Maize Dwarf Mosaic Virus: From Genome to Disease Management


Abstract
1. Introduction
2. Genome
3. Transmission
4. Symptom
5. Diagnostic Method
6. Control
6.1. Use of Insecticide and Johnsongrass Eradication
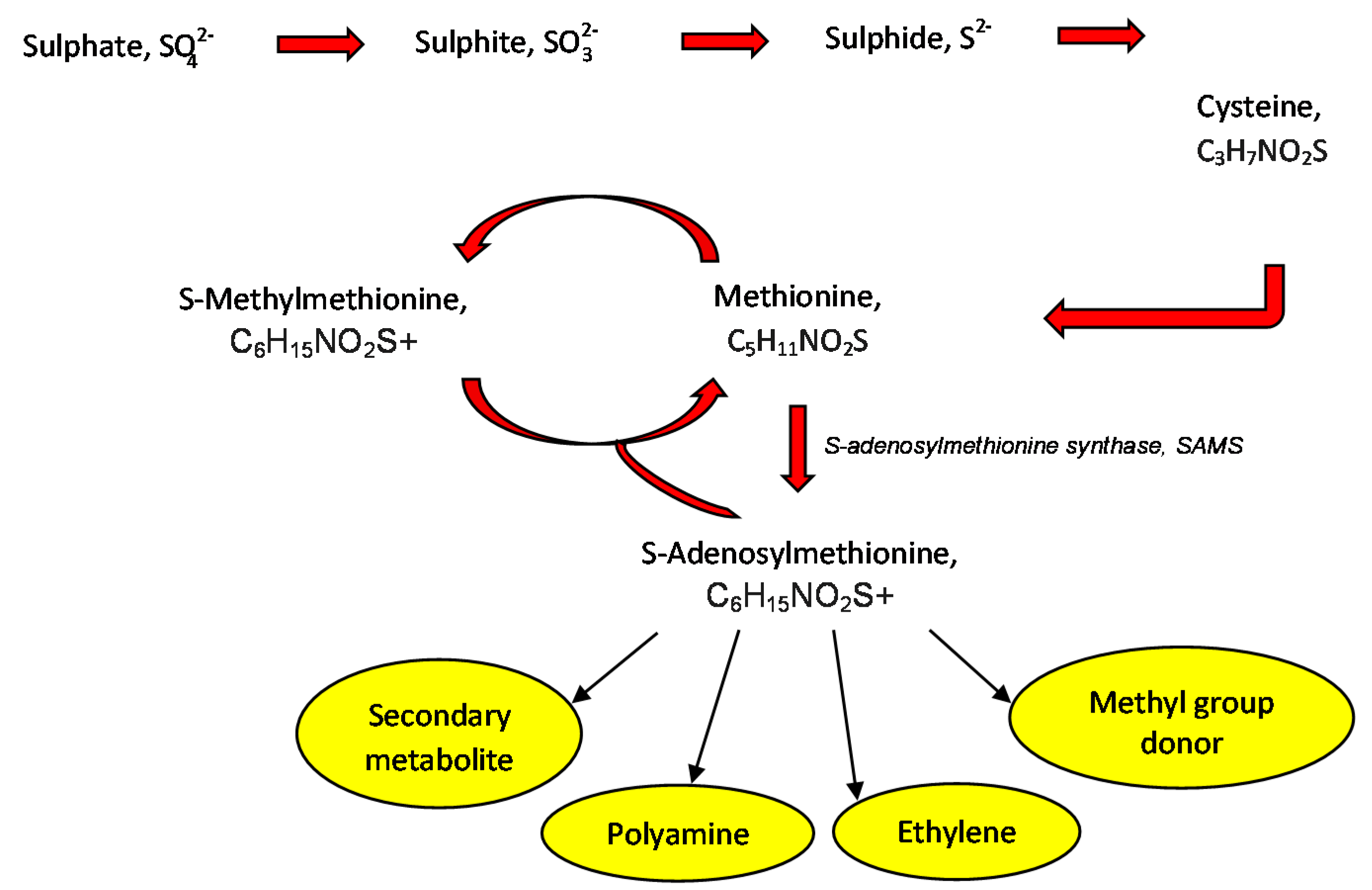
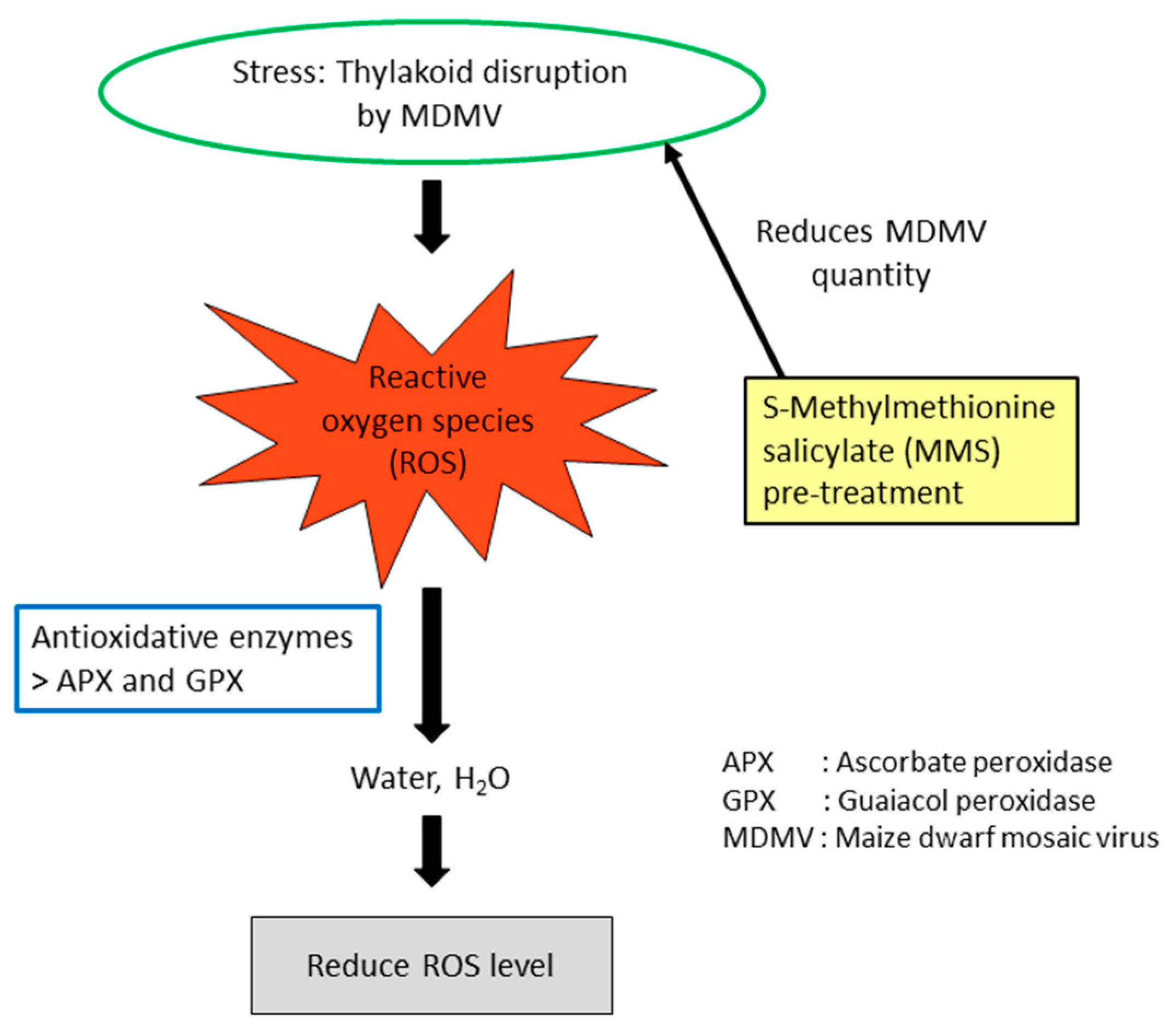
6.2. S-Methylmethionine Pre-Treatment
6.3. Conventional Resistance Breeding
6.4. Genetically Engineered Resistance
7. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ludmerszki, E.; Rácz, I.; Rudnóy, S. S-methylmethionine alters gene expression of candidate genes in Maize dwarf mosaic virus infected and drought stressed maize plants. Acta Biol. Szeged. 2014, 58, 1–5. [Google Scholar]
- Lukuyu, B.; Murdoch, A.; Mcleod, A.; Doward, P. The impact of Maize streak virus disease on quality and yield of maize forage outputs of the maize-dairy production systems in central highlands of Kenya. In Proceedings of the Livestock Development Group Student Conference, Focus on Livelihoods, University of Reading, Reading, UK, 16 October 2004; p. 16. [Google Scholar]
- Redinbaugh, M.G.; Zambrano, J.L. Control of Virus Diseases in Maize. Adv. Virus Res. 2014, 90, 391–429. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organizations. FAO Statistical Yearbook; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; ISBN 9788578110796. [Google Scholar]
- Lapierre, H.; Signoret, P.A. Viruses and Virus Diseases of Poaceae (Gramineae); Institut National de la Recherche Agronomique: Paris, France, 2004; ISBN 2738010881. [Google Scholar]
- Shukla, D.D.; Tosic, M.; Jilka, J.; Ford, R.E.; Toler, R.W.; Langham, M.A.C. Taxonomy of Potyviruses Infecting Maize, Sorghum, and Sugarcane in Australia and the United States as Determined by Reactivities of Polyclonal Antibodies Directed towards Virus-Specific N-Termini of Coat Proteins. Phytopathology 1989, 79, 223. [Google Scholar] [CrossRef]
- Shukla, D.; Ward, C. Structure of Potyvirus Coat Proteins and Its Application in the Taxonomy of the Potyvirus Group. Adv. Virus Res. 1989, 36, 273–314. [Google Scholar] [CrossRef] [PubMed]
- Seifers, D.L.; Salomon, R.; Marie-Jeanne, V.; Alliot, B.; Signoret, P.; Haber, S.; Loboda, A.; Ens, W.; She, Y.M.; Standing, K.G. Characterization of a novel potyvirus isolated from maize in Israel. Phytopathology 2000, 90, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Schurtleff, M.C. Compendium of Corn Diseases; APS Press: St. Paul, MN, USA, 1999; ISBN 0890540292. [Google Scholar]
- Gell, G.; Balázs, E.; Petrik, K. Genetic diversity of Hungarian Maize dwarf mosaic virus isolates. Virus Genes 2010, 40, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.T.; Bradfute, O.E.; Gingery, R.E.; Knoke, J.K.; Louie, R.; Nault, L.R.; Scott, G.E. Introduction: History, geographical distribution, pathogen characteristics, and economic importance. In Viruses and Viruselike Disease of Maize in the United States; Ohio State University: Wooster, OH, USA, 1981; pp. 1–12. [Google Scholar]
- Lübberstedt, T.; Ingvardsen, C.; Melchinger, A.E.; Xing, Y.; Salomon, R.; Redinbaugh, M.G. Two chromosome segments confer multiple potyvirus resistance in maize. Plant Breed. 2006, 125, 352–356. [Google Scholar] [CrossRef]
- Lesemann, D.E.; Shukla, D.D.; Tosic, M.; Huth, W. Differentiation of the four viruses of the Sugarcane mosaic virus subgroup based on cytopathology. Potyvirus Taxon. 1992, 5, 353–361. [Google Scholar] [CrossRef]
- Zielińska, L.; Trzmiel, K.; Jezewska, M. Ultrastructural changes in maize leaf cells infected with Maize dwarf mosaic virus and Sugarcane mosaic virus. Acta Biol. Crac. Ser. Bot. 2012, 54, 97–104. [Google Scholar] [CrossRef]
- Matthews, R. The classification and nomenclature of viruses: Summary of results of meetings of the International Committee on Taxonomy of Viruses in Strasbourg. Intervirology 1982, 16, 53–60. [Google Scholar] [CrossRef]
- Revers, F.; García, J.A. Molecular biology of potyviruses. Adv. Virus Res. 2015, 92, 101–199. [Google Scholar] [CrossRef] [PubMed]
