The Characteristics and Genome Analysis of vB_AviM_AVP, the First Phage Infecting Aerococcus viridans

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Isolation of the Aerococcus viridans Phage
2.3. CsCl Density Gradient Centrifugation
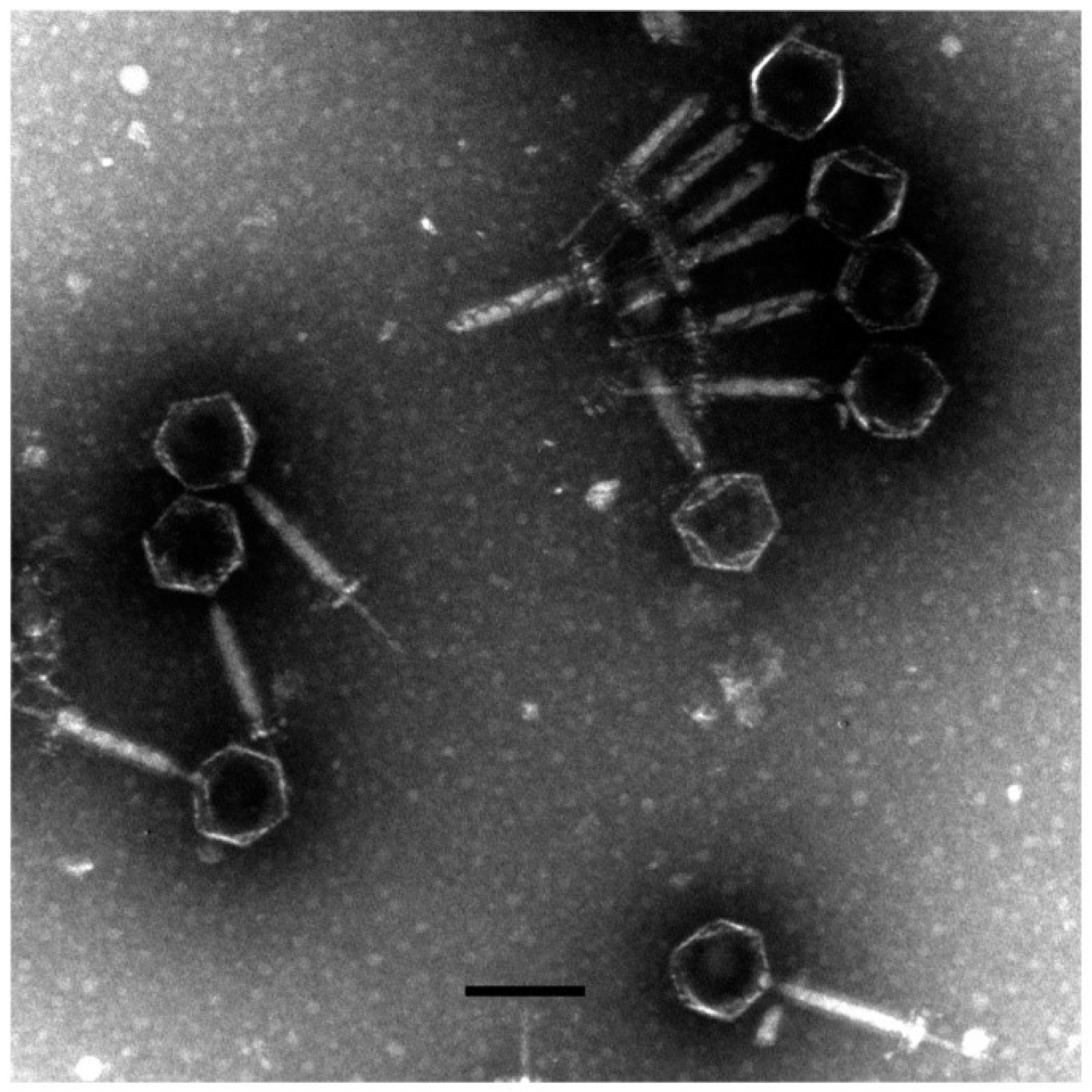
2.4. TEM of vB_AviM_AVP
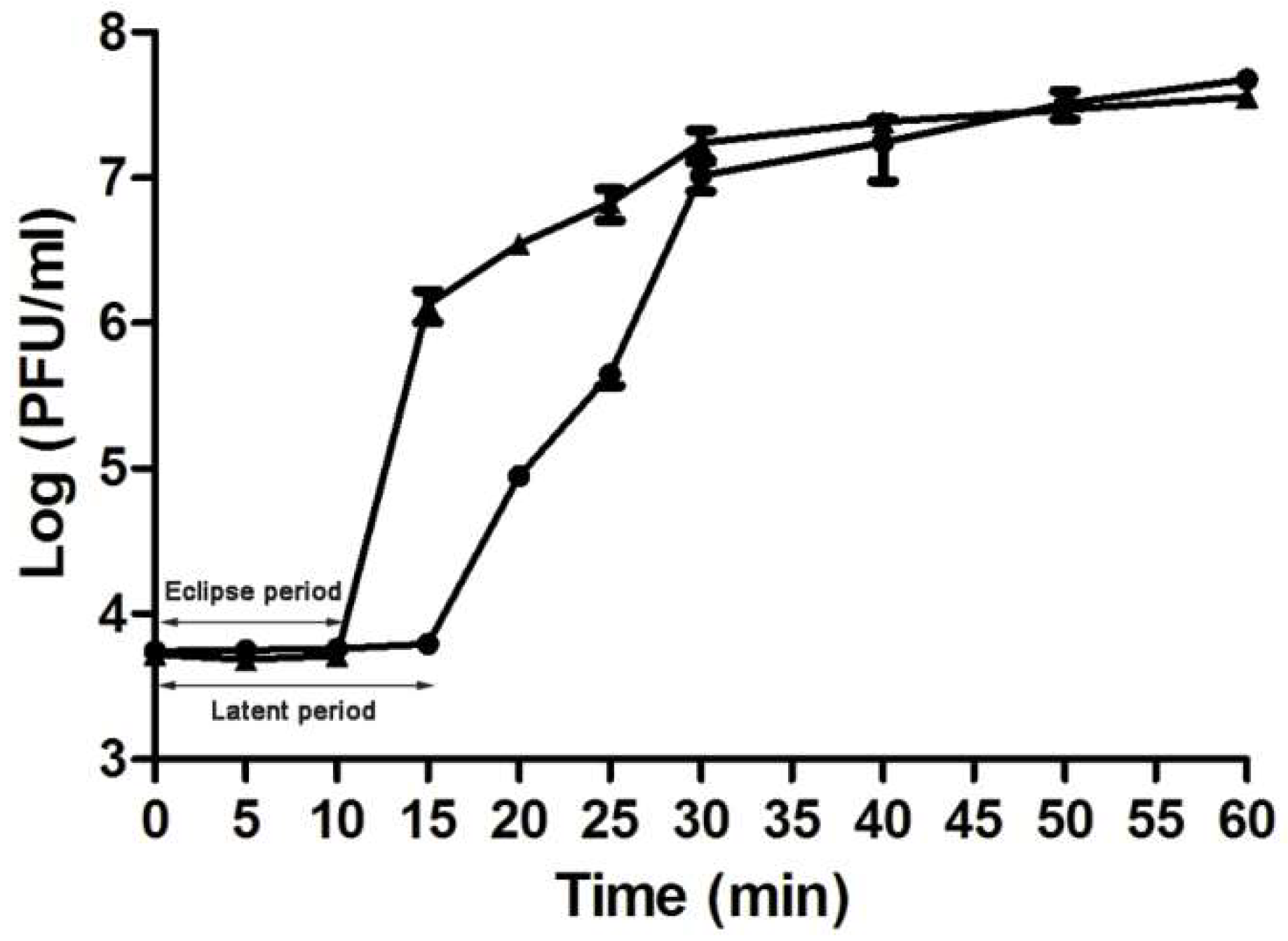
2.5. One-Step Growth Curve
2.6. Sequencing and Bioinformatics Analysis of the AVP Genome
3. Results and Discussion
3.1. The Characteristics of Aerococcus viridans
3.2. The Biological Characteristics of vB_AviM_AVP
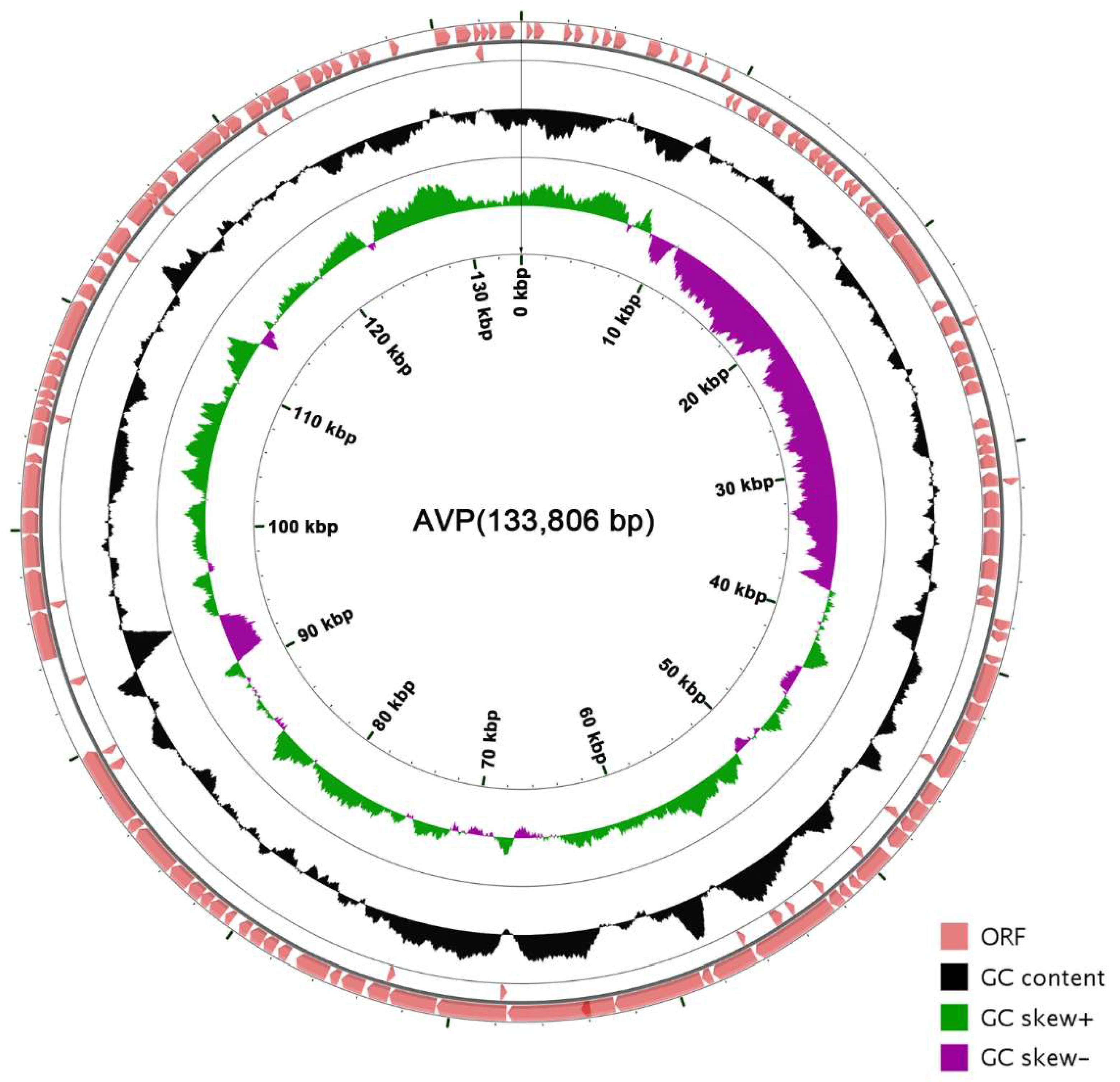
3.3. The General Characteristics of the AVP Genome
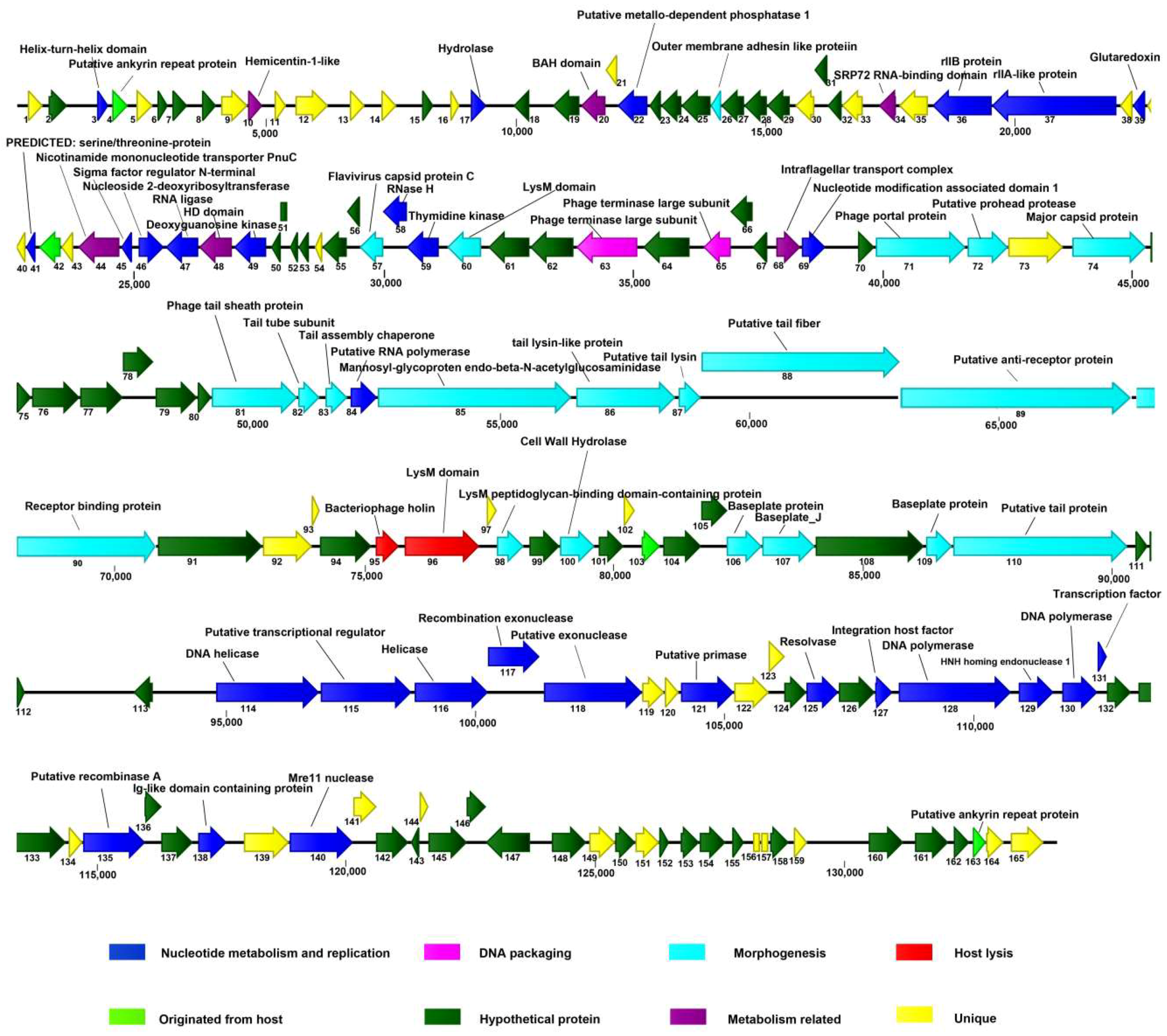
3.4. The Putative ORFs of AVP
3.5. The Lysis Module
3.6. The Nucleotide Metabolism and Replication Module
3.7. The Structure and Packaging Module
Nucleotide Sequence Accession Numbers
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sun, M.; Gao, J.; Ali, T.; Yu, D.; Zhang, S.; Khan, S.U.; Fanning, S.; Han, B. Characteristics of Aerococcus viridans isolated from bovine subclinical mastitis and its effect on milk SCC, yield, and composition. Trop. Anim. Health Prod. 2017, 49, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, M. Aerococci and aerococcal infections. J. Infect. 2013, 66, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Ma, Y.; Ma, J.; Dong, W.; Yao, H. Acute meningitis of piglets and mice caused by co-infected with Streptococcus suis and Aerococcus viridans. Microb. Pathog. 2017, 106, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Kerbaugh, M.A.; Evans, J.B. Aerococcus viridans in the hospital environment. Appl. Microbiol. 1968, 16, 519–523. [Google Scholar] [PubMed]
