IIV-6 Inhibits NF-?B Responses in Drosophila

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. RNA Isolation and qRT-PCR
2.2. The nCounter Analysis
2.3. Fly Stocks and In Vivo Studies
2.4. Cell Culture
2.5. Immunoblots
2.6. Confocal Microscopy
2.7. Virus Preparation
3. Results
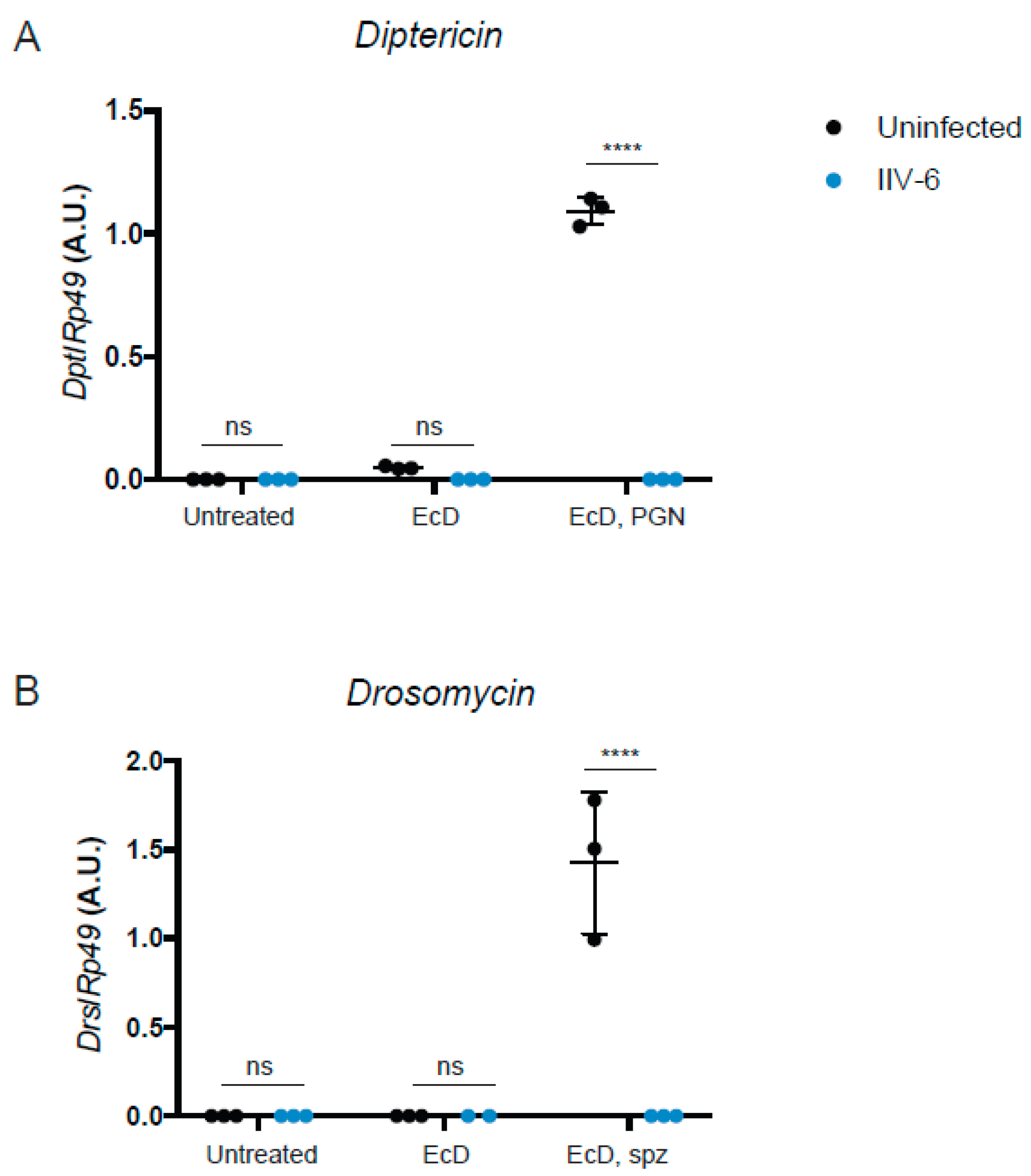
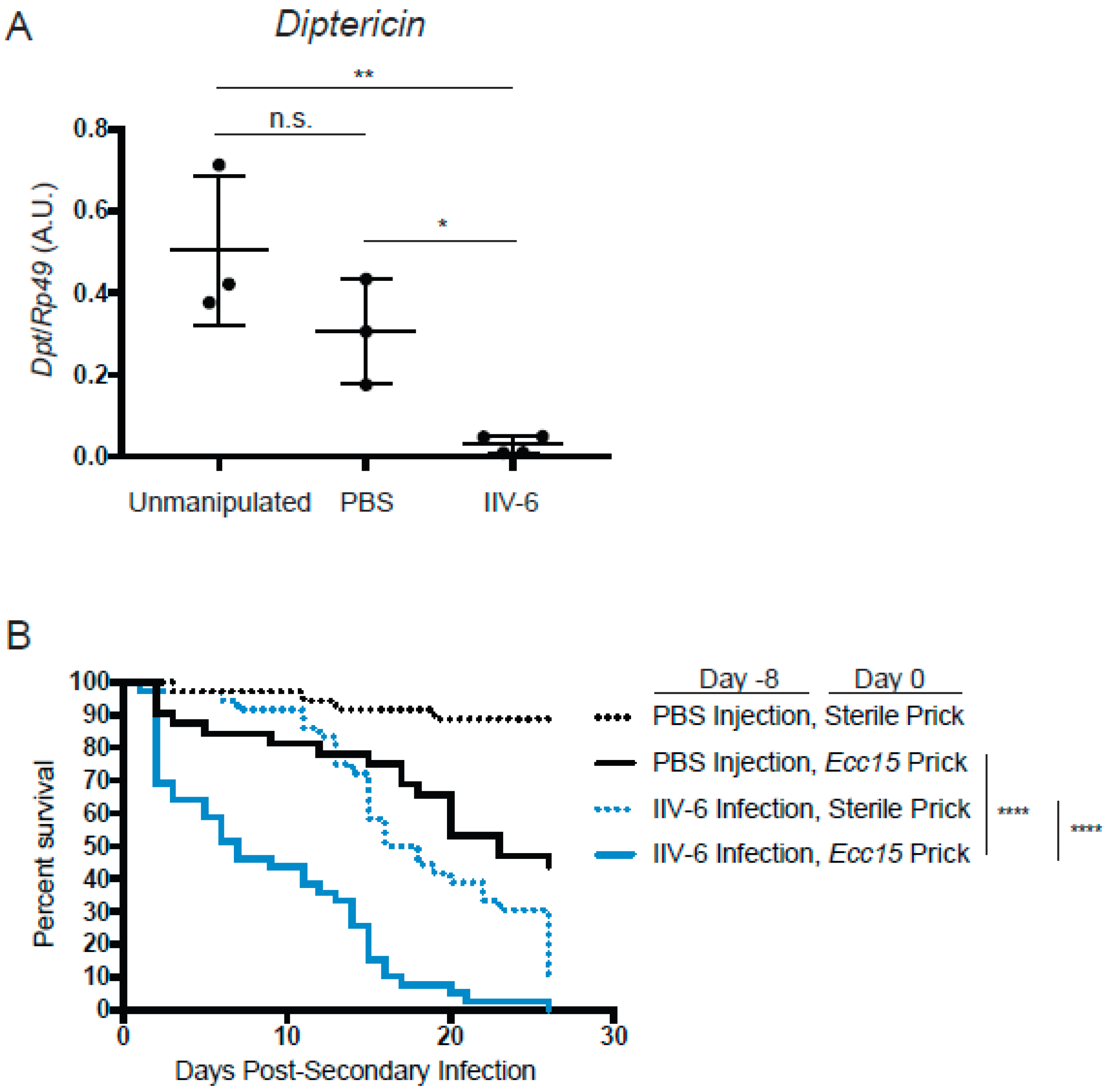
3.1. AMP Production is Suppressed in the Presence of IIV-6
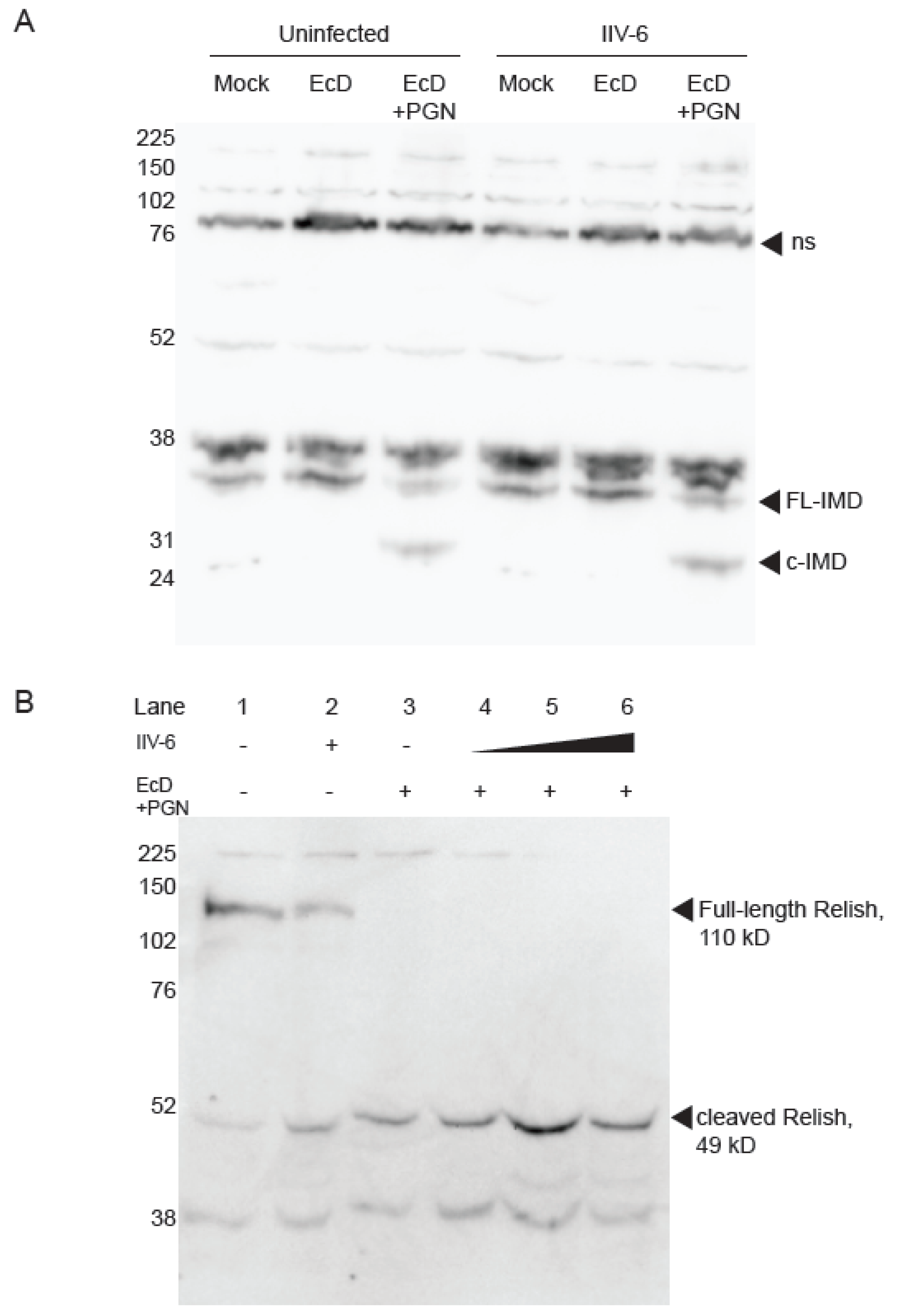
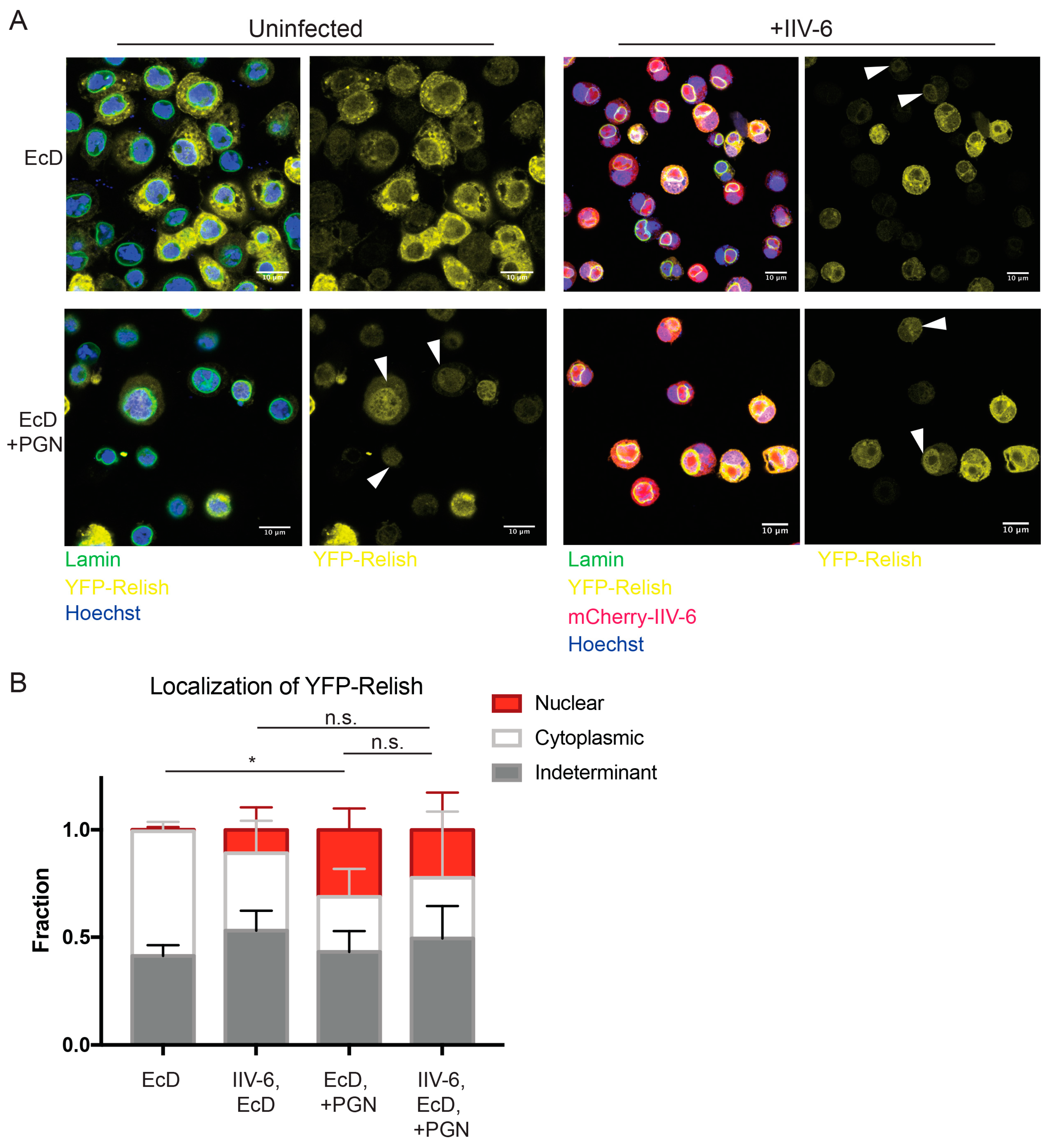
3.2. NF-κB Inhibition is Downstream of Imd and Relish
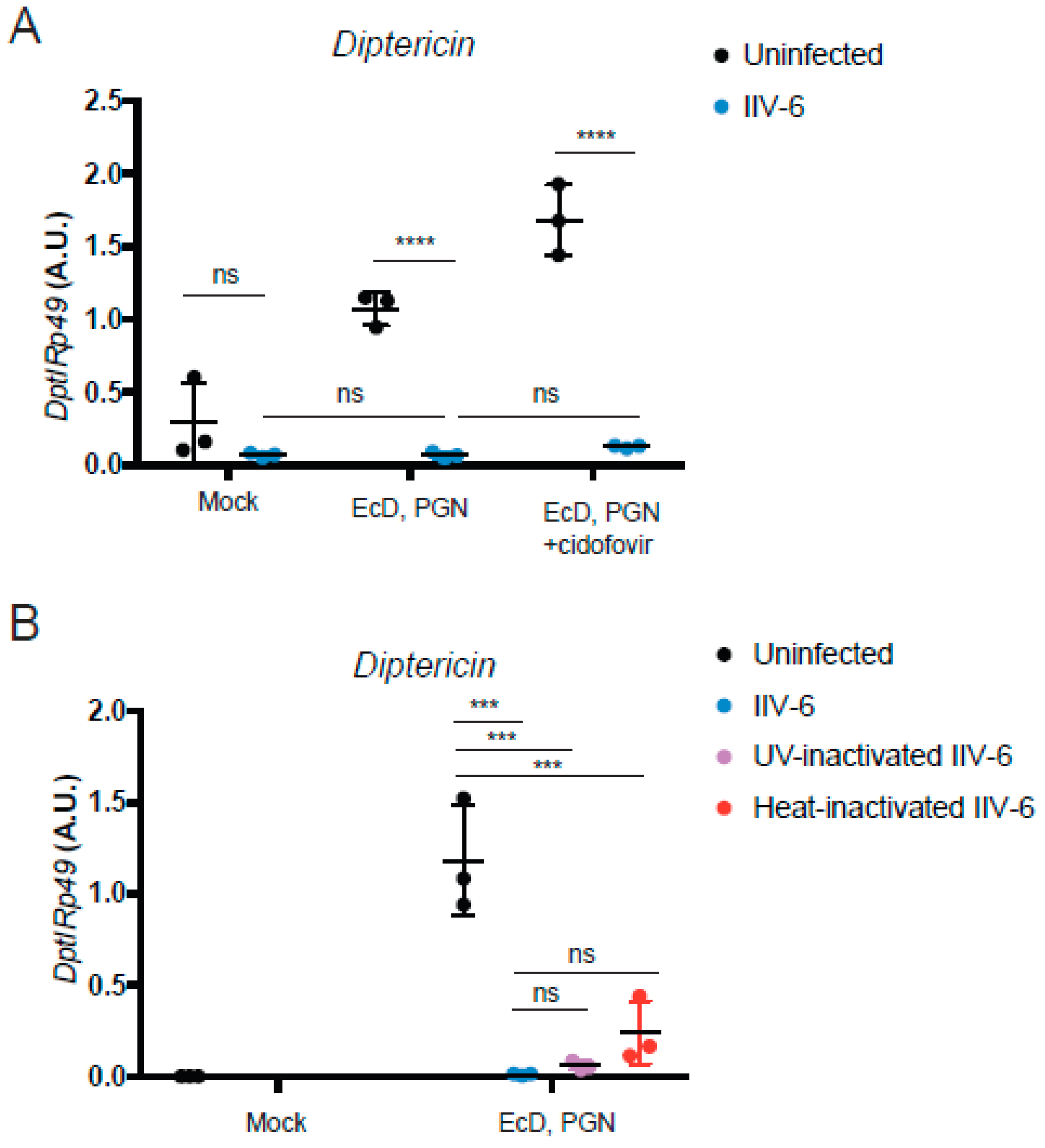
3.3. Inhibition of NF-κB Signaling is Mediated by an Immediate Early Gene
3.4. Flies Infected with IIV-6 are more Susceptible to Bacterial Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Valen, L.V. A new evolutionary law. Evol. Theory 1973, 1, 1–30. [Google Scholar]
- Lamiable, O.; Kellenberger, C.; Kemp, C.; Troxler, L.; Pelte, N.; Boutros, M.; Marques, J.T.; Daeffler, L.; Hoffmann, J.A.; Roussel, A.; et al. Cytokine diedel and a viral homologue suppress the imd pathway in drosophila. Proc. Natl. Acad. Sci. USA 2016, 113, 698–703. [Google Scholar] [CrossRef]
- Smith, G.L.; Benfield, C.T.; Maluquer de Motes, C.; Mazzon, M.; Ember, S.W.; Ferguson, B.J.; Sumner, R.P. Vaccinia virus immune evasion: Mechanisms, virulence and immunogenicity. J. Gen. Virol. 2013, 94, 2367–2392. [Google Scholar] [CrossRef]