- Tosic, M.; Ford, R.E.; Shukla, D.D.; Jilka, J. Differentiation of Sugarcane, Maize dwarf, Johnsongrass, and Sorghum mosaic viruses based on reactions of oat and some sorghum cultivars. Plant Dis. 1990, 74, 549–552. [Google Scholar] [CrossRef]
- Cao, P.; Eckstein, H.H.; De Rango, P.; Setacci, C.; Ricco, J.B.; de Donato, G.; Becker, F.; Robert-Ebadi, H.; Diehm, N.; Schmidli, J.; et al. Diagnostic Methods. Eur. J. Vasc. Endovasc. Surg. 2011, 42, S13–S32. [Google Scholar] [CrossRef]
- Giolitti, F.; Herrera, M.G.; Madariaga, M.; Lenardon, S.L. Detection of Maize dwarf mosaic virus (MDMV) on maize in Chile. Maydica 2005, 50, 101–104. [Google Scholar]
- Chauhan, R.A. Study of Filamentous Viruses in Maize and Smallgrains. Master’s Thesis, University of Cape Town, Caoe Town, South Africa, 1985. [Google Scholar]
- Gingery, R.E. Chemical and physical properties of maize viruses. South. Coop. Ser. Bull. 1981, 247, 38–39. [Google Scholar]
- Shi, C.L.; Hsu, S.H. Maize dwarf mosaic virus of corn and sorghum in Beijing. Acta Phytopathol. Sin. 1979, 9, 35–40. [Google Scholar]
- Mikel, M.A.; D’Arcy, C.J.; Rhodes, A.M.; Ford, R.E. Yield response of sweet corn to Maize dwarf mosaic virus. Plant Dis. 1981, 65, 900–901. [Google Scholar] [CrossRef]
- Mikel, M.A.; D’Arcy, C.J.; Rhodes, A.M.; Ford, R.E. Yield loss in sweet corn correlated with time of inoculation with Maize dwarf mosaic virus. Plant Dis. 1981, 65, 902–904. [Google Scholar] [CrossRef]
- Gates, D.W.; Gudauskas, R.T. Photosynthesis, respiration and evidence of a metabolic inhibitor in corn infected with Maize dwarf mosaic virus. Phytopathology 1969, 59, 575–580. [Google Scholar]
- Achon, M.A.; Larrañaga, A.; Alonso-Dueñas, N. The population genetics of Maize dwarf mosaic virus in Spain. Arch. Virol. 2012, 157, 2377–2382. [Google Scholar] [CrossRef] [PubMed]
- Kong, P.; Steinbiss, H.H. Complete nucleotide sequence and analysis of the putative poliprotein of Maize dwarf mosaic virus genomic RNA (Bulgarian isolate). Arch. Virol. 1998, 143, 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Urcuqui-Inchima, S.; Haenni, A.L.; Bernardi, F. Potyvirus proteins: A wealth of functions. Virus Res. 2001, 74, 1–2. [Google Scholar] [CrossRef]
- Tuo, D.; Shen, W.; Yan, P.; Li, X.; Zhou, P. Rapid Construction of Stable Infectious Full-Length cDNA Clone of Papaya leaf distortion mosaic virus Using In-Fusion Cloning. Viruses 2015, 7, 6241–6250. [Google Scholar] [CrossRef] [PubMed]
- Marie-Jeanne, V.; Ioos, R.; Peyre, J.; Alliot, B.; Signoret, P. Differentiation of Poaceae potyviruses by reverse transcription-polymerase chain reaction and restriction analysis. J. Phytopathol. 2000, 148, 141–151. [Google Scholar] [CrossRef]
- Shukla, D.D.; Strike, P.M.; Tracy, S.L.; Gough, K.H.; Ward, C.W. The N and C termini of the coat proteins of potyviruses are surface-located and the N terminus contains the major virus-specific epitopes. J. Gen. Virol. 1988, 69, 1497–1508. [Google Scholar] [CrossRef]
- Hill, J.H.; Ford, R.E.; Benner, H.I. Purification and partial characterization of Maize dwarf mosaic virus strain B (Sugarcane mosaic virus). J. Gen. Virol. 1973, 20, 327–339. [Google Scholar] [CrossRef]
- Petrik, K.; Sebestyén, E.; Gell, G.; Balázs, E. Natural insertions within the N-terminal region of the coat protein of Maize dwarf mosaic potyvirus (MDMV) have an effect on the RNA stability. Virus Genes 2010, 40, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Olspert, A.; Chung, B.Y.W.; Atkins, J.F.; Carr, J.P.; Firth, A.E. Transcriptional slippage in the positive-sense RNA virus family Potyviridae. EMBO Rep. 2015, 16, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.Y.W.; Miller, W.A.; Atkins, J.F.; Firth, A.E. An overlapping essential gene in the Potyviridae. Proc. Natl. Acad. Sci. USA 2008, 105, 5897–5902. [Google Scholar] [CrossRef] [PubMed]
- Hillung, J.; Elena, S.F.; Cuevas, J.M. Intra-specific variability and biological relevance of P3N-PIPO protein length in potyviruses. BMC Evol. Biol. 2013, 13, 249. [Google Scholar] [CrossRef] [PubMed]
- Vijayapalani, P.; Maeshima, M.; Nagasaki-Takekuchi, N.; Miller, W.A. Interaction of the trans-frame potyvirus protein P3N-PIPO with host protein PCaP1 facilitates potyvirus movement. PLoS Pathog. 2012, 8, e1002639. [Google Scholar] [CrossRef] [PubMed]