- Stebbing, P.D.; Pond, M.J.; Peeler, E.; Small, H.J.; Greenwood, S.J.; Verner-Jeffreys, D. Limited prevalence of gaffkaemia (Aerococcus viridans var. homari) isolated from wild-caught European lobsters Homarus gammarus in England and Wales. Dis. Aquat. Organ. 2012, 100, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Ke, X.; Lu, M.; Ye, X.; Gao, F.; Zhu, H.; Huang, Z. Recovery and pathogenicity analysis of Aerococcus viridans isolated from tilapia (Orecohromis niloticus) cultured in southwest of China. Aquaculture 2012, 342–343, 18–23. [Google Scholar] [CrossRef]
- Martin, V.; Vela, A.I.; Gilbert, M.; Cebolla, J.; Goyache, J.; Dominguez, L.; Fernandez-Garayzabal, J.F. Characterization of Aerococcus viridans isolates from swine clinical specimens. J. Clin. Microbiol. 2007, 45, 3053–3057. [Google Scholar] [CrossRef]
- Liu, G.; Liu, Y.; Ali, T.; Ferreri, M.; Gao, J.; Chen, W.; Yin, J.; Su, J.; Fanning, S.; Han, B. Molecular and Phenotypic Characterization of Aerococcus viridans Associated with Subclinical Bovine Mastitis. PLoS ONE 2015, 10, e0125001. [Google Scholar] [CrossRef]
- Saishu, N.; Morimoto, K.; Yamasato, H.; Ozaki, H.; Murase, T. Characterization of Aerococcus viridans isolated from milk samples from cows with mastitis and manure samples. J. Vet. Med. Sci. 2015, 77, 1037–1042. [Google Scholar] [CrossRef]
- Zadoks, R.N.; Gonzalez, R.N.; Boor, K.J.; Schukken, Y.H. Mastitis-causing streptococci are important contributors to bacterial counts in raw bulk tank milk. J. Food Prot. 2004, 67, 2644–2650. [Google Scholar] [CrossRef]
- Spakova, T.; Elecko, J.; Vasil, M.; Legath, J.; Pristas, P.; Javorsky, P. Limited genetic diversity of Aerococcus viridans strains isolated from clinical and subclinical cases of bovine mastitis in Slovakia. Pol. J. Vet. Sci. 2012, 15, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Gopalachar, A.; Akins, R.L.; Davis, W.R.; Siddiqui, A.A. Urinary tract infection caused by Aerococcus viridans, a case report. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2004, 10, CS73–CS75. [Google Scholar]