- Palmer, W.H.; Joosten, J.; Overheul, G.J.; Jansen, P.W.; Vermeulen, M.; Obbard, D.J.; van Rij, R.P. Induction and suppression of nf-kappab signalling by a DNA virus of drosophila. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Teixeira, L.; Ferreira, Á.; Ashburner, M. The bacterial symbiont wolbachia induces resistance to rna viral infections in drosophila melanogaster. PLoS Biol. 2008, 6, e1000002. [Google Scholar] [CrossRef]
- Bronkhorst, A.W.; Van Cleef, K.W.R.; Vodovar, N.; Ince, I.A.; Blanc, H.; Vlak, J.M.; Saleh, M.-C.; van Rij, R.P. The DNA virus invertebrate iridescent virus 6 is a target of the drosophila rnai machinery. Proc. Natl. Acad. Sci. USA 2012, 109, E3604–E3613. [Google Scholar] [CrossRef]
- Ince, I.A.; Boeren, S.A.; van Oers, M.M.; Vervoort, J.J.M.; Vlak, J.M. Proteomic analysis of chilo iridescent virus. Virology 2010, 405, 253–258. [Google Scholar] [CrossRef]
- Bronkhorst, A.W.; Van Cleef, K.W.R.; Venselaar, H.; Van Rij, R.P. A dsrna-binding protein of a complex invertebrate DNA virus suppresses the drosophila rnai response. Nucleic Acids Res. 2014, 42, 12237–12248. [Google Scholar] [CrossRef]
- Chitnis, N.S.; D’Costa, S.M.; Paul, E.R.; Bilimoria, S.L. Modulation of iridovirus-induced apoptosis by endocytosis, early expression, jnk, and apical caspase. Virology 2008, 370, 333–342. [Google Scholar] [CrossRef]
- Chitnis, N.S.; Paul, E.R.; Lawrence, P.K.; Henderson, C.W.; Ganapathy, S.; Taylor, P.V.; Virdi, K.S.; D’Costa, S.M.; May, A.R.; Bilimoria, S.L. A virion-associated protein kinase induces apoptosis. J. Virol. 2011, 85, 13144–13152. [Google Scholar] [CrossRef]
- Avadhanula, V.; Weasner, B.P.; Hardy, G.G.; Kumar, J.P.; Hardy, R.W. A novel system for the launch of alphavirus rna synthesis reveals a role for the imd pathway in arthropod antiviral response. PLoS Pathog. 2009, 5, e1000582. [Google Scholar] [CrossRef]