- Masumi, M.; Mostafavi Neishabouri, F.; Nasrollahnejad, S.; Ghahramani, T.; Izadpanah, K. Determination of the Complete Sequence of the Genome of Maize dwarf mosaic virus; Plant Virology Research Center, College of Agriculture, Shiraz University: Shiraz, Iran, 2011. [Google Scholar]
- Achon, M.A.; Serrano, L.; Alonso-Dueñas, N.; Porta, C. Complete genome sequences of Maize dwarf mosaic and Sugarcane mosaic virus isolates coinfecting maize in Spain. Arch. Virol. 2007, 152, 2073–2078. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L.R.; Bouchard, R.; Redinbaugh, M.G.; Meulia, T. Complete sequence and development of a full-length infectious clone of an Ohio isolate of Maize dwarf mosaic virus (MDMV). Virus Res. 2012, 165, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Maize Dwarf Mosaic Virus Isolate Italy, Complete Genome. Available online: https://www.ncbi.nlm.nih.gov/nuccore/402216197 (accessed on 16 August 2018).
- Boyer, J.C.; Haenni, A.L. Infectious Transcripts and cDNA Clones of RNA Viruses. Virology 1994, 198, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Duff-farrier, C.R.A.; Candresse, T.; Bailey, A.M.; Boonham, N.; Foster, G.D. Short Communication Evidence for different, host-dependent functioning of Rx against both wild-type and recombinant Pepino mosaic virus. Mol. Plant Pathol. 2016, 17, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Gell, G.M. Molecular Analysis of the Coat Protein Genes of Maize dwarf mosaic virus (MDMV) Populations. Ph.D. Thesis, University of Szent Istvan, Godollo, Hungary, 2011. [Google Scholar]
- Ali, M.C.; Omar, A.S.; Natsuaki, T. An infectious full-length cDNA clone of Potato virus YNTN-NW, a recently reported strain of PVY that causes potato tuber necrotic ringspot disease. Arch. Virol. 2011, 156, 2039–2043. [Google Scholar] [CrossRef]
- Shepherd, R.J.; Holdeman, Q.L. Seed transmission of the Johnsongrass strain of the Sugarcane mosaic virus in Corn. Plant Dis. Report. 1965, 49, 468–469. [Google Scholar]
- Snazelle, T.E.; Bancroft, J.B.; Ullstrup, A.J. Purification and serology of Maize dwarf mosaic and Sugarcane mosaic viruses. Phytopathology 1971, 61, 1059–1063. [Google Scholar] [CrossRef]
- Langenberg, W.G. Leaf-dip serology for the determination of strain relationships of elongated plant viruses. Phytopathology 1974, 64, 128–131. [Google Scholar] [CrossRef]
- Szirmai, J. The occurrence of stripe mosaic disease of maize in Hungary and possibilities of breeding for virus resistance. Acta Phytopathol. Hung. 1968, 3, 189–198. [Google Scholar]
- Shukla, D.D.; Jilka, J.; Tosic, M.; Ford, R.E. A novel approach to the serology of potyviruses involving affinity-purified polyclonal antibodies directed towards virus-specific N termini of coat proteins. J. Gen. Virol. 1989, 70, 13–23. [Google Scholar] [CrossRef]
- Seifers, D.L.; Hackerott, H.L. Estimates of yield loss and virus titre in sorghum hybrids infected with Maize dwarf mosaic virus strain B. Agric. Ecosyst. Environ. 1987, 19, 81–86. [Google Scholar] [CrossRef]
- Hill, J.H.; Benner, H.I. Properties of potyvirus RNAs: Turnip mosaic, Tobacco etch, and Maize dwarf mosaic viruses. Virology 1976, 75, 419–432. [Google Scholar] [CrossRef]
- Hill, J.H.; Shepherd, R.J. Biochemical properties of Turnip mosaic virus. Virology 1972, 47, 807–816. [Google Scholar] [CrossRef]
- Moury, B.; Fabre, F.; Senoussi, R. Estimation of the number of virus particles transmitted by an insect vector. Proc. Natl. Acad. Sci. USA 2007, 104, 17891–17896. [Google Scholar] [CrossRef] [PubMed]
- D’Arcy, C.J.; Nault, L.R. Insect transmission of plant viruses and mycoplasmalike and rickettsialike organisms. Plant Dis. 1982, 66, 99–104. [Google Scholar] [CrossRef]
- Knoke, J.K.; Louie, R. Epiphytology of maize virus diseases. South. Coop. Ser. Bull. 1981, 247, 92–102. [Google Scholar]
- Diagnostis Methods for Maize dwarf mosaic virus. Available online: http://www.padil.gov.au/pbt (accessed on 4 January 2018).