- Uh, Y.; Son, J.S.; Jang, I.H.; Yoon, K.J.; Hong, S.K. Penicillin-resistant Aerococcus viridans bacteremia associated with granulocytopenia. J. Korean Med. Sci. 2002, 17, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Nathavitharana, K.A.; Arseculeratne, S.N.; Aponso, H.A.; Vijeratnam, R.; Jayasena, L.; Navaratnam, C. Acute meningitis in early childhood caused by Aerococcus viridans. Br. Med. J. 1983, 286, 1248. [Google Scholar] [CrossRef]
- Facklam, R.; Elliott, J.A. Identification, classification, and clinical relevance of catalase-negative, gram-positive cocci, excluding the streptococci and enterococci. Clin. Microbiol. Rev. 1995, 8, 479–495. [Google Scholar] [CrossRef]
- Popescu, G.A.; Benea, E.; Mitache, E.; Piper, C.; Horstkotte, D. An unusual bacterium, Aerococcus viridans, and four cases of infective endocarditis. J. Heart Valve Dis. 2005, 14, 317–319. [Google Scholar] [PubMed]
- Buu-Hoi, A.; Le Bouguenec, C.; Horaud, T. Genetic basis of antibiotic resistance in Aerococcus viridans. Antimicrob. Agents Chemother. 1989, 33, 529–534. [Google Scholar] [CrossRef]
- Zhou, W.Q.; Niu, D.M.; Zhang, Z.Z.; Ning, M.Z.; Shen, H.; Zhang, K. Vancomycin resistance due to VanA in an Aerococcus viridans isolate. Indian J. Med. Microbiol. 2014, 32, 462–465. [Google Scholar]
- Hatfull, G.F.; Hendrix, R.W. Bacteriophages and their genomes. Curr. Opin. Virol. 2011, 1, 298–303. [Google Scholar] [CrossRef] [Green Version]
- Karpe, Y.A.; Kanade, G.D.; Pingale, K.D.; Arankalle, V.A.; Banerjee, K. Genomic characterization of Salmonella bacteriophages isolated from India. Virus Genes 2016, 52, 117–126. [Google Scholar] [CrossRef]
- Gu, J.; Liu, X.; Li, Y.; Han, W.; Lei, L.; Yang, Y.; Zhao, H.; Gao, Y.; Song, J.; Lu, R.; et al. A method for generation phage cocktail with great therapeutic potential. PLoS ONE 2012, 7, e31698. [Google Scholar] [CrossRef] [PubMed]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G., Jr. Bacteriophage therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Kropinski, A.M.; Waddell, T.; Meng, J.; Franklin, K.; Ackermann, H.W.; Ahmed, R.; Mazzocco, A.; Yates, J., 