- Costa, A.; Jan, E.; Sarnow, P.; Schneider, D. The imd pathway is involved in antiviral immune responses in drosophila. PLoS ONE 2009, 4, e7436. [Google Scholar] [CrossRef]
- Goto, A.; Okado, K.; Martins, N.; Cai, H.; Barbier, V.; Lamiable, O.; Troxler, L.; Santiago, E.; Kuhn, L.; Paik, D.; et al. The kinase ikkbeta regulates a sting- and nf-kappab-dependent antiviral response pathway in drosophila. Immunity 2018, 49, 225–234 e224. [Google Scholar] [CrossRef]
- Liu, Y.; Gordesky-Gold, B.; Leney-Greene, M.; Weinbren, N.L.; Tudor, M.; Cherry, S. Inflammation-induced, sting-dependent autophagy restricts zika virus infection in the drosophila brain. Cell Host Microbe 2018, 24, 57–68 e53. [Google Scholar] [CrossRef]
- Lemaitre, B.; Hoffmann, J. The host defense of drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef]
- Clemmons, A.W.; Lindsay, S.A.; Wasserman, S.A. An effector peptide family required for drosophila toll-mediated immunity. PLoS Pathog. 2015, 11, e1004876. [Google Scholar] [CrossRef]
- Lindsay, S.A.; Lin, S.J.H.; Wasserman, S.A. Short-form bomanins mediate humoral immunity in drosophila. J. Innate Immun. 2018, 10, 306–314. [Google Scholar] [CrossRef]
- Hanson, M.A.; Dostalova, A.; Ceroni, C.; Poidevin, M.; Kondo, S.; Lemaitre, B. Synergy and remarkable specificity of antimicrobial peptides in vivo using a systematic knockout approach. Elife 2019, 8. [Google Scholar] [CrossRef]
- Zambon, R.A.; Nandakumar, M.; Vakharia, V.N.; Wu, L.P. The toll pathway is important for an antiviral response in drosophila. Proc. Natl. Acad. Sci. USA 2005, 102, 7257–7262. [Google Scholar] [CrossRef]