- Bancroft, J.B.; Ullstrup, A.J.; Messieha, M.; Bracker, C.E.; Snazelle, T.E. Some biological and physical properties of a midwestern isolate of Maize dwarf mosaic virus. Phytopathology 1966, 56, 474–478. [Google Scholar]
- Wang, R.Y.; Pirone, T.P. Potyvirus transmission is not increased by pre-acquisition fasting of aphids reared on artificial diet. J. Gen. Virol. 1996, 77, 3145–3148. [Google Scholar] [CrossRef] [PubMed]
- Berger, P.; Toler, R.; Harris, K. Evidence for 2 modes of retention of Maize dwarf mosaic virus by Schizaphis graminum. Phytopatholog 1983, 73, 787. [Google Scholar]
- Messieha, M. Studies on aphid transmission of Maize dwarf mosaic virus. Phytopathology 1967, 57, 956–959. [Google Scholar]
- Thongmeearkom, P.; Ford, R.E.; Jedlinski, H. Aphid transmission of Maize dwarf mosaic virus strains. Phytopathology 1976, 66, 332–335. [Google Scholar] [CrossRef]
- Salomon, R.; Bernardi, F. Inhibition of viral aphid transmission by the N-terminus of the Maize dwarf mosaic virus coat protein. Virology 1995, 213, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Pirone, T.P.; Blanc, S. Helper-dependent vector transmission of plant viruses. Annu. Rev. Phytopathol. 1996, 34, 227–247. [Google Scholar] [CrossRef] [PubMed]
- Lacomme, C.; Glais, L.; Bellstedt, D.U.; Dupuis, B.; Karasev, A.V.; Jacquot, E. Virus Y: Biodiversity, Pathogenicity, Epidemiology and Management; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–261. [Google Scholar]
- Nault, L.R. Arthropod Transmission of plant viruses: A new synthesis. Ann. Entomol. Soc. Am. 1997, 90, 521–541. [Google Scholar] [CrossRef]
- Atreya, C.D.; Raccah, B.; Pirone, T.P. A point mutation in the coat protein abolishes aphid transmissibility of a potyvirus. Virology 1990, 178, 161–165. [Google Scholar] [CrossRef]
- Jiang, Y.X.; Lei, H.; Collar, J.L.; Martin, B.; Muniz, M.; Fereres, A. Probing and feeding behavior of two distinct biotypes of Bemisia tabaci (Homoptera: Aleyrodidae) on tomato plants. J. Econ. Entomol. 1999, 92, 357–366. [Google Scholar] [CrossRef]
- Wang, R.Y.; Ammar, E.D.; Thornbury, D.W.; Lopez-Moya, J.J.; Pirone, T.P. Loss of potyvirus transmissibility and helper-component activity correlate with non-retention of virions in aphid stylets. J. Gen. Virol. 1996, 77, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.C.; Ford, R.E. Factors affecting aphid transmission of Maize dwarf mosaic virus to corn. Phytopathology 1971, 61, 1516–1521. [Google Scholar] [CrossRef]
- Watson, M.A.; Roberts, F.M. A comparative study of the transmission of Hyoscyamus virus 3, Potato virus Y and Cucumber virus 1 by the vectors Myzus persicae (Sulz), M. circumflexus (Buckton), and Macrosiphum gei (Koch). Proc. R. Soc. Lond. B 1939, 127, 543–576. [Google Scholar] [CrossRef]
- Tsai, J.H.; Brown, L.G. Maize Dwarf Mosaic Virus; Plant Pathology Circular No. 320; Division of Plant Industry: Gainesville, FL, USA, 1989. [Google Scholar]
- Teyssandier, E.E.; Nome, S.F.; Bo, E. Maize virus diseases in Argentina. In Proceedings of the International Maize Virus Disease Colloquium and Workshop, Wooster, OH, USA, 2–6 August 1982; The Ohio State University: Wooster, OH, USA, 1982; pp. 93–99. [Google Scholar]
- Boothroyd, C.W. Seed transmission of Maize dwarf mosaic virus in sweet corn and yield reduction in plants from an infected seed lot. Am. Phytopathol. Soc. 1977, 4, 184. [Google Scholar] [CrossRef]
- Hill, J.H.; Martinson, C.A.; Russell, W.A. Seed Transmission of Maize dwarf mosaic and Wheat streak mosaic viruses in Maize and Response of Inbred Lines. Crop Sci. 1974, 14, 232–235. [Google Scholar] [CrossRef]
- Williams, L.E.; Findley, W.R.; Dollinger, E.J.; Ritter, R.M. Seed transmission studies of Maize dwarf mosaic virus in corn. Plant Dis. Report. 1968, 52, 863–864. [Google Scholar]
- Mikel, M.A.; D’Arcy, C.J.; Ford, R.E. Seed Transmission of Maize dwarf mosaic virus in Sweet Corn. J. Phytopathol. 1984, 110, 185–191. [Google Scholar] [CrossRef]
- Panayotou, P.C. Investigations on seed transmission of Maize dwarf mosaic virus and its effect on the establishment of seedlings. J. Plant Dis. Prot. 1981, 88, 621–625. [Google Scholar]
- Nault, L.R.; Gordon, D.T.; Loayza, J.C. Maize virus and mycoplasma diseases in Peru. Trop. Pest Manag. 1981, 27, 363–369. [Google Scholar] [CrossRef]
- Jones, M.W.; Boyd, E.C.; Redinbaugh, M.G. Responses of maize (Zea mays L.) near isogenic lines carrying Wsm1, Wsm2, and Wsm3 to three viruses in the potyviridae. Theor. Appl. Genet. 2011, 123, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Ford, R.E.; Tosic, M.; Shukla, D.D. Maize Dwarf Mosaic Virus: AAB Descriptions of Plant Viruses Online. 1989. Available online: http://sdb.im.ac.cn/vide/descr465.htm (accessed on 28 January 2018).