3rd; Lingohr, E.J.; Johnson, R.P. The host-range, genomics and proteomics of Escherichia coli O157:H7 bacteriophage rV5. Virol. J. 2013, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Wang, S.; Yan, G.; Sun, C.; Feng, X.; Gu, J.; Han, W.; Lei, L. Genome sequencing and analysis of an Escherichia coli phage vB_EcoM-ep3 with a novel lysin, Lysep3. Virus Genes 2015, 50, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Habann, M.; Leiman, P.G.; Vandersteegen, K.; Van den Bossche, A.; Lavigne, R.; Shneider, M.M.; Bielmann, R.; Eugster, M.R.; Loessner, M.J.; Klumpp, J. Listeria phage A511, a model for the contractile tail machineries of SPO1-related bacteriophages. Mol. Microbiol. 2014, 92, 84–99. [Google Scholar] [CrossRef] [PubMed]
- Denes, T.; Vongkamjan, K.; Ackermann, H.W.; Moreno Switt, A.I.; Wiedmann, M.; den Bakker, H.C. Comparative genomic and morphological analyses of Listeria phages isolated from farm environments. Appl. Environ. Microbiol. 2014, 80, 4616–4625. [Google Scholar] [CrossRef] [PubMed]
- Kwiatek, M.; Parasion, S.; Mizak, L.; Gryko, R.; Bartoszcze, M.; Kocik, J. Characterization of a bacteriophage, isolated from a cow with mastitis, that is lytic against Staphylococcus aureus strains. Arch. Virol. 2012, 157, 225–234. [Google Scholar] [CrossRef]
- Jia, H.; Dong, W.; Yuan, L.; Ma, J.; Bai, Q.; Pan, Z.; Lu, C.; Yao, H. Characterization and complete genome sequence analysis of Staphylococcus aureus bacteriophage JS01. Virus Genes 2015, 50, 345–348. [Google Scholar] [CrossRef]
- Bai, Q.; Zhang, W.; Yang, Y.; Tang, F.; Nguyen, X.; Liu, G.; Lu, C. Characterization and genome sequencing of a novel bacteriophage infecting Streptococcus agalactiae with high similarity to a phage from Streptococcus pyogenes. Arch. Virol. 2013, 158, 1733–1741. [Google Scholar] [CrossRef]
