Eradication of Vancomycin-Resistant Enterococci by Combining Phage and Vancomycin


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial and Phage Strains
2.2. Materials
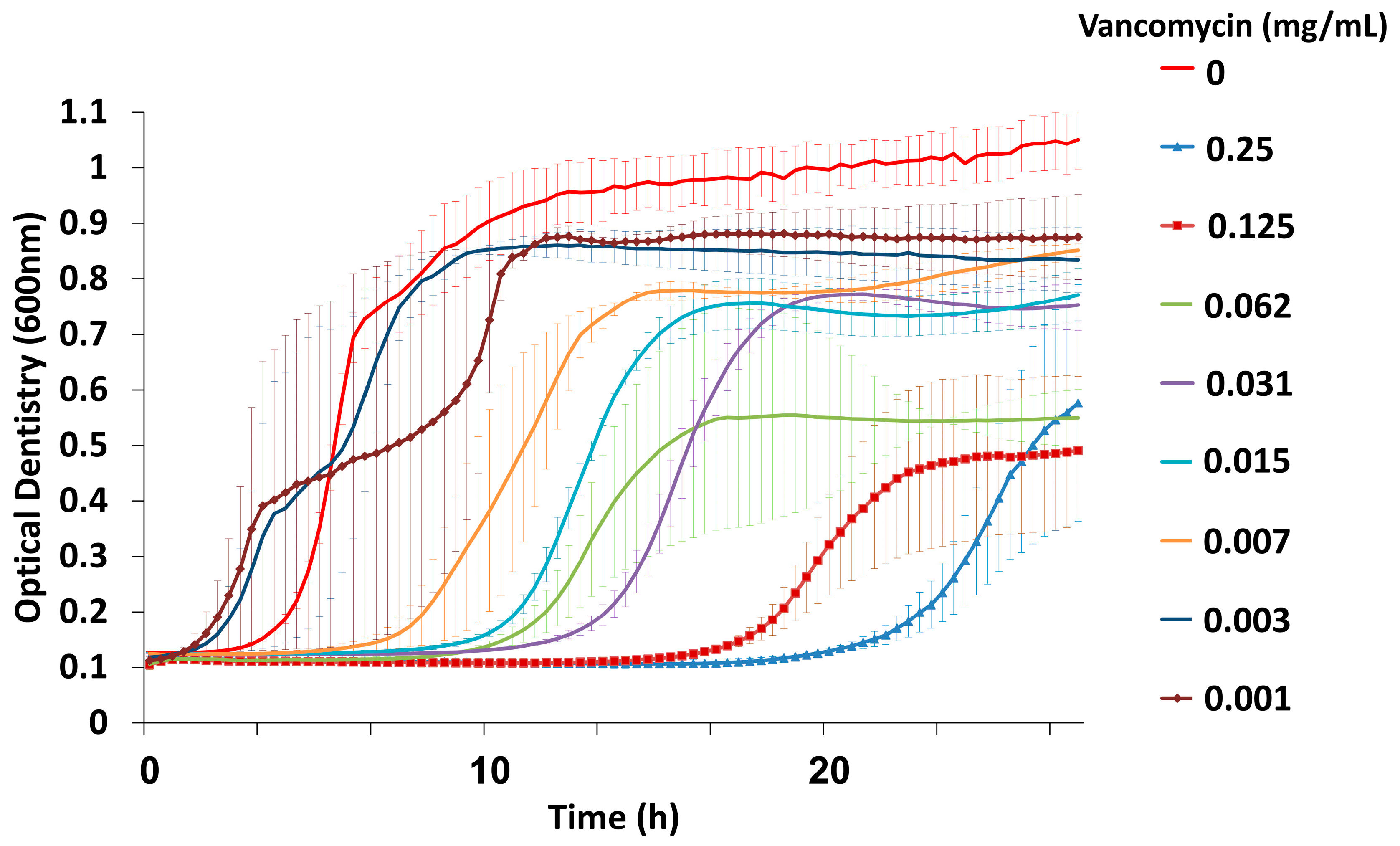
2.3. Determination of Vancomycin Minimal Inhibitory Concentration (MIC)
2.4. Host-Range Specificity Tests
2.5. Assessment of Phage and Antibiotic Lytic Activity in Planktonic Cultures
2.6. Assessment of Phage Lytic Activity in a Biofilm
2.7. Scanning Electron Microscopy (SEM)
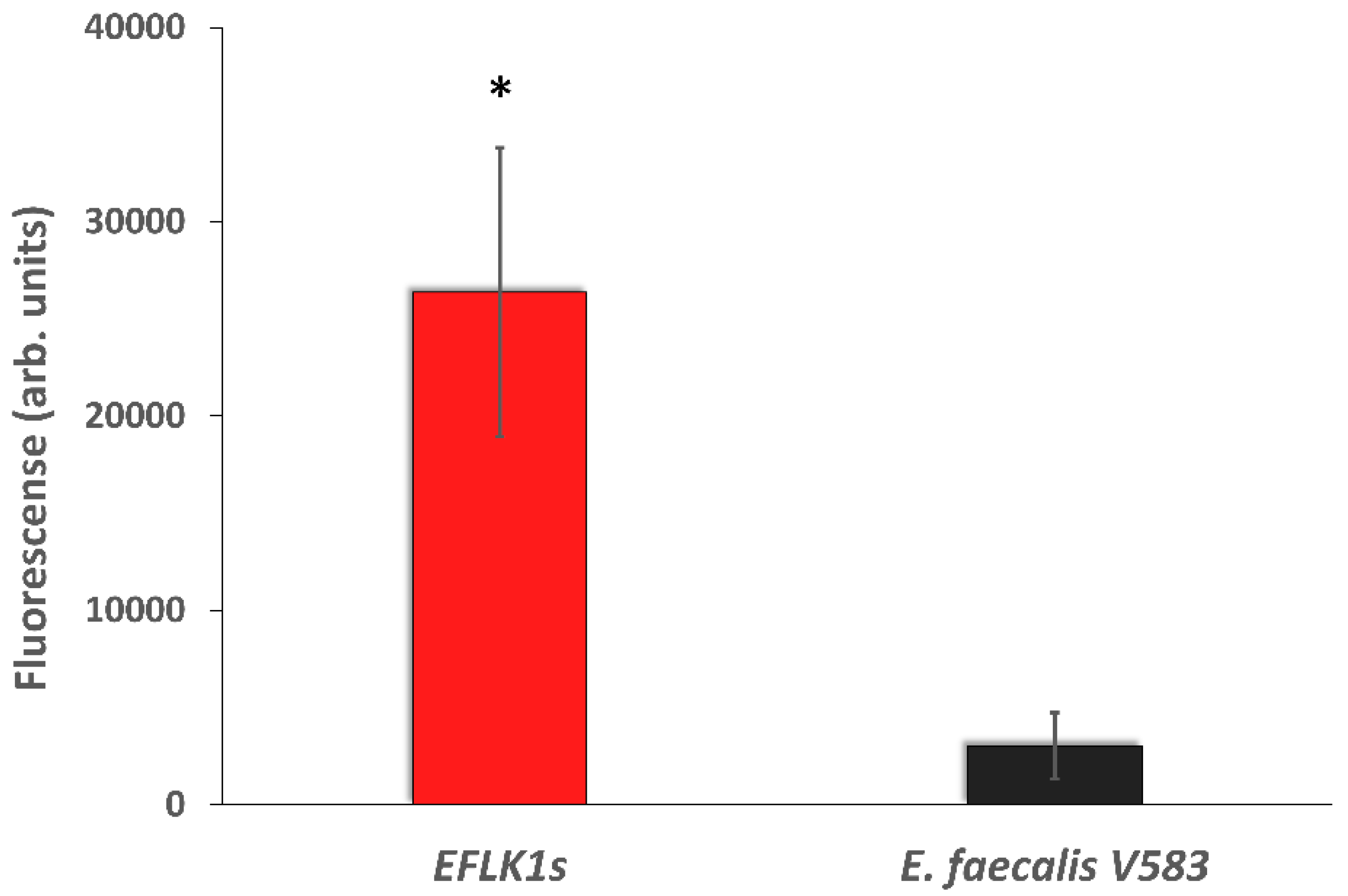
2.8. Detection of N-Acetylglucosamine in Cell-Wall
2.9. Statistical Analysis
3. Results
3.1. Combined Treatment with Vancomycin and Phage EFLK1 Reduced VRE Planktonic Growth
3.2. Combined Treatment of Vancomycin and Phage EFLK1 Reduced 72 h Biofilms
3.3. Identifying Cell Wall Changes Followed Phage Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bereket, W.; Hemalatha, K.; Getenet, B.; Wondwossen, T.; Solomon, A.; Zeynudin, A.; Kannan, S. Update on bacterial nosocomial infections. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 1039–1044. [Google Scholar] [PubMed]
- Cabiltes, I.; Coghill, S.; Bowe, S.J.; Athan, E. Enterococcal bacteraemia “Silent but deadly”: A population-based cohort study. Intern. Med. J. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kristich, C.J.; Rice, L.B.; Arias, C.A. Enterococcal Infection-Treatment and Antibiotic Resistance. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Perl, T.M. The threat of vancomycin resistance. Am. J. Med. 1999, 106, 26S–37S, discussion 48S–52S. [Google Scholar] [CrossRef]
- Ceci, M.; Delpech, G.; Sparo, M.; Mezzina, V.; Sanchez Bruni, S.; Baldaccini, B. Clinical and microbiological features of bacteremia caused by Enterococcus faecalis. J. Infect. Dev. Ctries. 2015, 9, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Brussow, H.; Canchaya, C.; Hardt, W.D. Phages and the evolution of bacterial pathogens: From genomic rearrangements to lysogenic conversion. Microbiol. Mol. Biol. Rev. 2004, 68, 560–602. [Google Scholar] [CrossRef] [PubMed]
- Cisek, A.A.; Dabrowska, I.; Gregorczyk, K.P.; Wyzewski, Z. Phage Therapy in Bacterial Infections Treatment: One Hundred Years After the Discovery of Bacteriophages. Curr. Microbiol. 2017, 74, 277–283. [Google Scholar] [CrossRef]