- Tsai, C.W.; McGraw, E.A.; Ammar, E.D.; Dietzgen, R.G.; Hogenhout, S.A. Drosophila melanogaster mounts a unique immune response to the rhabdovirus sigma virus. Appl. Environ. Microbiol. 2008, 74, 3251–3256. [Google Scholar] [CrossRef]
- Huang, Z.; Kingsolver, M.B.; Avadhanula, V.; Hardy, R.W. An antiviral role for antimicrobial peptides during the arthropod response to alphavirus replication. J. Virol. 2013, 87, 4272–4280. [Google Scholar] [CrossRef]
- West, C.; Silverman, N. P38b and jak-stat signaling protect against invertebrate iridescent virus 6 infection in drosophila. PLoS Pathog. 2018, 14, e1007020. [Google Scholar] [CrossRef]
- Stoven, S.; Silverman, N.; Junell, A.; Hedengren-Olcott, M.; Erturk, D.; Engstrom, Y.; Maniatis, T.; Hultmark, D. Caspase-mediated processing of the drosophila nf-kappab factor relish. Proc. Natl. Acad. Sci. USA 2003, 100, 5991–5996. [Google Scholar] [CrossRef]
- Paquette, N.; Broemer, M.; Aggarwal, K.; Chen, L.; Husson, M.; Ertürk-Hasdemir, D.; Reichhart, J.-M.; Meier, P.; Silverman, N. Caspase-mediated cleavage, iap binding, and ubiquitination: Linking three mechanisms crucial for drosophila nf-kappab signaling. Mol. Cell 2010, 37, 172–182. [Google Scholar] [CrossRef]
- Samakovlis, C.; Åsling, B.; Boman, H.G.; Gateff, E.; Hultmark, D. In vitro induction of cecropin genes--an immune response in a drosophila blood cell line. Biochem. Biophys. Res. Commun. 1992, 188, 1169–1175. [Google Scholar] [CrossRef]
- Silverman, N.; Zhou, R.; Stoven, S.; Pandey, N.; Hultmark, D.; Maniatis, T. A drosophila ikappab kinase complex required for relish cleavage and antibacterial immunity. Genes Dev. 2000, 14, 2461–2471. [Google Scholar] [CrossRef]