- Dalmasso, M.; de Haas, E.; Neve, H.; Strain, R.; Cousin, F.J.; Stockdale, S.R.; Ross, R.P.; Hill, C. Isolation of a Novel Phage with Activity against Streptococcus mutans Biofilms. PLoS ONE 2015, 10, e0138651. [Google Scholar] [CrossRef]
- Grant, K.A.; Dickinson, J.H.; Collins, M.D.; Kroll, R.G. Rapid identification of Aerococcus viridans using the polymerase chain reaction and an oligonucleotide probe. FEMS Microbiol. Lett. 1992, 74, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Xu, W.; Lei, L.; Huang, J.; Feng, X.; Sun, C.; Du, C.; Zuo, J.; Li, Y.; Du, T.; et al. LysGH15, a novel bacteriophage lysin, protects a murine bacteremia model efficiently against lethal methicillin-resistant Staphylococcus aureus infection. J. Clin. Microbiol. 2011, 49, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Saralamba, N.; Mayxay, M.; Newton, P.N.; Smithuis, F.; Nosten, F.; Archasuksan, L.; Pukrittayakamee, S.; White, N.J.; Day, N.P.J.; Dondorp, A.M.; et al. Genetic polymorphisms in the circumsporozoite protein of Plasmodium malariae show a geographical bias. Malar. J. 2018, 17, 269. [Google Scholar] [CrossRef] [PubMed]
- Gadagkar, R.; Gopinathan, K.P. Bacteriophage burst size during multiple infections. J. Biosci. 1980, 2, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Search and Contextual Analysis of Transfer RNA Genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Carkaci, D.; Dargis, R.; Nielsen, X.C.; Skovgaard, O.; Fuursted, K.; Christensen, J.J. Complete Genome Sequences of Aerococcus christensenii CCUG 28831T, Aerococcus sanguinicola CCUG 43001T, Aerococcus urinae CCUG 36881T, Aerococcus urinaeequi CCUG 28094T, Aerococcus urinaehominis CCUG 42038 BT, and Aerococcus viridans CCUG 4311T. Genome Announc. 2016, 4, e00302-16. [Google Scholar] [CrossRef]
- Stothard, P.; Wishart, D.S. Circular genome visualization and exploration using CGView. Bioinformatics 2005, 21, 537–539. [Google Scholar] [CrossRef]
