New Method for Differentiation of Granuloviruses (Betabaculoviruses) Based on Real-Time Polymerase Chain Reaction (Real-Time PCR)

 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
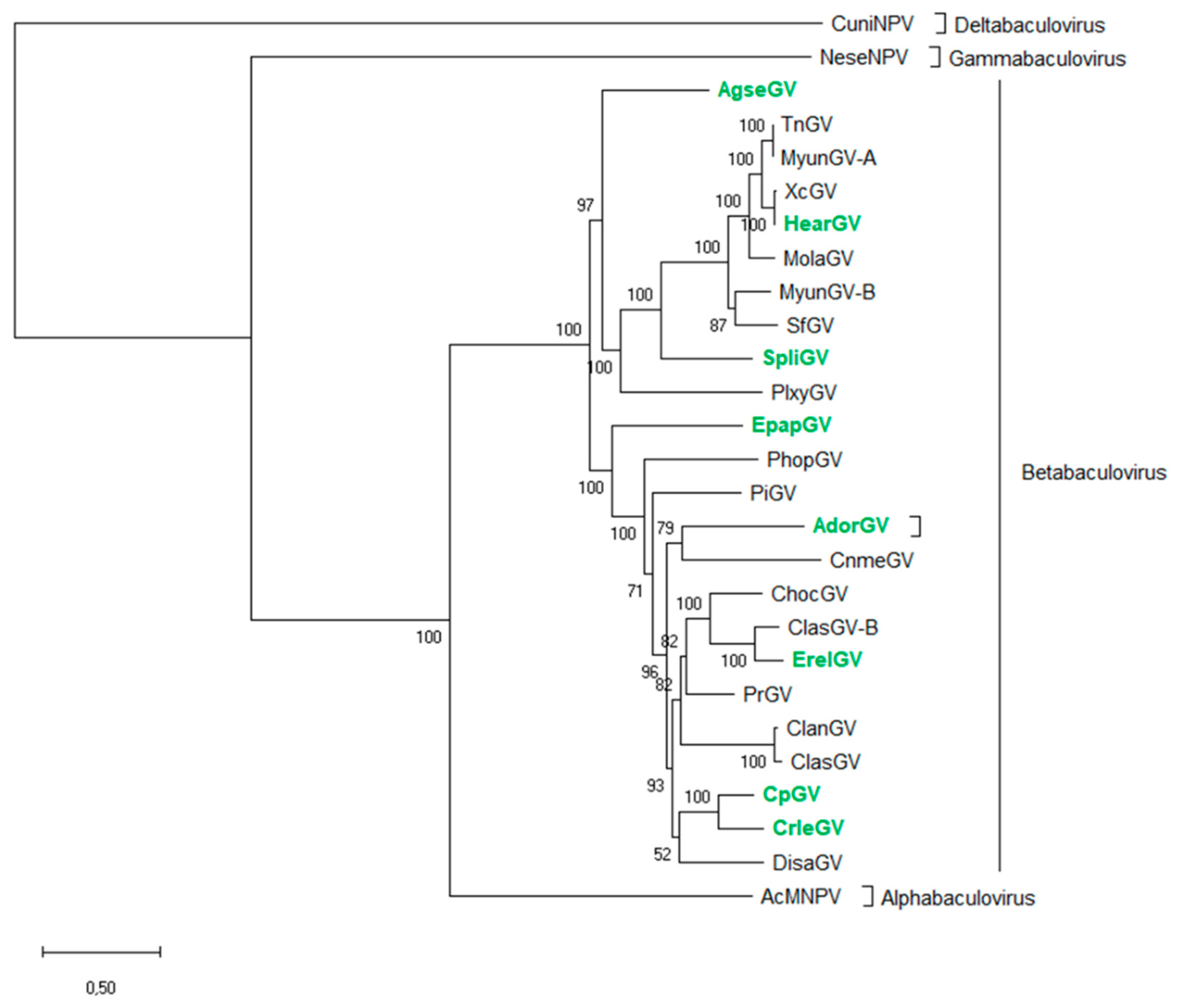
2.1. Phylogenetic Analysis of Granuloviruses