- Ertürk-Hasdemir, D.; Broemer, M.; Leulier, F.; Lane, W.S.; Paquette, N.; Hwang, D.; Kim, C.-H.; Stöven, S.; Meier, P.; Silverman, N. Two roles for the drosophila ikk complex in the activation of relish and the induction of antimicrobial peptide genes. Proc. Natl. Acad.Sci. USA 2009, 106, 9779–9784. [Google Scholar] [CrossRef] [PubMed]
- Rus, F.; Flatt, T.; Tong, M.; Aggarwal, K.; Okuda, K.; Kleino, A.; Yates, E.; Tatar, M.; Silverman, N. Ecdysone triggered pgrp-lc expression controls drosophila innate immunity. EMBO J. 2013, 32, 1626–1638. [Google Scholar] [CrossRef]
- Ozgen, A.; Muratoglu, H.; Demirbağ, Z.; Vlak, J.M.; van Oers, M.M.; Nalçacioğlu, R. Construction and characterization of a recombinant invertebrate iridovirus. Virus Res. 2014. [Google Scholar] [CrossRef]
- Nalçacioğlu, R.; Marks, H.; Vlak, J.M.; Demirbağ, Z.; van Oers, M.M. Promoter analysis of the chilo iridescent virus DNA polymerase and major capsid protein genes. Virology 2003, 317, 321–329. [Google Scholar] [CrossRef]
- Davis, M.M.; Primrose, D.A.; Hodgetts, R.B. A member of the p38 mitogen-activated protein kinase family is responsible for transcriptional induction of dopa decarboxylase in the epidermis of drosophila melanogaster during the innate immune response. Mol. Cell. Biol. 2008, 28, 4883–4895. [Google Scholar] [CrossRef]