- Visweswaran, G.R.; Leenhouts, K.; van Roosmalen, M.; Kok, J.; Buist, G. Exploiting the peptidoglycan-binding motif, LysM, for medical and industrial applications. Appl. Microbiol. Biotechnol. 2014, 98, 4331–4345. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Kong, J.; Kong, W.; Guo, T.; Ji, M. Characterization of a novel LysM domain from Lactobacillus fermentum bacteriophage endolysin and its use as an anchor to display heterologous proteins on the surfaces of lactic acid bacteria. Appl. Environ. Microbiol. 2010, 76, 2410–2418. [Google Scholar] [CrossRef] [PubMed]
- Guilliam, T.A.; Keen, B.A.; Brissett, N.C.; Doherty, A.J. Primase-polymerases are a functionally diverse superfamily of replication and repair enzymes. Nucleic Acids Res. 2015, 43, 6651–6664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanyal, S.J.; Yang, T.C.; Catalano, C.E. Integration host factor assembly at the cohesive end site of the bacteriophage lambda genome: Implications for viral DNA packaging and bacterial gene regulation. Biochemistry 2014, 53, 7459–7470. [Google Scholar] [CrossRef] [PubMed]
- Greenstein, D.; Zinder, N.D.; Horiuchi, K. Integration host factor interacts with the DNA replication enhancer of filamentous phage f1. Proc. Natl. Acad. Sci. USA 1988, 85, 6262–6266. [Google Scholar] [CrossRef]
- Saha, S.; Haggard-Ljungquist, E.; Nordstrom, K. Integration host factor is necessary for lysogenization of Escherichia coli by bacteriophage P2. Mol. Microbiol. 1990, 4, 3–11. [Google Scholar] [CrossRef]
- Bonocora, R.P.; Belfort, M. Mapping homing endonuclease cleavage sites using in vitro generated protein. Methods Mol. Biol. 2014, 1123, 55–67. [Google Scholar]
- Kwan, T.; Liu, J.; DuBow, M.; Gros, P.; Pelletier, J. The complete genomes and proteomes of 27 Staphylococcus aureus bacteriophages. Proc. Natl. Acad. Sci. USA 2005, 102, 5174–5179. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Liu, X.; Yang, M.; Li, Y.; Sun, C.; Lu, R.; Song, J.; Zhang, Q.; Lei, L.; Feng, X.; et al. Genomic characterization of lytic Staphylococcus aureus phage GH15: Providing new clues to intron shift in phages. J. Gen. Virol. 2013, 94 Pt 4, 906–915. [Google Scholar] [CrossRef]