2.2. Determination of Betabaculovirus Representative Group
2.3. Virus Purification and DNA Isolation
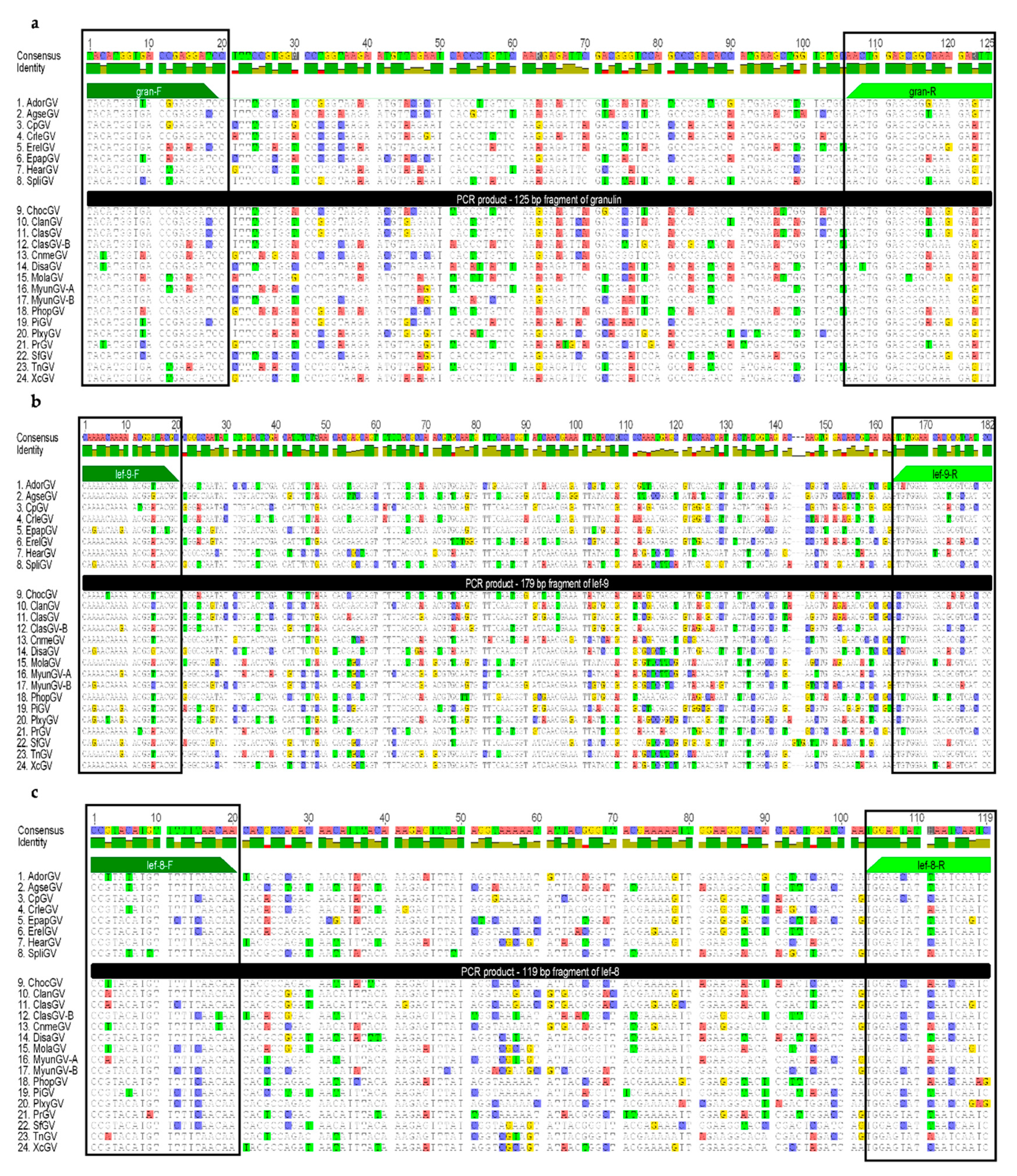
2.4. Granulin, Late Expression Factor-9 and Late Expression Factor-8 Nucleotide Sequence Alignments and Degenerate Primer Design; Amino Acid Alignment of the PCR Products.