- Abedon, S.T. Ecology of anti-biofilm agents II: Bacteriophage exploitation and biocontrol of biofilm bacteria. Pharmaceuticals (Basel) 2015, 8, 559–589. [Google Scholar] [CrossRef]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage therapy: An alternative to antibiotics in the age of multi-drug resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162–173. [Google Scholar] [CrossRef]
- Khalifa, L.; Gelman, D.; Shlezinger, M.; Dessal, A.L.; Coppenhagen-Glazer, S.; Beyth, N.; Hazan, R. Defeating antibiotic-and phage-resistant Enterococcus faecalis using a phage cocktail in Vitro and in a clot model. Front. Microbiol. 2018, 9, 326. [Google Scholar] [CrossRef]
- Paisano, A.F.; Spira, B.; Cai, S.; Bombana, A.C. In vitro antimicrobial effect of bacteriophages on human dentin infected with Enterococcus faecalis ATCC 29212. Oral Microbiol. Immunol. 2004, 19, 327–330. [Google Scholar] [CrossRef]
- Shlezinger, M.; Friedman, M.; Houri-Haddad, Y.; Hazan, R.; Beyth, N. Phages in a thermoreversible sustained-release formulation targeting E. Faecalis in vitro and in vivo. PLoS ONE 2019, 14, e0219599. [Google Scholar] [CrossRef] [PubMed]
- Szafranski, S.P.; Winkel, A.; Stiesch, M. The use of bacteriophages to biocontrol oral biofilms. J. Biotechnol. 2017, 250, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorski, A.; Miedzybrodzki, R.; Jonczyk-Matysiak, E.; Borysowski, J.; Letkiewicz, S.; Weber-Dabrowska, B. The fall and rise of phage therapy in modern medicine. Expert Opin. Biol. Ther. 2019, 19, 1115–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifa, L.; Coppenhagen-Glazer, S.; Shlezinger, M.; Kott-Gutkowski, M.; Adini, O.; Beyth, N.; Hazan, R. Complete Genome Sequence of Enterococcus Bacteriophage EFLK1. Genome Announc. 2015, 3, 6. [Google Scholar] [CrossRef] [PubMed]
- Howard-Varona, C.; Hargreaves, K.R.; Abedon, S.T.; Sullivan, M.B. Lysogeny in nature: Mechanisms, impact and ecology of temperate phages. ISME J. 2017, 11, 1511–1520. [Google Scholar] [CrossRef]
- Accetto, T.; Janez, N. The lytic Myoviridae of Enterobacteriaceae form tight recombining assemblages separated by discontinuities in genome average nucleotide identity and lateral gene flow. Microb. Genom. 2018, 4, e000169. [Google Scholar] [CrossRef]
