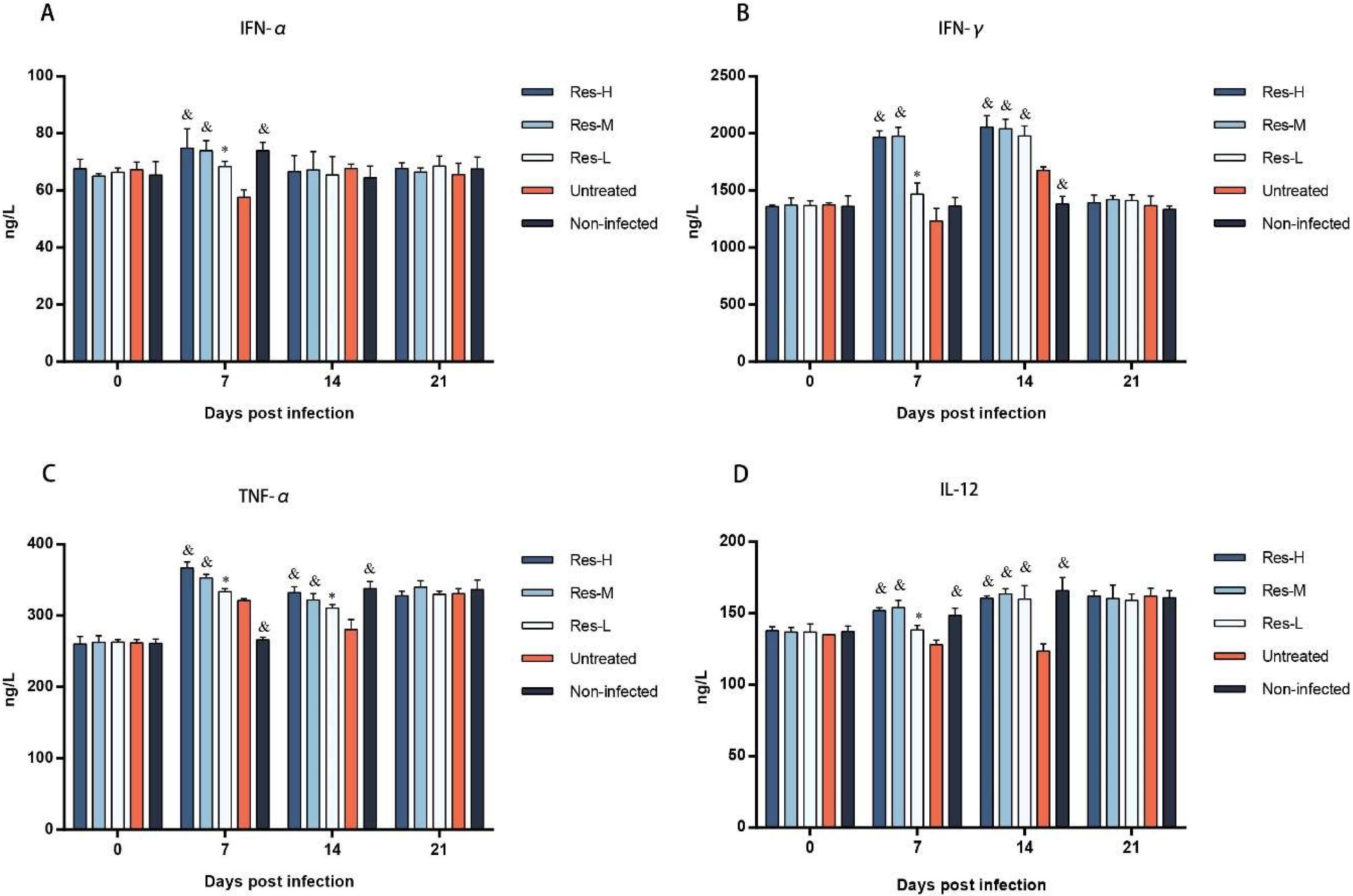
3.4. The Concentrations of Cytokines Were Affected by Res
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ma, W.; Lager, K.M.; Richt, J.A.; Stoffregen, W.C.; Zhou, F.; Yoon, K.J. Development of real-time polymerase chain reaction assays for rapid detection and differentiation of wild-type pseudorabies and gene-deleted vaccine viruses. J. Vet. Diagn. Investig. 2008, 20, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: Impact on neurovirology and veterinary medicine. Microbiol. Mol. Biol. Rev. 2005, 69, 462–500. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, G.; Rziha, H.-J. Aujeszky’s Disease (Pseudorabies) in Pigs; Springer: New York, NY, USA, 1989. [Google Scholar]
- Zuckermann, F.A. Aujeszky’s disease virus: Opportunities and challenges. Vet. Res. 2000, 31, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhou, Z.; Hu, D.; Zhang, Q.; Han, T.; Li, X.; Gu, X.; Yuan, L.; Zhang, S.; Wang, B.; et al. Pathogenic pseudorabies virus, China, 2012. Emerg. Infect. Dis. 2014, 20, 102–104. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Luo, Y.; Wang, C.H.; Yuan, J.; Li, N.; Song, K.; Qiu, H.J. Control of swine pseudorabies in China: Opportunities and limitations. Vet. Microbiol. 2016, 183, 119–124. [Google Scholar] [CrossRef] [PubMed]
- An, T.Q.; Peng, J.M.; Tian, Z.J.; Zhao, H.Y.; Li, N.; Liu, Y.M.; Chen, J.Z.; Leng, C.L.; Sun, Y.; Chang, D.; et al. Pseudorabies virus variant in Bartha-K61-vaccinated pigs, China, 2012. Emerg. Infect. Dis. 2013, 19, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Bai, C.; Sun, J.; Chang, S.; Zhang, X. Emergence of virulent pseudorabies virus infection in Northern China. J. Vet. Sci. 2013, 14, 363–365. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.; Liu, F.; Zheng, H.; Liang, C.; Zhou, Y.-J.; Jang, Y.-F.; Shan, T.-L.; Gao, F.; Li, G.-X.; Tong, G.-Z. Emergence of a Pseudorabies virus variant with increased virulence to piglets. Vet. Microbiol. 2015, 181, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Zeng, X.; Zhang, G.; Wu, Q.; Niu, J.; Sun, B.; Xie, Q.; Ma, J. Molecular characterization and phylogenetic analysis of pseudorabies virus variants isolated from Guangdong Province of Southern China during 2013–2014. J. Vet. Sci. 2015, 17, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qiao, S.; Li, X.; Xie, W.; Guo, J.; Li, Q.; Liu, X.; Hou, J.; Xu, Y.; Wang, L.; et al. Molecular epidemiology of outbreak-associated pseudorabies virus (PRV) strains in central China. Virus Genes 2015, 50, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Zhang, Z.; Lv, L.; Xiao, Y.; Qu, Y.; Ma, H.; Niu, Y.; Wang, G.; Liu, S. Outbreak of variant pseudorabies virus in Bartha-K61-vaccinated piglets in central Shandong Province, China. J. Vet. Diagn. Investig. 