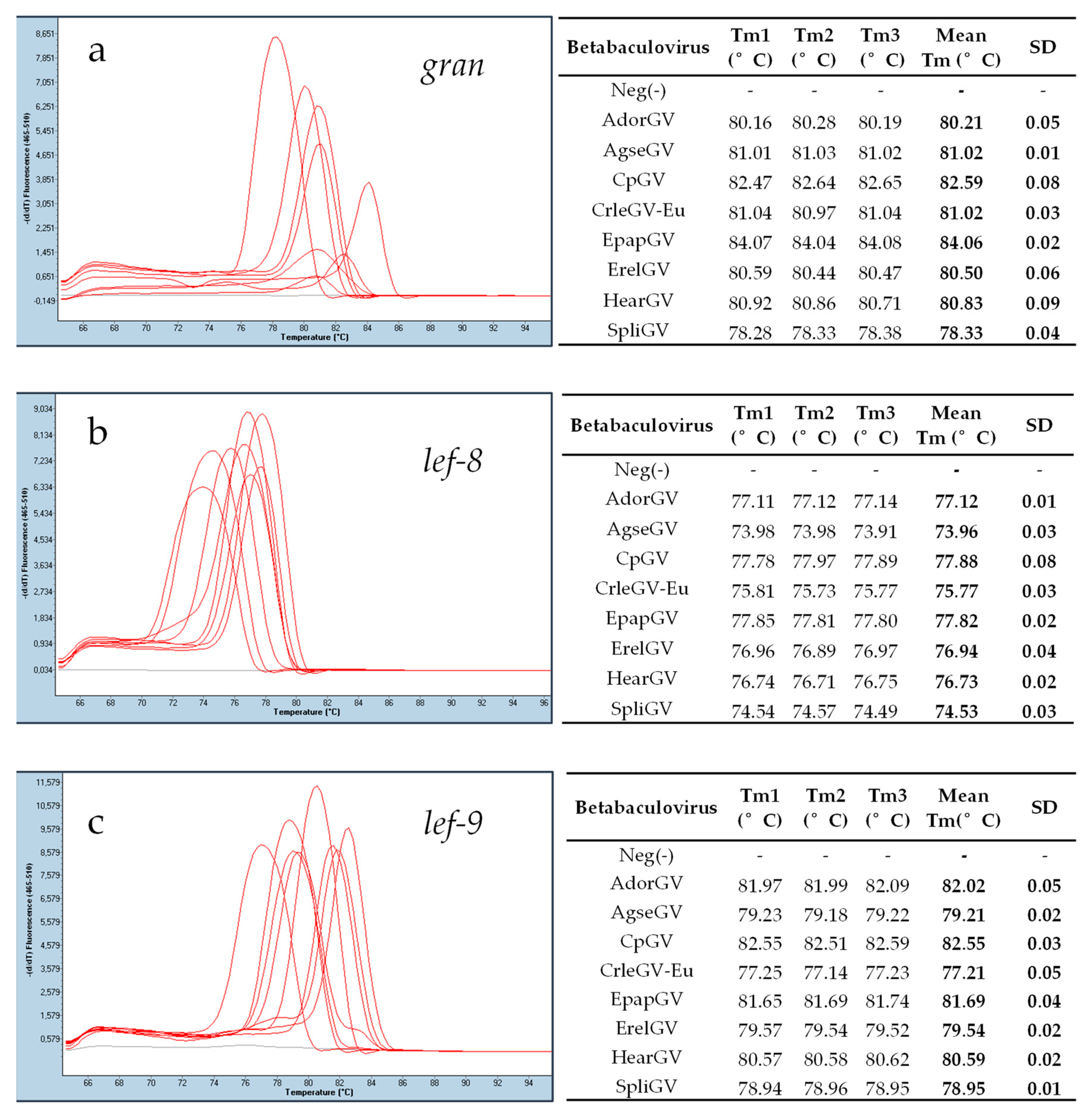
2.5. Real-Time PCR Reaction
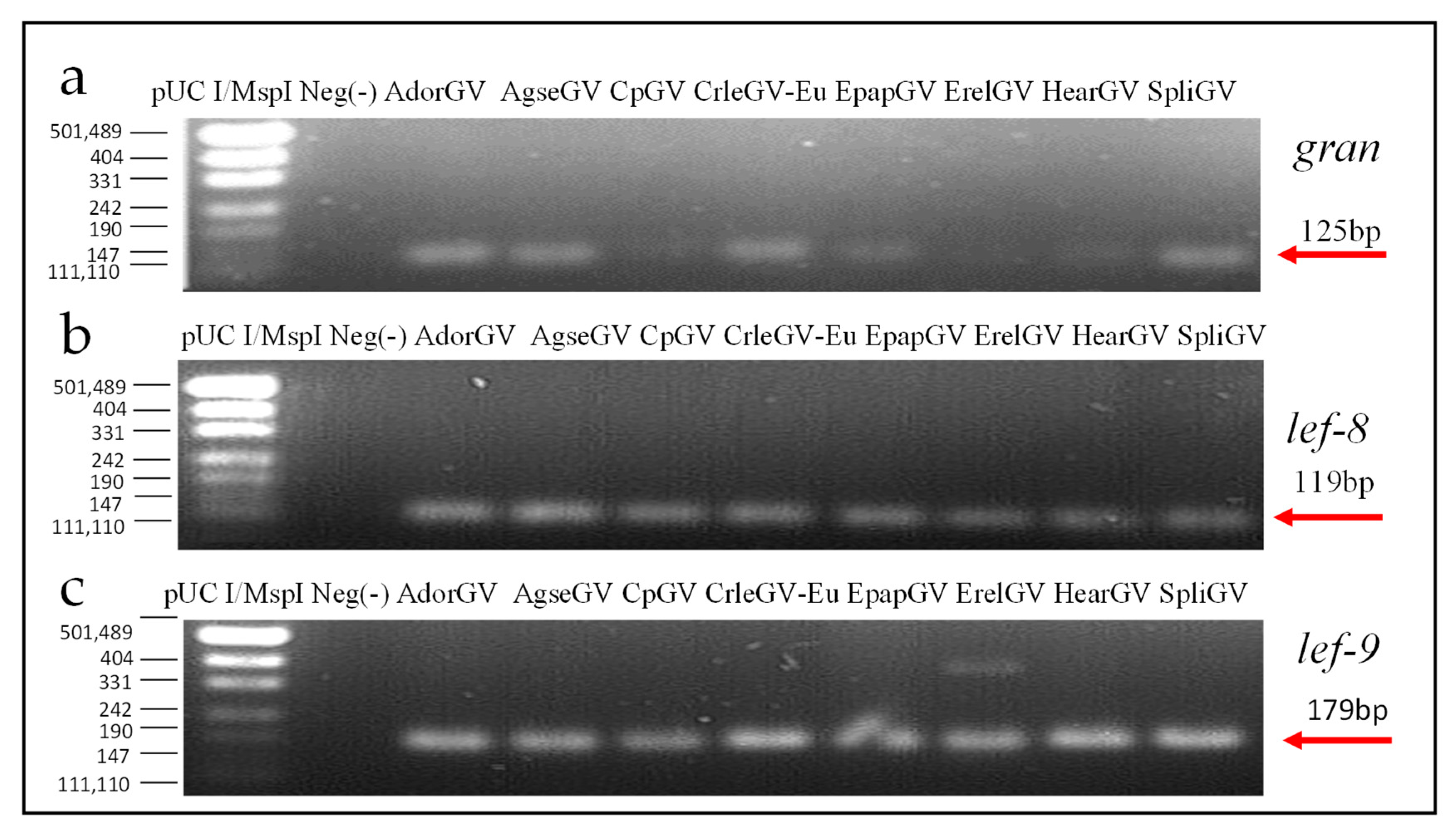
2.6. Agarose Gel Electrophoresis
3. Results
3.1. Phylogenetic Analysis and the Representative group of Betabaculovirus
3.2. Granulin, Late Expression Factor-9, and Late Expression Factor-8 Nucleotide and Amino Acid Sequence Alignment
3.3. Real-Time PCR Assay
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martignioni, M.; Iwai, P.J. A catalogue of viral diseases of insects, mites and thicks. In Microbial Control of Pests and Plant Diseases; Burges, H.D., Ed.; Arcade: London, UK, 1987; pp. 897–911. [Google Scholar]
- Szewczyk, B.; Souza, M.L.; Castro, M.E.B.; Moscardi, M.L.; Moscardi, F. Baculovirus Biopesticides. In Pesticides–Formulation, Effects, Fate; Stoytcheva, M., Ed.; Intech Open: London, UK, 2011; pp. 25–36. [Google Scholar]
- Haase, S.; Sciocco-Cap, A.; Romanowski, V. Baculovirus insecticides in Latin America: Historical overview, current status and future perspectives. Viruses 2015, 7, 2230–2267. [Google Scholar] [CrossRef] [PubMed]
- Blissard, G.W.; Rohrmann, G.F. Baculovirus Diversity and Molecular Biology. Annu. Rev. Entomol. 1990, 35, 127–155. [Google Scholar] [CrossRef]
- Lauzon, H.A.M.; Lucarotti, C.J.; Krell, P.J.; Feng, Q.; Retnakaran, A.; Arif, B.M. Sequence and organization of the Neodiprion lecontei nucleopolyhedrovirus genome. J. Virol. 2004, 78, 7023–7035. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, T.; Ko, R.; Okano, K.; Seong, S.I.; Goto, C.; Maeda, S. Sequence analysis of the Xestia c-nigrum granulovirus genome. Virology 1999, 262, 277–297. [Google Scholar] [CrossRef] [PubMed]
- Jehle, J.A.; Blissard, G.W.; Bonning, B.C.; Cory, J.S.; Herniou, E.A.; Rohrmann, G.F.; Theilmann, D.A.; Thiem, S.M.; Vlak, J.M. On the classification and nomenclature of baculoviruses: A proposal for revision. Arch. Virol. 2006, 151, 1257–1266. [Google Scholar] [CrossRef] [Green Version]
- Herniou, E.A.; Jehle, J.A. Baculovirus phylogeny and evolution. Curr. Drug Targets 2007, 8, 1043–1050. [Google Scholar] [CrossRef]
- Rohrmann, G.F. Polyhedrin structure. J. Gen. Virol. 1986, 67, 1499–1513. [Google Scholar] [CrossRef]
- Rohrmann, G.F. Baculovirus Molecular Biology, 3rd ed.; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2013. [Google Scholar]
- Federici, B.A. Baculovirus pathogenesis. In The Baculoviruses; Miller, L.K., Ed.; Springer Science & Business Media: New York, NY, USA, 1997; pp. 33–59. [Google Scholar]
- Harrison, R.L.; Popham, H.J.R. Genomic sequence analysis of a granulovirus isolated from the Old World bollworm, Helicoverpa armigera. Virus Genes 2008, 36, 565–581. [Google Scholar] [CrossRef]