- Nir-Paz, R.; Gelman, D.; Khouri, A.; Sisson, B.M.; Fackler, J.; Alkalay-Oren, S.; Khalifa, L.; Rimon, A.; Yerushalmy, O.; Bader, R.; et al. Successful treatment of antibiotic resistant poly-microbial bone infection with bacteriophages and antibiotics combination. Clin. Infect. Dis. 2019. [Google Scholar] [CrossRef]
- Gill, J.J.; Hyman, P. Phage choice, isolation, and preparation for phage therapy. Curr. Pharm. Biotechnol. 2010, 11, 2–14. [Google Scholar] [CrossRef]
- Torres-Barcelo, C.; Hochberg, M.E. Evolutionary Rationale for Phages as Complements of Antibiotics. Trends Microbiol. 2016, 24, 249–256. [Google Scholar] [CrossRef]
- Crain, C.M.; Kroeker, K.; Halpern, B.S. Interactive and cumulative effects of multiple human stressors in marine systems. Ecol. Lett. 2008, 11, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Gelman, D.; Beyth, S.; Lerer, V.; Adler, K.; Poradosu-Cohen, R.; Coppenhagen-Glazer, S.; Hazan, R. Combined bacteriophages and antibiotics as an efficient therapy against VRE Enterococcus faecalis in a mouse model. Res. Microbiol. 2018, 169, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Comeau, A.M.; Tetart, F.; Trojet, S.N.; Prere, M.F.; Krisch, H.M. Phage-Antibiotic Synergy (PAS): Beta-lactam and quinolone antibiotics stimulate virulent phage growth. PLoS ONE 2007, 2, e799. [Google Scholar] [CrossRef] [PubMed]
- Ryan, E.M.; Alkawareek, M.Y.; Donnelly, R.F.; Gilmore, B.F. Synergistic phage-antibiotic combinations for the control of Escherichia coli biofilms in vitro. FEMS Immunol. Med. Microbiol. 2012, 65, 395–398. [Google Scholar] [CrossRef]
- Bedi, M.S.; Verma, V.; Chhibber, S. Amoxicillin and specific bacteriophage can be used together for eradication of biofilm of Klebsiella pneumoniae B5055. World J. Microbiol. Biotechnol. 2009, 25, 1145–1151. [Google Scholar] [CrossRef]
- Kamal, F.; Dennis, J.J. Burkholderia cepacia complex Phage-Antibiotic Synergy (PAS): Antibiotics stimulate lytic phage activity. Appl. Environ. Microbiol. 2015, 81, 1132–1138. [Google Scholar] [CrossRef] [PubMed]
- Himmelweit, F. Combined action of penicillin and bacteriophage on staphylococci. Lancet 1945, 246, 104–105. [Google Scholar] [CrossRef]