- Krass, C.J.; Ford, R.E. Ultrastructure of Corn systemically infected with Maize dwarf mosaic virus. Phytopathology 1969, 59, 431–439. [Google Scholar]
- Chen, S.; Das, P.; Hari, V. In situ localization of ATPase activity in cells of plants infected by Maize dwarf mosaic potyvirus. Arch. Virol. 1994, 134, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Gregory, L.V.; Ayers, J.E. Effect of inoculation with Maize dwarf mosaic virus at several growth stages on yield of sweet corn. Plant Dis. 1982, 66, 801–804. [Google Scholar] [CrossRef]
- Mikel, M.A.; D’Arcy, C.J.; Rhodes, A.M.; Ford, R.E. Effect of Maize dwarf mosaic virus infection on sweet corn pollen and silk. Phytopathology 1982, 72, 428–431. [Google Scholar] [CrossRef]
- Fuchs, E.; Grüntzig, M. Influence of Sugarcane mosaic virus (SCMV) and Maize dwarf mosaic virus (MDMV) on the growth and yield of two maize varieties. J. Plant Dis. Prot. 1995, 102, 44–50. [Google Scholar]
- Williams, M.M.; Pataky, J.K. Interactions between maize dwarf mosaic and weed interference on sweet corn. Field Crop. Res. 2012, 128, 48–54. [Google Scholar] [CrossRef]
- Antignus, Y. Comparative study of two Maize dwarf mosaic virus strains infecting corn and johnsongrass in Israel. Plant Dis. 1987, 71, 687–691. [Google Scholar] [CrossRef]
- Tóbiás, I.; Palkovics, L. An unusual feature at the N-terminal end of the coat protein of Maize dwarf mosaic virus isolated in Hungary. J. Phytopathol. 2004, 152, 445–457. [Google Scholar] [CrossRef]
- Robaglia, C.; Bruening, G.; Haseloff, J.; Gerlach, W.L. Evolution and replication of Tobacco ringspot virus satellite RNA mutants. EMBO J. 1993, 12, 2969–2976. [Google Scholar] [PubMed]
- Jones, F.E.; Hill, J.H.; Durand, D.P. Detection and differentiation of Maize dwarf mosaic virus, strains A and B, by use of different class immunoglobulins in a double-antibody sandwich enzyme-linked immunosorbent assay. Phytopathology 1988, 78, 1118–1124. [Google Scholar] [CrossRef]
- Sum, I.; Németh, M.; Pacsa, A.S. Detection of Maize dwarf mosaic virus with Enzyme-Linked Immunosorbent Assay (ELISA). J. Phytopathol. 1979, 95, 274–279. [Google Scholar] [CrossRef]
- Lenardon, S.L.; Giolitti, F.J. Maize Dwarf Mosaic Potyvirus on Maize in Argentina; IFFIVE, INTA: Cordoba, Argentina, 2006. [Google Scholar]
- Chen, J.; Chen, J.; Adams, M.J. A universal PCR primer to detect members of the Potyviridae and its use to examine the taxonomic status of several members of the family. Arch. Virol. 2001, 146, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.B.; Xing, D.; Zhu, D.B.; Zhou, X.M. High sensitive method detection of plant RNA viruses by Electrochemiluminescence reverse transcription PCR. Prog. Biomed. Opt. Imaging- Proc. SPIE 2007, 6534, 1–8. [Google Scholar] [CrossRef]
- Hill, E.K.; Hill, J.H.; Durand, D.P. Production of monoclonal antibodies to viruses in the potyvirus group: Use in radioimmunoassay. J. Gen. Virol. 1984, 65, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, K.; Uszczyńska-Ratajczak, B.; Majewska, A.; Borodynko-Filas, N. DNA microarray-based detection and identification of bacterial and viral pathogens of maize. J. Plant Dis. Prot. 2017, 124, 577–583. [Google Scholar] [CrossRef]
- Van Slogteren, E.; Van Slogteren, D.H. Serological identification of plant viruses and serological diagnosis of virus diseases of plants. Annu. Rev. Microbiol. 1957, 11, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Siti-Rohana, B.O.; Ahmad, I.B.; Nasuruddin, B.A. Application of protein-A indirect ELISA (PAI-ELISA) for the detection of anti-Smith antibodies in systemic lupus erythematosus patients. World J. Microbiol. Biotechnol. 1995, 11, 206–208. [Google Scholar] [CrossRef] [PubMed]
- Rybicki, E.P.; Von Wechmar, M.B. The serology of the Bromoviruses I. Serological interrelationships of the Bromoviruses. Virology 1981, 109, 391–402. [Google Scholar] [CrossRef]
- Diaco, R.; Lister, R.M.; Hill, J.H.; Durand, D.P. Demonstration of serological relationships among isolates of barley yellow dwarf virus by using polyclonal and monoclonal antibodies. J. Gen. Virol. 1986, 67, 353–362. [Google Scholar] [CrossRef]
- Koenig, R. ELISA in the study of homologous and heterologous reactions of plant viruses. J. Gen. Virol. 1978, 40, 309–318. [Google Scholar] [CrossRef]
- Van Regenmortel, M.H.V. Serology and immunochemistry of plant viruses. Immunol. Commun. 1982, 11, 509–525. [Google Scholar] [CrossRef]
- Ferro, D.N.; Mackenzie, J.D.; Margolies, D.C. Effect of mineral oil and a systemic insecticide on field spread of aphid-borne maize dwarf mosaic virus in sweet corn. J. Econ. Entomol. 1980, 73, 730–735. [Google Scholar] [CrossRef]
- Ismail, B.S.; Ingon, D.; Omar, O. Effects of metolachlor on activities of enzymes in a Malaysian soil. J. Environ. Sci. Health Part B 1996, 31, 1267–1278. [Google Scholar] [CrossRef]
- Toler, R.W. Maize dwarf mosaic, the most important virus disease of sorghum. Plant Dis. 1985, 69, 1011–1015. [Google Scholar] [CrossRef]
- Gordon, D.T.; Bradfute, O.E.; Gingery, R.E.; Knoke, J.K.; Nault, L.R.; Loden, H.D.; Wilkinson, D. Maize virus disease complexes in the United States: Real and potential disease problems. In 33rd Annual Corn and Sorghum Research Conference; American Seed Trade Association: Washington, DC, USA, 1978; pp. 102–133. [Google Scholar]
- Knoke, J.K.; Louie, R.; Madden, L.V.; Gordon, D.T. Spread of Maize dwarf mosaic virus from johnsongrass to corn. Plant Dis. 1983, 67, 367–370. [Google Scholar] [CrossRef]
- Yossen, V.; Nome, S.F.; Latorre, B.A.; Conci, L.R. Occurrence of Maize dwarf mosaic virus (MDMV) in Johnson grass (Sorghum halepense (L.) Pers.) in Chile. Cienc. Investig. Agrar. 1988, 15, 39–45. [Google Scholar]
- Angeles Achon, M.; Sobrepere, M. Incidence of potyviruses in commercial maize fields and their seasonal cycles in Spain. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz 2001, 108, 399–406. [Google Scholar]
- Stewart, L.R.; Teplier, R.; Todd, J.C.; Jones, M.W.; Cassone, B.J.; Wijeratne, S.; Wijeratne, A.; Redinbaugh, M.G. Viruses in Maize and Johnsongrass in Southern Ohio. Phytopathology 2014, 104, 1360–1369. [Google Scholar] [CrossRef] [PubMed]
- Hruska, A.J.; Gladstone, S.M.; Obando, R. Epidemic roller coaster: Maize stunt disease in Nicaragua. Am. Entomol. 1996, 42, 248–252. [Google Scholar] [CrossRef]
- Shepherd, D.N.; Martin, D.P.; Van der Walt, E.; Dent, K.; Varsani, A.; Rybicki, E.P. Maize streak virus: An old and complex ‘emerging’ pathogen. Mol. Plant Pathol. 2010, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Leustek, T. Repression of cystathionine γ-synthase in Arabidopsis thaliana produces partial methionine auxotrophy and developmental abnormalities. Plant Sci. 2000, 151, 9–18. [Google Scholar] [CrossRef]