- Luque, T.; Finch, R.; Crook, N.; O’Reilly, D.R.; Winstanley, D. The complete sequence of the Cydia pomonella granulovirus genome. J. Gen. Virol. 2001, 82, 2531–2547. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.; Kirkman, W.; Richards, G.; Stephen, P.; Moore, S.D.; Kirkman, W.; Richards, G.I.; Stephen, P.R. The Cryptophlebia Leucotreta Granulovirus—10 Years of Commercial Field Use. Viruses 2015, 7, 1284–1312. [Google Scholar] [CrossRef] [Green Version]
- Tanada, Y.; Kaya, H. Insect Pathology, 1st ed.; Academic Press: Cambridge, MA, USA, 1993; ISBN 978-0-08-092625-4. [Google Scholar]
- Ferrelli, M.L.; Salvador, R.; Biedma, M.E.; Berretta, M.F.; Haase, S.; Sciocco-Cap, A.; Ghiringhelli, P.D.; Romanowski, V. Genome of Epinotia aporema granulovirus (EpapGV), a polyorganotropic fast killing betabaculovirus with a novel thymidylate kinase gene. BMC Genom. 2012, 13, 548. [Google Scholar] [CrossRef]
- Federici, B.A.; Stern, V.M. Replication and occlusion of a granulosis virus in larval and adult midgut epithelium of the western grapeleaf skeletonizer, Harrisina brillians. J. Invertebr. Pathol. 1990, 56, 401–414. [Google Scholar] [CrossRef]
- Bideshi, D.K.; Bigot, Y.; Federici, B.A. Molecular characterization and phylogenetic analysis of the Harrisina brillians granulovirus granulin gene. Arch. Virol. 2000, 145, 1933–1945. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Zhu, Z.; Liu, X.; Hou, D.; Wang, J.; Zhang, L.; Wang, M.; Kou, Z.; Wang, H.; Deng, F.; et al. The Complete Genome of a New Betabaculovirus from Clostera anastomosis. PLoS ONE 2015, 10, e0132792. [Google Scholar] [CrossRef] [PubMed]
- Miele, S.A.B.; Garavaglia, M.J.; Belaich, M.N.; Ghiringhelli, P.D. Baculovirus: Molecular Insights on Their Diversity and Conservation. Int. J. Evol. Biol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Garavaglia, M.J.; Miele, S.A.B.; Iserte, J.A.; Belaich, M.N.; Ghiringhelli, P.D. The ac53, ac78, ac101, and ac103 Genes Are Newly Discovered Core Genes in the Family Baculoviridae. J. Virol. 2012, 86, 12069–12079. [Google Scholar] [CrossRef] [Green Version]
- Javed, M.A.; Biswas, S.; Willis, L.G.; Harris, S.; Pritchard, C.; Oers, M.M.; van Donly, B.C.; Erlandson, M.A.; Hegedus, D.D.; Theilmann, D.A. Autographa californica Multiple Nucleopolyhedrovirus AC83 is a per os infectivity factor (PIF) protein required for occlusion-derived virus (ODV) and budded virus nucleocapsid assembly as well as assembly of the PIF complex in ODV envelopes. J. Virol. 2017, 91. [Google Scholar] [CrossRef]
- Guarino, L.A.; Xu, B.; Jin, J.; Dong, W. A virus-encoded RNA polymerase purified from baculovirus-infected cells. J. Virol. 1998, 72, 7985–7991. [Google Scholar]