- Knezevic, P.; Curcin, S.; Aleksic, V.; Petrusic, M.; Vlaski, L. Phage-antibiotic synergism: A possible approach to combatting pseudomonas aeruginosa. Res. Microbiol. 2013, 164, 55–60. [Google Scholar] [CrossRef]
- Akturk, E.; Oliveira, H.; Santos, S.B.; Costa, S.; Kuyumcu, S.; Melo, L.D.R.; Azeredo, J. Synergistic action of phage and antibiotics: Parameters to enhance the killing efficacy against mono and dual-species biofilms. Antibiotics 2019, 8, 103. [Google Scholar] [CrossRef]
- Oechslin, F.; Piccardi, P.; Mancini, S.; Gabard, J.; Moreillon, P.; Entenza, J.M.; Resch, G.; Que, Y.A. Synergistic interaction between phage therapy and antibiotics clears pseudomonas aeruginosa infection in endocarditis and reduces virulence. J. Infect. Dis. 2017, 215, 703–712. [Google Scholar] [CrossRef]
- Lin, Y.; Chang, R.Y.K.; Britton, W.J.; Morales, S.; Kutter, E.; Li, J.; Chan, H.K. Inhalable combination powder formulations of phage and ciprofloxacin for P. aeruginosa respiratory infections. Eur. J. Pharm. Biopharm. 2019, 142, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Sahm, D.F.; Kissinger, J.; Gilmore, M.S.; Murray, P.R.; Mulder, R.; Solliday, J.; Clarke, B. In vitro susceptibility studies of vancomycin-resistant Enterococcus faecalis. Antimicrob. Agents Chemother. 1989, 33, 1588–1591. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, I.T.; Banerjei, L.; Myers, G.S.; Nelson, K.E.; Seshadri, R.; Read, T.D.; Fouts, D.E.; Eisen, J.A.; Gill, S.R.; Heidelberg, J.F.; et al. Role of mobile DNA in the evolution of vancomycin-resistant Enterococcus faecalis. Science 2003, 299, 2071–2074. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, L.; Brosh, Y.; Gelman, D.; Coppenhagen-Glazer, S.; Beyth, S.; Poradosu-Cohen, R.; Que, Y.A.; Beyth, N.; Hazan, R. Targeting Enterococcus faecalis biofilms with phage therapy. Appl. Environ. Microbiol. 2015, 81, 2696–2705. [Google Scholar] [CrossRef] [PubMed]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Nir-Paz, R.; Eugster, M.R.; Zeiman, E.; Loessner, M.J.; Calendar, R. Listeria monocytogenes tyrosine phosphatases affect wall teichoic acid composition and phage resistance. FEMS Microbiol. Lett. 2012, 326, 151–160. [Google Scholar] [CrossRef]