- Foley, S.; Bruttin, A.; Brussow, H. Widespread distribution of a group I intron and its three deletion derivatives in the lysin gene of Streptococcus thermophilus bacteriophages. J. Virol. 2000, 74, 611–618. [Google Scholar] [CrossRef]
- O’Flaherty, S.; Coffey, A.; Edwards, R.; Meaney, W.; Fitzgerald, G.F.; Ross, R.P. Genome of staphylococcal phage K: A new lineage of Myoviridae infecting gram-positive bacteria with a low G+C content. J. Bacteriol. 2004, 186, 2862–2871. [Google Scholar] [CrossRef] [PubMed]
- Buis, J.; Wu, Y.; Deng, Y.; Leddon, J.; Westfield, G.; Eckersdorff, M.; Sekiguchi, J.M.; Chang, S.; Ferguson, D.O. Mre11 nuclease activity has essential roles in DNA repair and genomic stability distinct from ATM activation. Cell 2008, 135, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, L.; Coppenhagen-Glazer, S.; Shlezinger, M.; Kott-Gutkowski, M.; Adini, O.; Beyth, N.; Hazan, R. Complete Genome Sequence of Enterococcus Bacteriophage EFLK1. Genome Announc. 2015, 3, e01308-15. [Google Scholar] [CrossRef] [PubMed]
- Casjens, S.; Hendrix, R. Control Mechanisms in dsDNA Bacteriophage Assembly. In The Bacteriophages; Springer: Basel, Switzerland, 1988; Chapter 2; pp. 15–91. [Google Scholar]
- Gutierrez, D.; Vandenheuvel, D.; Martinez, B.; Rodriguez, A.; Lavigne, R.; Garcia, P. Two Phages, phiIPLA-RODI and phiIPLA-C1C, Lyse Mono- and Dual-Species Staphylococcal Biofilms. Appl. Environ. Microbiol. 2015, 81, 3336–3348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Droge, A.; Tavares, P. In vitro packaging of DNA of the Bacillus subtilis bacteriophage SPP1. J. Mol. Biol. 2000, 296, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Fokine, A.; Rossmann, M.G. Common Evolutionary Origin of Procapsid Proteases, Phage Tail Tubes, and Tubes of Bacterial Type VI Secretion Systems. Structure 