- Jehle, J.A.; Lange, M.; Wang, H.; Hu, Z.; Wang, Y.; Hauschild, R. Molecular identification and phylogenetic analysis of baculoviruses from Lepidoptera. Virology 2006, 346, 180–193. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Niu, Y.D.; Li, J.; Stanford, K.; McAllister, T.A. Rapid and accurate detection of bacteriophage activity against Escherichia coli O157:H7 by propidium monoazide real-time PCR. Biomed. Res. Int. 2014, 2014, 319351. [Google Scholar] [CrossRef] [PubMed]
- Groner, A. Specificity and safety of baculoviruses. In The Biology of Baculoviruses; Granados, R.R., Federici, B.A., Eds.; CRC Press: Boca Raton, FL, USA, 1986; Volume I, pp. 177–201. [Google Scholar]
- Harrison, R.L.; Mowery, J.D.; Rowley, D.L.; Bauchan, G.R.; Theilmann, D.A.; Rohrmann, G.F.; Erlandson, M.A. The complete genome sequence of a third distinct baculovirus isolated from the true armyworm, Mythimna unipuncta, contains two copies of the lef-7 gene. Virus Genes 2018, 54, 297–310. [Google Scholar] [PubMed]
- Harrison, R.L.; Bonning, B.C. The nucleopolyhedroviruses of Rachiplusia ou and Anagrapha falcifera are isolates of the same virus. J. Gen. Virol. 1999, 80, 2793–2798. [Google Scholar] [CrossRef] [PubMed]
- de Moraes, R.R.; Maruniak, J.E. Detection and identification of multiple baculoviruses using the polymerase chain reaction (PCR) and restriction endonuclease analysis. J. Virol. Methods 1997, 63, 209–217. [Google Scholar] [CrossRef]
- Lange, M.; Wang, H.; Zhihong, H.; Jehle, J.A. Towards a molecular identification and classification system of lepidopteran-specific baculoviruses. Virology 2004, 325, 36–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szewczyk, B.; Barski, P.; Sihler, W.; Rabalski, L.; Skrzecz, I.; Hoyos-Carvajal, L.; de Souza, M.L. Detection and identification of baculovirus pesticides by multitemperature single-strand conformational polymorphism. J. Environ. Sci. Health B 2008, 43, 539–545. [Google Scholar] [CrossRef]
- Krejmer-Rabalska, M.; Rabalski, L.; Lobo de Souza, M.; Moore, S.D.; Szewczyk, B. New method for differentiation of granuloviruses (Betabaculoviruses) based on multitemperature single stranded conformational polymorphism. Int. J. Mol. Sci. 2017, 19. [Google Scholar] [CrossRef]
- Kaczanowski, R.; Trzeciak, L.; Kucharczyk, K. Multitemperature single-strand conformation polymorphism. Electrophoresis 2001, 22, 3539–3545. [Google Scholar] [CrossRef]
- Moreau, F.; Fetouchi, R.; Micalessi, I.; Brejeon, V.; Bacon, N.; Jannes, G.; Le Pendeven, C.; Lekbaby, B.; Kremsdorf, D.; Lacau Saint Guily, J.; et al. Detection and genotyping of human papillomavirus by real-time PCR assay. J. Clin. Virol. 2013, 56, 244–249. [Google Scholar] [CrossRef]
- Günes, A.; Marek, A.; Grafl, B.; Berger, E.; Hess, M. Real-time PCR assay for universal detection and quantitation of all five species of fowl adenoviruses (FAdV-A to FAdV-E). J. Virol. Methods 2012, 183, 147–153. [Google Scholar] [CrossRef]