- Shlezinger, M.; Khalifa, L.; Houri-Haddad, Y.; Coppenhagen-Glazer, S.; Resch, G.; Que, Y.A.; Beyth, S.; Dorfman, E.; Hazan, R.; Beyth, N. Phage Therapy: A new horizon in the antibacterial treatment of oral pathogens. Curr. Top. Med. Chem. 2017, 17, 1199–1211. [Google Scholar] [CrossRef]
- Sheviakova, O.I.; Sherman, R.Z.; Tatarinova, S.D. Treatment of dysentery in children with polymyxin and bacteriophage. Antibiotiki 1964, 9, 269–272. [Google Scholar]
- Sheviakova, O.I. Effect of bacteriophage on resistance of flexner’s bacillus to antibiotics. Antibiotiki (Mosc) 1956, 1, 40–42. [Google Scholar]
- Sheviakova, O.I.; IuS, K. Effect of combined antibiotics of the tetracycline series with bacteriophage on Shigella dysenteriae. Antibiotiki (Mosc) 1958, 3, 96–98. [Google Scholar]
- Chanishvili, N.; Chanishvili, T.; Tediashvili, M.; Barrow, P.A. Phages and their application against drug-resistant bacteria. J. Chem. Technol. Biotechnol. 2001, 76, 689–699. [Google Scholar] [CrossRef]
- Wegrzyn, G.; Licznerska, K.; Wegrzyn, A. Phage lambda—New insights into regulatory circuits. Adv. Virus Res. 2012, 82, 155–178. [Google Scholar] [PubMed]
- Huff, W.E.; Huff, G.R.; Rath, N.C.; Balog, J.M.; Donoghue, A.M. Therapeutic efficacy of bacteriophage and Baytril (enrofloxacin) individually and in combination to treat colibacillosis in broilers. Poult. Sci. 2004, 83, 1944–1947. [Google Scholar] [CrossRef] [PubMed]
- Hagens, S.; Habel, A.; Blasi, U. Augmentation of the antimicrobial efficacy of antibiotics by filamentous phage. Microb. Drug Resist. 2006, 12, 164–168. [Google Scholar] [CrossRef]
- Cheng, M.; Liang, J.; Zhang, Y.; Hu, L.; Gong, P.; Cai, R.; Zhang, L.; Zhang, H.; Ge, J.; Ji, Y.; et al. The bacteriophage ef-p29 efficiently protects against lethal vancomycin-resistant Enterococcus faecalis and alleviates gut microbiota imbalance in a murine bacteremia model. Front. Microbiol. 2017, 8, 837. [Google Scholar] [CrossRef]
- Duerkop, B.A.; Huo, W.; Bhardwaj, P.; Palmer, K.L.; Hooper, L.V. Molecular basis for lytic bacteriophage resistance in enterococci. MBio 2016, 7, 4–1304. [Google Scholar] [CrossRef]
- Henein, A. What are the limitations on the wider therapeutic use of phage? Bacteriophage 2013, 3, e24872. [Google Scholar] [CrossRef] [Green Version]