2015, 27, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, X.-T.; Hu, B.; Deng, X.-Y.; Zhang, L.; Lian, S.-Z.; Zhang, H.-L.; Lv, S.; Xue, X.-H.; Lu, R.-G.; et al. Outbreak of severe pseudorabies virus infection in pig-offal-fed farmed mink in Liaoning Province, China. Arch. Virol. 2017, 162, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Xu, J.; Song, X.; Jia, R.; Yin, Z.; Cheng, A.; Jia, R.; Zou, Y.; Li, L.; Yin, L.; et al. Antiviral effect of resveratrol in ducklings infected with virulent duck enteritis virus. Antivir. Res. 2016, 130, 93–100. [Google Scholar] [CrossRef] [PubMed]
- De, L.A.; Arena, G.; Stecca, C.; Raciti, M.; Mattia, E. Resveratrol inhibits proliferation and survival of Epstein Barr virus-infected Burkitt’s lymphoma cells depending on viral latency program. Mol. Cancer Res. 2011, 9, 1346–1355. [Google Scholar]
- Docherty, J.J.; Smith, J.S.; Fu, M.M.; Stoner, T.; Booth, T. Effect of topically applied resveratrol on cutaneous herpes simplex virus infections in hairless mice. Antivir. Res. 2004, 61, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Heredia, A.; Davis, C.; Amin, M.N.; Le, N.M.; Wainberg, M.A.; Oliveira, M.; Deeks, S.G.; Wang, L.X.; Redfield, R.R. Targeting Host Nucleotide Biosynthesis with Resveratrol Inhibits Emtricitabine (FTC)-resistant HIV-1. Aids 2014, 28, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Heredia, A.; Davis, C.E.; Reitz, M.S.; Le, N.M.; Wainberg, M.A.; Foulke, J.S.; Wang, L.X.; Redfield, R.R. Targeting of the Purine Biosynthesis Host Cell Pathway Enhances the Activity of Tenofovir Against Sensitive and Drug-Resistant HIV-1. J. Infect. Dis. 2013, 208, 2085–2094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zang, N.; Xie, X.; Deng, Y.; Wu, S.; Wang, L.; Peng, C.; Li, S.; Ni, K.; Luo, Y.; Liu, E. Resveratrol-Mediated Gamma Interferon Reduction Prevents Airway Inflammation and Airway Hyperresponsiveness in Respiratory Syncytial Virus-Infected Immunocompromised Mice. J. Virol. 2011, 85, 13061–13068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, V.; Zeichner, S.L. Host cell gene expression during human immunodeficiency virus type 1 latency and reactivation and effects of targeting genes that are differentially expressed in viral latency. J. Virol. 2004, 78, 9458–9473. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cui, Q.; Fu, Q.; Song, X.; Jia, R.; Yang, Y.; Zou, Y.; Li, L.; He, C.; Liang, X.; et al. Antiviral properties of resveratrol against pseudorabies virus are associated with the inhibition of IκB kinase activation. Sci. Rep. 2017, 7, 8782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Q.; Cui, Q.; Yang, Y.; Zhao, X.; Song, X.; Wang, G.; Bai, L.; Chen, S.; Tian, Y.; Zou, Y.; et al. Effect of Resveratrol Dry Suspension on Immune Function of Piglets. Evid.