- Hacham, Y.; Avraham, T.; Amir, R. The N-terminal region of Arabidopsis cystathionine γ-synthase plays an important regulatory role in methionine metabolism. Plant Physiol. 2002, 128, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Ludmerszki, E.; Paldi, K.; Racz, I.; Szigeti, Z.; Rudnoy, S.Z. The promising role of exogenous s-methylmethionine in agriculture, in the case of maize cultivation. Appl. Ecol. Environ. Res. 2014, 12, 777–785. [Google Scholar] [CrossRef]
- Ranocha, P.; McNeil, S.D.; Ziemak, M.J.; Li, C.; Tarczynski, M.C.; Hanson, A.D. The S-methylmethionine cycle in angiosperms: Ubiquity, antiquity and activity. Plant J. 2001, 25, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.; Eliot, A.C.; Kirsch, J.F. S-methylmethionine is both a substrate and an inactivator of 1-aminocyclopropane-1-carboxylate synthase. Arch. Biochem. Biophys. 2004, 421, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Ludmerszki, E.; Rudnóy, S.; Almási, A.; Szigeti, Z.; Rácz, I. The beneficial effects of S-methyl-methionine in maize in the case of Maize dwarf mosaic virus infection. Acta Biol. Szeged. 2011, 55, 109–112. [Google Scholar]
- Kocsis, M.G.; Nolte, K.D.; Rhodes, D.; Shen, T.L.; Gage, D.A.; Hanson, A.D. Dimethylsulfoniopropionate Biosynthesis inSpartina alterniflora 1: Evidence That S-Methylmethionine and Dimethylsulfoniopropylamine Are Intermediates. Plant Physiol. 1998, 117, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Ludmerszki, E.; Almasi, A.; Racz, I.; Szigeti, Z.; Solti, Á.; Olah, C.; Rudnoy, S. S-methylmethionine contributes to enhanced defense against Maize dwarf mosaic virus infection in maize. Braz. J. Bot. 2015, 38, 771–782. [Google Scholar] [CrossRef]
- Asada, K. Radical production and scavenging in the chloroplasts. Photosynth. Environ. 1996, 123–150. [Google Scholar] [CrossRef]
- Kingston-Smith, A.H.; Foyer, C.H. Bundle sheath proteins are more sensitive to oxidative damage than those of the mesophyll in maize leaves exposed to paraquat or low temperatures. J. Exp. Bot. 2000, 51, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Fryer, M.J.; Andrews, J.R.; Oxborough, K.; Blowers, D.A.; Baker, N.R. Relationship between CO2 assimilation, photosynthetic electron transport, and active O2 metabolism in leaves of maize in the field during periods of low temperature. Plant Physiol. 1998, 116, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Ludmerszki, E.; Chounramany, S.; Oláh, C.; Kátay, G.; Rácz, I.; Almási, A.; Solti, Á.; Bélai, I.; Rudnóy, S. Protective role of S-methylmethionine-salicylate in maize plants infected with Maize dwarf mosaic virus. Eur. J. Plant Pathol. 2017, 149, 145–156. [Google Scholar] [CrossRef]
- Mayhew, D.E.; Ford, R.E. Detection of ribonuclease-resistant RNA in chloroplasts of corn leaf tissue infected with Maize dwarf mosaic virus. Virology 1974, 57, 503–509. [Google Scholar] [CrossRef]
- Wei, T.; Huang, T.S.; McNeil, J.; Laliberté, J.F.; Hong, J.; Nelson, R.S.; Wang, A. Sequential recruitment of the endoplasmic reticulum and chloroplasts for plant potyvirus replication. J. Virol. 2010, 84, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Beddard, G.S.; Porter, G. Concentration quenching in chlorophyll. Nature 1976, 260, 366–367. [Google Scholar] [CrossRef]
- Scott, G.E.; Findley, W.R. Breeding for resistance to maize virus diseases: A challenge. Maize Virus Dis. Newsl. 1984, 1984, 12–14. [Google Scholar]
- Martin, I.F.; McCarthy, T.E.; Persely, D.M.; Greber, R.S. Breeding sweet corn for resistance to Sugarcane mosaic virus—Johnsongrass strain in Australia. Maize Virus Dis. Newsl. 1984, 1, 15–16. [Google Scholar]
- Kovács, G.; Milinkó, I.; Gyulavári, O.; Farády, L. Resistance of F1, F2 and BC1generations to Maize dwarf mosaic virus in maize. Arch. Phytopathol. Plant Prot. 1990, 26, 363–368. [Google Scholar] [CrossRef]
- Chahal, G.S.; Gosal, S.S. Principles and Procedures of Plant Breeding: Biotechnological and Conventional Approaches; CRC Press: Boca Raton, FL, USA, 2002; ISBN 084931321X. [Google Scholar]
- Lei, J.D.; Agrios, G.N. Mechanisms of resistance in corn to Maize dwarf mosaic virus. Phytopathology 1986, 76, 1034–1040. [Google Scholar] [CrossRef]
- Louie, R. Effects of genotype and inoculation protocols on resistance evaluation of maize to Maize dwarf mosaic virus strains. Phytopathology 1986, 76, 769–773. [Google Scholar] [CrossRef]
- Findley, W.R.; Dollinger, E.J.; Louie, R.; Knoke, J.K. Locating Genes for Maize Dwarf Mosaic Resistance by Means of Chromosomal Translocations in Corn (Zea mays L.) 1. Crop Sci. 1973, 13, 608–611. [Google Scholar] [CrossRef]
- Scott, G.E.; Rosenkranz, E. Effectiveness of resistance to Maize dwarf mosaic and Maize chlorotic dwarf viruses in maize. Phytopathology 1981, 71, 937–941. [Google Scholar] [CrossRef]
- Scott, G.E. Linkage between Maize dwarf mosaic virus resistance and endosperm color in maize. Crop Sci. 1989, 29, 1478–1480. [Google Scholar] [CrossRef]
- McMullen, M.D.; Louie, R. Identification of a gene for resistance to Wheat streak mosaic virus in maize. Phytopathology 1991, 81, 624–627. [Google Scholar] [CrossRef]
- Ng, C.H.; Wickneswari, R.; Salmijah, S.; Teng, Y.T.; Ismail, B.S. Gene polymorphisms in glyphosate-resistant and -susceptible biotypes of Eleusine indica from Malaysia. Weed Res. 2003, 43, 108–115. [Google Scholar] [CrossRef]
- McMullen, M.D.; Louie, R. The linkage of molecular markers to a gene controlling the symptom response in maize to Maize dwarf mosaic virus. Mol. Plant-Microbe Interact. 1989, 2, 309–311. [Google Scholar] [CrossRef]
- Jones, M.W.; Redinbaugh, M.G.; Louie, R. The Mdm1 locus and maize resistance to Maize dwarf mosaic virus. Plant Dis. 2007, 91, 185–190. [Google Scholar] [CrossRef]
- Louie, R.; Findley, W.R.; Knoke, J.K.; McMullen, M.D. Genetics basis of resistance in maize to five Maize dwarf mosaic virus strains. Crop Sci. 1991, 31, 14–18. [Google Scholar] [CrossRef]
- Zambrano, J.L.; Jones, M.W.; Brenner, E.; Francis, D.M.; Tomas, A.; Redinbaugh, M.G. Genetic analysis of resistance to six virus diseases in a multiple virus-resistant maize inbred line. Theor. Appl. Genet. 2014, 127, 867–880. [Google Scholar] [CrossRef] [PubMed]
- Pataky, J.K.; Murphy, J.F.; D’arcy, C.J. Resistance to Maize dwarf mosaic virus, severity of symptoms, titer of virus, and yield of sweet corn. Plant Dis. 1990, 74, 359–364. [Google Scholar] [CrossRef]
- Scott, G.E.; Rosenkranz, E. Variable Reaction within Corn Inbreds to Maize dwarf mosaic virus not Genetically Controlled1. Crop Sci. 1987, 27, 78–79. [Google Scholar] [CrossRef]
- Jones, R.K.; Tolin, S.A. Concentration of Maize dwarf mosaic virus in susceptible and resistant corn hybrids. Phytopathology 1972, 62, 640–644. [Google Scholar] [CrossRef]