- Busque, L.; Porwit, A.; Day, R.; Olney, H.J.; Leber, B.; Ethier, V.; Sirhan, S.; Foltz, L.; Prchal, J.; Kamel-Reid, S.; et al. Laboratory investigation of Myeloproliferative Neoplasms (MPNs): Recommendations of the Canadian Mpn Group. Am. J. Clin. Pathol. 2016, 146, 408–422. [Google Scholar] [CrossRef]
- Reynolds, P.E. Structure, biochemistry and mechanism of action of glycopeptide antibiotics. Eur. J. Clin. Microbiol. Infect. Dis. 1989, 8, 943–950. [Google Scholar] [CrossRef]
- Gold, H.S. Vancomycin-resistant enterococci: Mechanisms and clinical observations. Clin. Infect. Dis. 2001, 33, 210–219. [Google Scholar] [CrossRef]
- Young, I.; Wang, I.; Roof, W.D. Phages will out: Strategies of host cell lysis. Trends Microbiol. 2000, 8, 120–128. [Google Scholar] [CrossRef]
- Latka, A.; Maciejewska, B.; Majkowska-Skrobek, G.; Briers, Y.; Drulis-Kawa, Z. Bacteriophage-encoded virion-associated enzymes to overcome the carbohydrate barriers during the infection process. Appl. Microbiol. Biotechnol. 2017, 101, 3103–3119. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Origin a | Antibiotic Resistance b | Phage EFLK1 c | Vancomycin MIC (mg/mL) d | Phage EFLK1 + Vancomycin MIC (mg/mL) e |
|---|---|---|---|---|---|
| Enterococcus faecalis V583 | ATCC 700802 | Vancomycin Gentamicin | S | >0.25 | 0.015 |
| Enterococcus faecalis Aef01 | Clinically isolated from urine | - | S | 0.003 | <0.001 |
| Enterococcus faecalis Aef03 | Clinically isolated from urine | - | S | 0.007 | <0.001 |
| Enterococcus faecalis Aef05 | Clinically isolated from venal blood flow | Erythromycin Gentamicin | S | 0.003 | <0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shlezinger, M.; Coppenhagen-Glazer, S.; Gelman, D.; Beyth, N.; Hazan, R. Eradication of Vancomycin-Resistant Enterococci by Combining Phage and Vancomycin. Viruses 2019, 11, 954. https://doi.org/10.3390/v11100954
Shlezinger M, Coppenhagen-Glazer S, Gelman D, Beyth N, Hazan R. Eradication of Vancomycin-Resistant Enterococci by Combining Phage and Vancomycin. Viruses. 2019; 11(10):954. https://doi.org/10.3390/v11100954
Chicago/Turabian StyleShlezinger, Mor, Shunit Coppenhagen-Glazer, Daniel Gelman, Nurit Beyth, and Ronen Hazan. 2019. "Eradication of Vancomycin-Resistant Enterococci by Combining Phage and Vancomycin" Viruses 11, no. 10: 954. https://doi.org/10.3390/v11100954