-Based Complement. Altern. Med. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhang, Y.; Yin, Z.; Zhao, X.; Liang, X.; He, C.; Yin, L.; Lv, C.; Zhao, L.; Ye, G. Antiviral effect of sulfated Chuanmingshen violaceum polysaccharide in chickens infected with virulent Newcastle disease virus. Virology 2015, 476, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yin, Z.; Li, L.; Cheng, A.; Jia, R.; Song, X.; Lu, H.; Dai, S.; Lv, C.; Liang, X. Inhibitory Effect of Resveratrol against Duck Enteritis Virus In Vitro. PLoS ONE 2013, 8, e65213. [Google Scholar] [CrossRef] [PubMed]
- Dory, D.; Torché, A.M.; Béven, V.; Blanchard, P.; Loizel, C.; Cariolet, R.; Jestin, A. Effective protection of pigs against lethal Pseudorabies virus infection after a single injection of low-dose Sindbis-derived plasmids encoding PrV gB, gC and gD glycoproteins. Vaccine 2005, 23, 3483–3491. [Google Scholar] [CrossRef] [PubMed]
- Maresch, C.; Lange, E.; Teifke, J.P.; Fuchs, W.; Klupp, B.; Müller, T.; Mettenleiter, T.C.; Vahlenkamp, T.W. Oral immunization of wild boar and domestic pigs with attenuated live vaccine protects against Pseudorabies virus infection. Vet. Microbiol. 2013, 161, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Hu, Z.; Song, X.; Cui, Q.; Fu, Q.; Jia, R.; Zou, Y.; Li, L.; Yin, Z. Analgesic and Anti-Inflammatory Activities of Resveratrol through Classic Models in Mice and Rats. Evid.-Based Complement. Altern. Med. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Hu, Z.; Fu, Q.; Song, X.; Cui, Q.; Jia, R.; Zou, Y.; He, C.; Li, L.; Yin, Z. Resveratrol mitigates lipopolysaccharide-mediated acute inflammation in rats by inhibiting the TLR4/NF-κBp65/MAPKs signaling cascade. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Biron, C.A. Role of early cytokines, including alpha and beta interferons (IFN-α\β), in innate and adaptive immune responses to viral infections. Semin. Immunol. 1998, 10, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Chesler, D.A.; Reiss, C.S. The role of IFN-gamma in immune responses to viral infections of the central nervous system. Cytokine Growth Factor Rev. 2002, 13, 441–454. [Google Scholar] [CrossRef]
- Su, H.C.; Cousens, L.P.; Fast, L.D.; Slifka, M.K.; Bungiro, R.D.; Ahmed, R.; Biron, C.A. CD4+ and CD8+ T cell interactions in IFN-gamma and IL-4 responses to viral infections: Requirements for IL-2. J. Immunol. 1998, 160, 5007–5017. [Google Scholar] [PubMed]