- Sabin, L.R.; Zhou, R.; Gruber, J.J.; Lukinova, N.; Bambina, S.; Berman, A.; Lau, C.-K.; Thompson, C.B.; Cherry, S. Ars2 regulates both mirna- and sirna- dependent silencing and suppresses rna virus infection in drosophila. Cell 2009, 138, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Silverman, N.; Zhou, R.; Erlich, R.L.; Hunter, M.; Bernstein, E.; Schneider, D.; Maniatis, T. Immune activation of nf-kappab and jnk requires drosophila tak1. J. Biol. Chem. 2003, 278, 48928–48934. [Google Scholar] [CrossRef]
- Kim, C.H.; Paik, D.; Rus, F.; Silverman, N. The caspase-8 homolog dredd cleaves imd and relish but is not inhibited by p35. J. Biol. Chem. 2014, 289, 20092–20101. [Google Scholar] [CrossRef]
- Sumner, R.P.; Maluquer de Motes, C.; Veyer, D.L.; Smith, G.L. Vaccinia virus inhibits nf- b-dependent gene expression downstream of p65 translocation. J. Virol. 2014, 88, 3092–3102. [Google Scholar] [CrossRef] [PubMed]
- Revilla, Y.; Callejo, M.; Rodriguez, J.M.; Culebras, E.; Nogal, M.L.; Salas, M.L.; Vinuela, E.; Fresno, M. Inhibition of nuclear factor kappab activation by a virus-encoded ikappab-like protein. J. Biol. Chem. 1998, 273, 5405–5411. [Google Scholar] [CrossRef]
- Stuart, J.H.; Sumner, R.P.; Lu, Y.; Snowden, J.S.; Smith, G.L. Vaccinia virus protein c6 inhibits type i ifn signalling in the nucleus and binds to the transactivation domain of stat2. PLoS Pathog. 2016, 12, e1005955. [Google Scholar] [CrossRef]
- Ferguson, B.J.; Benfield, C.T.; Ren, H.; Lee, V.H.; Frazer, G.L.; Strnadova, P.; Sumner, R.P.; Smith, G.L. Vaccinia virus protein n2 is a nuclear irf3 inhibitor that promotes virulence. J. Gen. Virol. 2013, 94, 2070–2081. [Google Scholar] [CrossRef]
- Kleino, A.; Ramia, N.F.; Bozkurt, G.; Shen, Y.; Nailwal, H.; Huang, J.; Napetschnig, J.; Gangloff, M.; Chan, F.K.; Wu, H.; et al. Peptidoglycan-sensing receptors trigger the formation of functional amyloids of the adaptor protein imd to initiate drosophila nf-kappab signaling. Immunity 2017, 47, 635–647 e636. [Google Scholar] [CrossRef] [PubMed]