- Arneodo, J.D.; König, G.A.; Berretta, M.F.; Rienzo, J.A.D.; Taboga, O.; Sciocco-Cap, A. Detection and kinetic analysis of Epinotia aporema granulovirus in its lepidopteran host by real-time PCR. Arch. Virol. 2012, 157, 1149–1153. [Google Scholar] [CrossRef]
- Afonso, C.L.; Tulman, E.R.; Lu, Z.; Balinsky, C.A.; Moser, B.A.; Becnel, J.J.; Rock, D.L.; Kutish, G.F. Genome sequence of a baculovirus pathogenic for Culex nigripalpus. J. Virol. 2001, 75, 11157–11165. [Google Scholar] [CrossRef] [PubMed]
- Herniou, E.A.; Olszewski, J.A.; O’Reilly, D.R.; Cory, J.S. Ancient coevolution of baculoviruses and their insect hosts. J. Virol. 2004, 78, 3244–3251. [Google Scholar] [CrossRef] [PubMed]
- Wennmann, J.T.; Keilwagen, J.; Jehle, J.A. Baculovirus Kimura two-parameter species demarcation criterion is confirmed by the distances of 38 core gene nucleotide sequences. J. Gen. Virol. 2018, 99, 1307–1320. [Google Scholar] [CrossRef]
- Brito, A.F.; de Braconi, C.T.; Weidmann, M.; Dilcher, M.; Alves, J.M.P.; Gruber, A.; de Zanotto, P.M.A. The Pangenome of the Anticarsia gemmatalis Multiple Nucleopolyhedrovirus (AgMNPV). Genome Biol. Evol. 2015, 8, 94–108. [Google Scholar] [CrossRef]
- Bonsall, D.; Black, S.; Howe, A.Y.; Chase, R.; Ingravallo, P.; Pak, I.; Brown, A.; Smith, D.A.; Bowden, R.; Barnes, E. Characterization of hepatitis C virus resistance to grazoprevir reveals complex patterns of mutations following on-treatment breakthrough that are not observed at relapse. Infect. Drug Resist. 2018, 11, 1119–1135. [Google Scholar] [CrossRef] [PubMed]
- Cory, J.S.; Green, B.M.; Paul, R.K.; Hunter-Fujita, F. Genotypic and phenotypic diversity of a baculovirus population within an individual insect host. J. Invertebr. Pathol. 2005, 89, 101–111. [Google Scholar] [CrossRef]
- Chateigner, A.; Bézier, A.; Labrousse, C.; Jiolle, D.; Barbe, V.; Herniou, E.A. Ultra Deep Sequencing of a Baculovirus Population Reveals Widespread Genomic Variations. Viruses 2015, 7, 3625–3646. [Google Scholar] [CrossRef] [Green Version]
- Wennmann, J.T.; Radtke, P.; Eberle, K.E.; Gueli Alletti, G.; Jehle, J.A. Deciphering Single Nucleotide Polymorphisms and Evolutionary Trends in Isolates of the Cydia pomonella granulovirus. Viruses 2017, 9. [Google Scholar] [CrossRef]
- Van der Merwe, M.; Jukes, M.D.; Rabalski, L.; Knox, C.; Opoku-Debrah, J.K.; Moore, S.D.; Krejmer-Rabalska, M.; Szewczyk, B.; Hill, M.P. Genome Analysis and Genetic Stability of the Cryptophlebia leucotreta Granulovirus (CrleGV-SA) after 15 Years of Commercial Use as a Biopesticide. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef]
- Harrison, R.L.; Rowley, D.L.; Mowery, J.; Bauchan, G.R.; Theilmann, D.A.; Rohrmann, G.F.; Erlandson, M.A. The Complete Genome Sequence of a Second Distinct Betabaculovirus from the True Armyworm, Mythimna unipuncta. PLoS ONE 2017, 12, e0170510. [Google Scholar] [CrossRef] [PubMed]