- Kuntze, L.; Fuchs, E.; Grüntzig, M.; Schulz, B.; Klein, D.; Melchinger, A.E. Resistance of early-maturing European maize germplasm to Sugarcane mosaic virus (SCMV) and Maize dwarf mosaic virus (MDMV). Plant Breed. 1997, 116, 499–501. [Google Scholar] [CrossRef]
- Law, M.D.; Moyer, J.W.; Payne, G.A. The effect of host resistance on pathogenesis of Maize dwarf mosaic virus. Phytopathology 1989, 79, 757–761. [Google Scholar] [CrossRef]
- Azad, M.; Kalam, A.; Amin, L.; Sidik, N.M. Gene technology for Papaya ringspot virus disease management. Sci. World J. 2014, 2014, 768038. [Google Scholar] [CrossRef] [PubMed]
- Prins, M.; Laimer, M.; Noris, E.; Schubert, J.; Wassenegger, M.; Tepfer, M. Strategies for antiviral resistance in transgenic plants. Mol. Plant Pathol. 2008, 9, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Louie, R. Vascular puncture of maize kernels for the mechanical transmission of Maize white line mosaic virus and other viruses of maize. Phytopathology 1995, 85, 139–143. [Google Scholar] [CrossRef]
- Atabekov, J.G.; Dorokhov, Y.L. Plant virus-specific transport function and resistance of plants to viruses. Adv. Virus Res. 1984, 29, 313–364. [Google Scholar] [CrossRef] [PubMed]
- Bennetzen, J.L.; Hake, S.C. Handbook of Maize: Its Biology; Springer: New York, NY, USA, 2009; pp. 121–126. ISBN 978-0-387-79417-4e. [Google Scholar]
- Cassone, B.J.; Chen, Z.; Chiera, J.; Stewart, L.R.; Redinbaugh, M.G. Responses of highly resistant and susceptible maize to vascular puncture inoculation with Maize dwarf mosaic virus. Physiol. Mol. Plant Pathol. 2014, 86, 19–27. [Google Scholar] [CrossRef]
- Xing, Y.; Ingvardsen, C.; Salomon, R.; Lübberstedt, T. Analysis of Sugarcane mosaic virus resistance in maize in an isogenic dihybrid crossing scheme and implications for breeding potyvirus-resistant maize hybrids. Genome 2006, 49, 1274–1282. [Google Scholar] [CrossRef] [PubMed]
- Johal, G.S.; Briggs, S.P. Reductase activity encoded by the HM1 disease resistance gene in maize. Science 1992, 258, 985–987. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.B.; Brommonschenkel, S.H.; Chunwongse, J.; Frary, A.; Ganal, M.W.; Spivey, R.; Tanksley, S.D. Map-based cloning of a protein kinase gene conferring disease resistance in tomato. Science 1993, 262, 1432–1436. [Google Scholar] [CrossRef] [PubMed]
- Simcox, K.D.; McMullen, M.D.; Louie, R. Co-segregation of the Maize dwarf mosaic virus resistance gene, Mdm1, with the nucleolus organizer region in maize. Theor. Appl. Genet. 1995, 90, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.Y.; Zhang, S.H.; Li, X.H.; Xie, C.X.; Li, M.S.; Hao, Z.F.; Zhang, D.G.; Liang, Y.H.; Bai, L.; Zhang, S.H. Identification and mapping of a novel Sugarcane mosaic virus resistance gene in maize. Acta Agron. Sin. 2008, 34, 1494–1499. [Google Scholar] [CrossRef]
- Liu, X.; Tan, Z.; Li, W.; Zhang, H.; He, D. Cloning and transformation of SCMV CP gene and regeneration of transgenic maize plants showing resistance to SCMV strain MDB. Afr. J. Biotechnol. 2009, 8, 3747–3753. [Google Scholar]
- Murry, L.E.; Elliott, L.G.; Capitant, S.A.; West, J.A.; Hanson, K.K.; Scarafia, L.; Johnston, S.; DeLuca-Flaherty, C.; Nichols, S.; Cunanan, D.; et al. Transgenic corn plants expressing MDMV strain B coat protein are resistant to mixed infections of Maize dwarf mosaic virus and Maize chlorotic mottle virus. Nat. Biotechnol. 1993, 11, 1559–1564. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Fu, F.L.; Gou, L.; Wang, H.G.; Li, W.C. RNA interference-based transgenic maize resistant to Maize dwarf mosaic virus. J. Plant Biol. 2010, 53, 297–305. [Google Scholar] [CrossRef]
- Behboodian, B.; Ali, Z.M.; Ismail, I.; Zainal, Z. Postharvest analysis of lowland transgenic tomato fruits harboring hpRNAi-ACO1 construct. Sci. World J. 2012, 2012, 439870. [Google Scholar] [CrossRef] [PubMed]
- Fahim, M.; Ayala-Navarrete, L.; Millar, A.A.; Larkin, P.J. Hairpin RNA derived from viral NIa gene confers immunity to Wheat streak mosaic virus infection in transgenic wheat plants. Plant Biotechnol. J. 2010, 8, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Praveen, S.; Ramesh, S.V.; Mishra, A.K.; Koundal, V.; Palukaitis, P. Silencing potential of viral derived RNAi constructs in Tomato leaf curl virus-AC4 gene suppression in tomato. Transgenic Res. 2010, 19, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Yang, L.; Zhou, S.F.; Wang, H.G.; Li, W.C.; Fu, F.L. Improvement of resistance to maize dwarf mosaic virus mediated by transgenic RNA interference. J. Biotechnol. 2011, 153, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.K.; Lohuis, D.; Goldbach, R.; Prins, M. High frequency induction of RNA-mediated resistance against Cucumber mosaic virus using inverted repeat constructs. Mol. Breed. 2004, 14, 215–226. [Google Scholar] [CrossRef]
- Hily, J.M.; Ravelonandro, M.; Damsteegt, V.; Bassett, C.; Petri, C.; Liu, Z.; Scorza, R. Plum pox virus coat protein gene Intron-hairpin-RNA (ihpRNA) constructs provide resistance to Plum pox virus in Nicotiana benthamiana and Prunus domestica. J. Am. Soc. Hortic. Sci. 2007, 132, 850–858. [Google Scholar]
- Zhang, Z.Y.; Wang, Y.G.; Shen, X.J.; Li, L.; Zhou, S.F.; Li, W.C.; Fu, F.L. RNA interference-mediated resistance to Maize dwarf mosaic virus. Plant Cell Tissue Organ Cult. 2013, 113, 571–578. [Google Scholar] [CrossRef]
- Redinbaugh, M.G.; Jones, M.W.; Gingery, R.E. The genetics of virus resistance in maize (Zea mays L.). Maydica 2004, 49, 183–190. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MDMV Isolates | Geographical Origin | Genbank Accession | References |
|---|---|---|---|
| MDMV | Golestan | JQ280313 | [38] |
| MDMV-Bg | Bulgaria | NC003377 | [27] |
| MDMV-Sp | Spain | AJ416645 | [39] |
| MDMV-OH1 | Ohia | JQ403608 | [40] |
| MDMV-OH2 | Ohia | JQ403609 | [40] |
| MDMV-It | Italy | JX185302 | [41] |
| Method | Platform | Effect on MDMV Variation | Reliability | Efficiency | References |
|---|---|---|---|---|---|
| Indirect Enzyme-Linked Immunosorbent Assay (ELISA) |
| MDMV-A and MDMV-B | Allow detection of MDMV-A and MDMV-B bound IgG among tested leaves sap infected with several different strains of MDMV and Sugarcane mosaic virus (SCMV). | MDMV-A-specific ELISA = antigen detected in sap at a dilution of 1:60 with optimal sample pH (7.5–8.5). MDMV-B-specific ELISA = detect antigen in sap at dilution end point of 1:2560 | [91] |
| Indirect Enzyme-Linked Immunosorbent Assay (ELISA) |
| M-C and M-D | M-C antigen reacts strongly with M-A, M-D, Sorghum red stripe virus (SRV) antisera. M-D antigen reacts strongly with M-D, M-C antisera. | M-C particles react with M-A and SRV antisera in the range of 1:128 to 1:512 dilution end points, with M-D antisera at dilution end point of 1:4. M-D antigen reacted with homologous antisera, M-D up to a dilution of 1:512, with M-C antisera at 1:16 dilution. | [88] |