- Upton, J.W.; Kaiser, W.J.; Mocarski, E.S. Cytomegalovirus m45 cell death suppression requires receptor-interacting protein (rip) homotypic interaction motif (rhim)-dependent interaction with rip1. J. Biol. Chem. 2008, 283, 16966–16970. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Muller, A.; Sumser, H.; Horren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef]
- Tokarz, R.; Firth, C.; Street, C.; Cox-Foster, D.L.; Lipkin, W.I. Lack of evidence for an association between iridovirus and colony collapse disorder. PLoS ONE 2011, 6, e21844. [Google Scholar] [CrossRef]
- Kaneko, T.; Goldman, W.E.; Mellroth, P.; Steiner, H.; Fukase, K.; Kusumoto, S.; Harley, W.; Fox, A.; Golenbock, D.; Silverman, N. Monomeric and polymeric gram-negative peptidoglycan but not purified lps stimulate the drosophila imd pathway. Immunity 2004, 20, 637–649. [Google Scholar] [CrossRef]
- Kaneko, T.; Yano, T.; Aggarwal, K.; Lim, J.-H.; Ueda, K.; Oshima, Y.; Peach, C.; Erturk-Hasdemir, D.; Goldman, W.E.; Oh, B.-H.; et al. Pgrp-lc and pgrp-le have essential yet distinct functions in the drosophila immune response to monomeric dap-type peptidoglycan. Nat. Immunol. 2006, 7, 715–723. [Google Scholar] [CrossRef]
- Sansone, C.L.; Cohen, J.; Yasunaga, A.; Xu, J.; Osborn, G.; Subramanian, H.; Gold, B.; Buchon, N.; Cherry, S. Microbiota-dependent priming of antiviral intestinal immunity in drosophila. Cell Host Microbe 2015, 18, 571–581. [Google Scholar] [CrossRef]