- Asser-Kaiser, S.; Fritsch, E.; Undorf-Spahn, K.; Kienzle, J.; Eberle, K.E.; Gund, N.A.; Reineke, A.; Zebitz, C.P.W.; Heckel, D.G.; Huber, J.; et al. Rapid emergence of baculovirus resistance in codling moth due to dominant, sex-linked inheritance. Science 2007, 317, 1916–1918. [Google Scholar] [CrossRef] [PubMed]
- Alletti, G.G.; Sauer, A.J.; Weihrauch, B.; Fritsch, E.; Undorf-Spahn, K.; Wennmann, J.T.; Jehle, J.A. Using next generation sequencing to identify and quantify the genetic composition of resistance-breaking commercial isolates of Cydia pomonella granulovirus. Viruses 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Asser-Kaiser, S.; Heckel, D.G.; Jehle, J.A. Sex linkage of CpGV resistance in a heterogeneous field strain of the codling moth Cydia pomonella (L.). J. Invertebr. Pathol. 2010, 103, 59–64. [Google Scholar] [CrossRef]
- Asser-Kaiser, S.; Radtke, P.; El-Salamouny, S.; Winstanley, D.; Jehle, J.A. Baculovirus resistance in codling moth (Cydia pomonella L.) caused by early block of virus replication. Virology 2011, 410, 360–367. [Google Scholar] [CrossRef]
- Jehle, J.A.; Schulze-Bopp, S.; Undorf-Spahn, K.; Fritsch, E. Evidence for a second type of resistance against Cydia pomonella granulovirus in field populations of codling moths. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef]
- Sauer, A.J.; Schulze-Bopp, S.; Fritsch, E.; Undorf-Spahn, K.; Jehle, J.A. A third type of resistance to Cydia pomonella granulovirus in codling moths shows a mixed Z-linked and autosomal inheritance pattern. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abrreviation | Betabaculovirus Name | Host Family | GenBank Accession Number | Genome Size (bp) |
|---|---|---|---|---|
| AdorGV | Adoxophyes orana granulovirus | Tortricidae | AF547984 | 99,657 |
| AgseGV | Agrotis segetum granulovirus | Noctuidae | AY522332 | 131,680 |
| ChocGV | Choristoneura occidentalis granulovirus | Tortricidae | DQ333351 | 104,710 |
| ClanGV | Clostera anachoreta granulovirus | Notodontidae | HQ116624 | 101,487 |
| ClasGV | Clostera anastomosis granulovirus | Notodontidae | KC179784 | 101,818 |
| ClasGV-B | Clostera anastomosis granulovirus B | Notodontidae | KR091910 | 107,439 |
| CnmeGV | Cnaphalocrocis medinalis granulovirus | Crambidae | KU593505 | 111,246 |
| CpGV | Cydia pomonella granulovirus | Tortricidae | U53466 | 123,500 |
| CrleGV | Cryptophlebia leucotreta granulovirus | Tortricidae | AY229987 | 110,907 |
| DisaGV | Diatraea saccharalis granulovirus | Crambidae | KP296186 | 98,392 |
| EpapGV | Epinotia aporema granulovirus | Tortricidae | JN408834 | 119,082 |
| ErelGV | Erinnyis ello granulovirus | Sphingidae | KJ406702 | 102,759 |