| Double-Antibody Sandwich Enzyme-Linked Immunosorbent Assay (DAS ELISA) |
| MDMV-A | Visual evaluation at absorbance of 405 nm gives reliable information on MDMV-A presence in leaves extracts tested. | Visible yellow colour formed in wells with antigen diluted up to 10−4. Sensitivity is 100 times better than conventional infectivity test. | [92] |
| Double-Antibody Sandwich Enzyme-Linked Immunosorbent Assay (DAS ELISA) |
| MDMV-A, MVMV-J, MDMV-L, MDMV-SP, MDMV-YU | Among all reference strains, only MDMV strains react positively in DAS-ELISA (recorded OD 405 nm values at least twice the healthy sap OD 405 nm values) with anti-MDMV IgG. | IgG antibody dilution to 1 μg/mL able to detect MDMV antigen dilution to 1/100. | [30] |
| Double-Antibody Sandwich Enzyme-Linked Immunosorbent Assay (DAS ELISA) |
| MDMV | DAS-ELISA absorbance values of infected maize leaf samples from field grown plants and maize leaf sample with MDMV-Arg isolate were highly significant while the values from healthy control and buffer were very low. | MDMV-Arg specific polyclonal IgG strongly reactive up to 1:2000 dilution with MDMV antigen from infected field, Chile. | [19] |
| Capture Reverse Transcription-Polymerase Chain Reaction (RT-PCR) |
| MDMV | Single band of expected size (1104 bp) obtained. | Detect MDMV samples from Chile which closely related to MDMV-Arg (Argentina strain) efficiently. | [19] |
| Reverse Transcription-Polymerase Chain Reaction (RT-PCR) |
| MDMV-SP, MDMV-Bu, MDMVJIL | Single product of expected size (327 nts) obtained for all tested MDMV isolates. | Efficient for dealing with well-characterized strains, field collected isolates. | [30] |
| Reverse Transcription-Polymerase Chain Reaction (RT-PCR) |
| MDMV | cDNA fragments of expected size were amplified from the 3′ terminus of RNA genomes of 21 different viruses under family potyviridae including MDMV. | Universal primer designed based on an alignment of the amino acid sequences around the conserved GNNSGQP motif in nuclear inclusion body b (NIb) gene of family potyviridae members. Hence, it is proved useful for detection of potyviridae members. | [94] |
| Reverse Transcription-Polymerase Chain Reaction (RT-PCR) |
| MDMV | Supposed to yield a PCR product of expected size (336 bp) | MDMV can be distinguishable from Sugarcane mosaic virus, SCMV and Johnsongrass mosaic virus, JGMV through RT-PCR | [57] |
| Combined Reverse Transcription Polymerase Chain Reaction (RT-PCR) with Electrochemiluminescence method |
| MDMV | PCR yielded a product with a single band of expected size (643 bp) for all 4 tested viruses including MDMV. | This method has higher sensitivity and lower cost than others. It can effectively detect the MDMV with simplicity and stability. | [95] |
| Competitive Radioimmunoassay (RIA) |
| MDMV-A (referred here as MDMV-AP) and MDMV-B | Feasible alternative to the use of polyclonal antisera in detecting homologous viruses (MDMV, Soybean mosaic, SMV, Lettuce mosaic virus, LMV). | Antigen (purified virus) detected at dilution of 10–50 ng/mL. | [96] |
| Dot Blot Immunoassay |
| MDMV | MDMV symptomatic field grown plants had strong reaction with the polyclonal antiserum against MDMV-Arg isolate while healthy plants were negative. | MDMV samples reacted with MDMV-Arg polyclonal antiserum of dilution up to 1:5000 | [19] |
| Sodium dodecyl sulfate (SDS) immunodiffusion test |
| MDMV-C and MDMV-D | M-C antiserum reacted with both M-C and M-D antigens forming a spur, which indicate partial serological relatedness. M-D antiserum reacted with its homologous viral antigen, M-D. | No precipitin lines were obtained when antisera reacted against healthy crude sap | [88] |
| DNA Microarray (Maizepath)-based detection | Microarray with 60-mer oligonucleotide probes complementary to genomes of 5 maize pathogens including MDMV | MDMV | Obtained results indicate that the fluorescence signals from MDMV, other pathogens and control probes are well distinguished in all performed experiments. | Gives more than 180 K probes in total, thereby classified as high-density microarray that able to investigate thousands of genomic loci in a high-resolution manner. | [97] |
| Chromosome | Locus | Resistance Source | Screening Method | Level of Resistance | References |
|---|---|---|---|---|---|
| 6 (short arm) | Mdm1 | Pa405 Oh1V1 | Mechanical inoculation/Greenhouse Mechanical inoculation/Field | High resistance (Dominant gene) | [140,143] |
| 6 | Mdm1 co-localizes with Wsm1 | Wsm1 NIL (near isogenic lines) Oh28SS/RR/SS Left: Wsm2 alleles Middle: Mdm1/Wsm1 alleles Right: Wsm3 (Two Mdm1/Wsm1 allele) | Mechanical inoculation/Greenhouse/Field condition | High resistance | [80] |
| 6 | Mdm1 co-localizes with Wsm1 | Wsm1 × Oh28 F1 Oh28SS/RS/SS Left: Wsm2 alleles Middle: Mdm1/Wsm1 alleles Right: Wsm3 (One Mdm1/Wsm1 allele) | Mechanical inoculation/Greenhouse/Field condition | Intermediate resistance | [80] |
| 3 & 6 | Wsm2 combine with Mdm1/Wsm1 | Wsm1 Wsm2 NIL Oh28RR/RR/SS Left: Wsm2 alleles Middle: Mdm1/Wsm1 alleles Right: Wsm3 | Mechanical inoculation/Greenhouse/Field condition | Lowers symptom incidence | [80,142] |
| 3 & 6 | Wsm2 combine with Mdm1/Wsm1 | Wsm1 Wsm2 × Oh28 F1 Oh28RS/RS/SS Left: Wsm2 alleles Middle: Mdm1/Wsm1 alleles Right: Wsm3 | Mechanical inoculation/Greenhouse/Field condition | Lowers incidence and severity of disease | [80,142] |
| 6 & 10 | Wsm3 combine with Mdm1/Wsm1 | Wsm1 Wsm3 × Oh28 F1 Oh28SS/RS/RS Left: Wsm2 alleles Middle: Mdm1/Wsm1 alleles Right: Wsm3 | Mechanical inoculation/Greenhouse/Field condition | Lowers disease incidence and severity | [80,141] |
| 3 & 6 | Scmv2 combined with Scmv1 | F7 RR/RR Left: Scmv2 alleles Right: Scmv1 alleles R: Susceptible parent F7 R: Resistant parent FAP1360A | Mechanical inoculation by rubbing infected leaves | Complete resistance | [155] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kannan, M.; Ismail, I.; Bunawan, H. Maize Dwarf Mosaic Virus: From Genome to Disease Management. Viruses 2018, 10, 492. https://doi.org/10.3390/v10090492
Kannan M, Ismail I, Bunawan H. Maize Dwarf Mosaic Virus: From Genome to Disease Management. Viruses. 2018; 10(9):492. https://doi.org/10.3390/v10090492
Chicago/Turabian StyleKannan, Maathavi, Ismanizan Ismail, and Hamidun Bunawan. 2018. "Maize Dwarf Mosaic Virus: From Genome to Disease Management" Viruses 10, no. 9: 492. https://doi.org/10.3390/v10090492
APA StyleKannan, M., Ismail, I., & Bunawan, H. (2018). Maize Dwarf Mosaic Virus: From Genome to Disease Management. Viruses, 10(9), 492. https://doi.org/10.3390/v10090492