- McGettigan, J.; McLennan, R.K.; Broderick, K.E.; Kean, L.; Allan, A.K.; Cabrero, P.; Regulski, M.R.; Pollock, V.P.; Gould, G.W.; Davies, S.A.; et al. Insect renal tubules constitute a cell-autonomous immune system that protects the organism against bacterial infection. Insect Biochem. Mol. Biol. 2005, 35, 741–754. [Google Scholar] [CrossRef]
- Verma, P.; Tapadia, M.G. Immune response and anti-microbial peptides expression in malpighian tubules of drosophila melanogaster is under developmental regulation. PLoS ONE 2012, 7, e40714. [Google Scholar] [CrossRef]
- Paik, D.; Monahan, A.; Caffrey, D.R.; Elling, R.; Goldman, W.E.; Silverman, N. Slc46 family transporters facilitate cytosolic innate immune recognition of monomeric peptidoglycans. J. Immunol. 2017, 199, 263–270. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
West, C.; Rus, F.; Chen, Y.; Kleino, A.; Gangloff, M.; Gammon, D.B.; Silverman, N. IIV-6 Inhibits NF-?B Responses in Drosophila. Viruses 2019, 11, 409. https://doi.org/10.3390/v11050409
West C, Rus F, Chen Y, Kleino A, Gangloff M, Gammon DB, Silverman N. IIV-6 Inhibits NF-?B Responses in Drosophila. Viruses. 2019; 11(5):409. https://doi.org/10.3390/v11050409
Chicago/Turabian StyleWest, Cara, Florentina Rus, Ying Chen, Anni Kleino, Monique Gangloff, Don B. Gammon, and Neal Silverman. 2019. "IIV-6 Inhibits NF-?B Responses in Drosophila" Viruses 11, no. 5: 409. https://doi.org/10.3390/v11050409
APA StyleWest, C., Rus, F., Chen, Y., Kleino, A., Gangloff, M., Gammon, D. B., & Silverman, N. (2019). IIV-6 Inhibits NF-?B Responses in Drosophila. Viruses, 11(5), 409. https://doi.org/10.3390/v11050409