| HearGV | Helicoverpa armigera granulovirus | Noctuidae | EU255577 | 169,794 |
| MolaGV | Mocis latipes granulovírus | Noctuidae | KR011718 | 134,272 |
| MyunGV-A | Mythimna unipuncta granulovirus A | Noctuidae | EU678671 | 176,677 |
| MyunGV-B | Mythimna unipuncta granulovírus B | Noctuidae | KX855660 | 144,673 |
| PhopGV | Phthorimaea operculella granulovirus | Gelechiidae | AF499596 | 119,217 |
| PiGV | Plodia interpunctella granulovirus | Pyralidae | KX151395 | 112,536 |
| PlxyGV | Plutella xylostella granulovirus | Plutellidae | AF270937 | 100,999 |
| PrGV | Pieris rapae granulovirus isolate | Pieridae | GQ884143 | 108,592 |
| SfGV | Spodoptera frugiperda granulovirus | Noctuidae | KM371112 | 140,913 |
| SpliGV | Spodoptera litura granulovirus | Noctuidae | DQ288858 | 124,121 |
| TnGV | Trichoplusia ni granulovirus | Noctuidae | KU752557 | 175,360 |
| XcGV | Xestia c-nigrum granulovirus | Noctuidae | AF162221 | 178,733 |
| Isolate | Tm1 (°C) | Tm2 (°C) | Tm3 (°C) | Mean Tm (°C) | SD | |ΔMean Tm| (°C) | SNPs | |
|---|---|---|---|---|---|---|---|---|
| gran | Neg(-) | - | - | - | - | - | - | |
| CrleGV-Eu | 80.97 | 81.04 | 81.04 | 81.02 | 0.03 | 0.95 | 1 | |
| CrleGV-SA | 82.05 | 81.92 | 81.95 | 81.97 | 0.06 | |||
| lef-8 | Neg(-) | - | - | - | - | - | - | |
| CrleGV-Eu | 75.73 | 75.77 | 75.81 | 75.77 | 0.03 | 1.09 | 2 | |
| CrleGV-SA | 76.89 | 76.85 | 76.84 | 76.86 | 0.02 | |||
| lef-9 | Neg(-) | - | - | - | - | - | - | |
| CrleGV-Eu | 77.14 | 77.23 | 77.25 | 77.21 | 0.05 | 0.29 | 3 | |
| CrleGV-SA | 77.53 | 77.45 | 77.53 | 77.50 | 0.04 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krejmer-Rabalska, M.; Rabalski, L.; Jukes, M.D.; Lobo de Souza, M.; Moore, S.D.; Szewczyk, B. New Method for Differentiation of Granuloviruses (Betabaculoviruses) Based on Real-Time Polymerase Chain Reaction (Real-Time PCR). Viruses 2019, 11, 115. https://doi.org/10.3390/v11020115
Krejmer-Rabalska M, Rabalski L, Jukes MD, Lobo de Souza M, Moore SD, Szewczyk B. New Method for Differentiation of Granuloviruses (Betabaculoviruses) Based on Real-Time Polymerase Chain Reaction (Real-Time PCR). Viruses. 2019; 11(2):115. https://doi.org/10.3390/v11020115
Chicago/Turabian StyleKrejmer-Rabalska, Martyna, Lukasz Rabalski, Michael D. Jukes, Marlinda Lobo de Souza, Sean D. Moore, and Boguslaw Szewczyk. 2019. "New Method for Differentiation of Granuloviruses (Betabaculoviruses) Based on Real-Time Polymerase Chain Reaction (Real-Time PCR)" Viruses 11, no. 2: 115. https://doi.org/10.3390/v11020115
APA StyleKrejmer-Rabalska, M., Rabalski, L., Jukes, M. D., Lobo de Souza, M., Moore, S. D., & Szewczyk, B. (2019). New Method for Differentiation of Granuloviruses (Betabaculoviruses) Based on Real-Time Polymerase Chain Reaction (Real-Time PCR). Viruses, 11(2), 115. https://doi.org/10.3390/v11020115