- Trevejo, J.M.; Marino, M.W.; Philpott, N.; Josien, R.; Richards, E.C.; Elkon, K.B.; Falck-Pedersen, E. TNF-alpha -dependent maturation of local dendritic cells is critical for activating the adaptive immune response to virus infection. Proc. Natl. Acad. Sci. USA 2001, 98, 12162. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.B.; Salazar-Mather, T.P.; Dalod, M.Y.; Van Deusen, J.B.; Wei, X.Q.; Liew, F.Y.; Caligiuri, M.A.; Durbin, J.E.; Biron, C.A. Coordinated and distinct roles for IFN-alpha beta, IL-12, and IL-15 regulation of NK cell responses to viral infection. J. Immunol. 2002, 169, 4279–4287. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Wolcott, R.M.; Chervenak, R.; Jennings, S.R. Control of acute cutaneous Herpes Simplex Virus infection: T cell-ediated viral clearance is dependent upon Interferon gamma. Virology 1994, 202, 76–88. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days Post Infection | Survival Rate (%) | ||||
|---|---|---|---|---|---|
| Res-H | Res-M | Res-L | Untreated | Non-Infected | |
| 1 | 100 | 100 | 100 | 100 | 100 |
| 2 | 100 | 100 | 100 | 100 | 100 |
| 3 | 100 | 100 | 100 | 100 | 100 |
| 4 | 100 | 100 | 100 | 100 | 100 |
| 5 | 100 | 100 | 100 | 100 | 100 |
| 6 | 100 | 100 | 100 | 80 | 100 |
| 7 a | 100 | 100 | 90 | 60 | 100 |
| Group | Res-H | Res-M | Res-L | Untreated |
|---|---|---|---|---|
| Brain | 0.72 ± 0.15 & | 0.69 ± 0.21 & | 3.90 ± 0.36 * | 8.02 ± 0.85 |
| Lung | 3.54 ± 0.06 * | 3.73 ± 0.21 * | 5.74 ± 0.43 | 7.41 ± 1.43 |
| Kidney | 0.65 ± 0.06 & | 0.99 ± 0.10 & | 4.32 ± 0.31 * | 6.39 ± 0.55 |
| Liver | 1.84 ± 0.24 * | 2.01 ± 0.30 * | 4.75 ± 0.23 | 6.33 ± 1.13 |
| Spleen | N.D. | N.D. | 2.28 ± 0.42 * | 5.38 ± 0.86 |
| Heart | 0.88 ± 0.79 * | 0.77 ± 1.33 * | 2.48 ± 0.46 * | 5.33 ± 0.66 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Tong, W.; Song, X.; Jia, R.; Li, L.; Zou, Y.; He, C.; Liang, X.; Lv, C.; Jing, B.; et al. Antiviral Effect of Resveratrol in Piglets Infected with Virulent Pseudorabies Virus. Viruses 2018, 10, 457. https://doi.org/10.3390/v10090457
Zhao X, Tong W, Song X, Jia R, Li L, Zou Y, He C, Liang X, Lv C, Jing B, et al. Antiviral Effect of Resveratrol in Piglets Infected with Virulent Pseudorabies Virus. Viruses. 2018; 10(9):457. https://doi.org/10.3390/v10090457
Chicago/Turabian StyleZhao, Xinghong, Wenzhi Tong, Xu Song, Renyong Jia, Lixia Li, Yuanfeng Zou, Changliang He, Xiaoxia Liang, Cheng Lv, Bo Jing, and et al. 2018. "Antiviral Effect of Resveratrol in Piglets Infected with Virulent Pseudorabies Virus" Viruses 10, no. 9: 457. https://doi.org/10.3390/v10090457
APA StyleZhao, X., Tong, W., Song, X., Jia, R., Li, L., Zou, Y., He, C., Liang, X., Lv, C., Jing, B., Lin, J., Yin, L., Ye, G., Yue, G., Wang, Y., & Yin, Z. (2018). Antiviral Effect of Resveratrol in Piglets Infected with Virulent Pseudorabies Virus. Viruses, 10(9), 457. https://doi.org/10.3390/v10090457