2016, 24, 1928–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casjens, S.R.; Gilcrease, E.B.; Winn-Stapley, D.A.; Schicklmaier, P.; Schmieger, H.; Pedulla, M.L.; Ford, M.E.; Houtz, J.M.; Hatfull, G.F.; Hendrix, R.W. The generalized transducing Salmonella bacteriophage ES18: Complete genome sequence and DNA packaging strategy. J. Bacteriol. 2005, 187, 1091–1104. [Google Scholar] [CrossRef]
- Rossmann, M.G. Structure of viruses: A short history. Q. Rev. Biophys. 2013, 46, 133–180. [Google Scholar] [CrossRef]
- Uchiyama, J.; Rashel, M.; Takemura, I.; Wakiguchi, H.; Matsuzaki, S. In silico and in vivo evaluation of bacteriophage phiEF24C, a candidate for treatment of Enterococcus faecalis infections. Appl. Environ. Microbiol. 2008, 74, 4149–4163. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xi, H.; Dai, J.; Tong, Y.; Cheng, M.; Zhao, F.; Fan, H.; Li, X.; Cai, R.; Ji, Y.; Sun, C.; et al. The Characteristics and Genome Analysis of vB_AviM_AVP, the First Phage Infecting Aerococcus viridans. Viruses 2019, 11, 104. https://doi.org/10.3390/v11020104
Xi H, Dai J, Tong Y, Cheng M, Zhao F, Fan H, Li X, Cai R, Ji Y, Sun C, et al. The Characteristics and Genome Analysis of vB_AviM_AVP, the First Phage Infecting Aerococcus viridans. Viruses. 2019; 11(2):104. https://doi.org/10.3390/v11020104
Chicago/Turabian StyleXi, Hengyu, Jiaxin Dai, Yigang Tong, Mengjun Cheng, Feiyang Zhao, Hang Fan, Xinwei Li, Ruopeng Cai, Yalu Ji, Changjiang Sun, and et al. 2019. "The Characteristics and Genome Analysis of vB_AviM_AVP, the First Phage Infecting Aerococcus viridans" Viruses 11, no. 2: 104. https://doi.org/10.3390/v11020104
APA StyleXi, H., Dai, J., Tong, Y., Cheng, M., Zhao, F., Fan, H., Li, X., Cai, R., Ji, Y., Sun, C., Feng, X., Lei, L., Rahman, S. u., Han, W., & Gu, J. (2019). The Characteristics and Genome Analysis of vB_AviM_AVP, the First Phage Infecting Aerococcus viridans. Viruses, 11(2), 104. https://doi.org/10.3390/v11020104