Equine Parvovirus-Hepatitis Frequently Detectable in Commercial Equine Serum Pools

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Serum Sample Collection
2.2. Detection of EqPV-H DNA
2.3. Detection of Anti-EqPV-H Antibodies
2.4. Sequencing and Phylogeny
2.5. Purification of Viral Particles
3. Results and Discussion
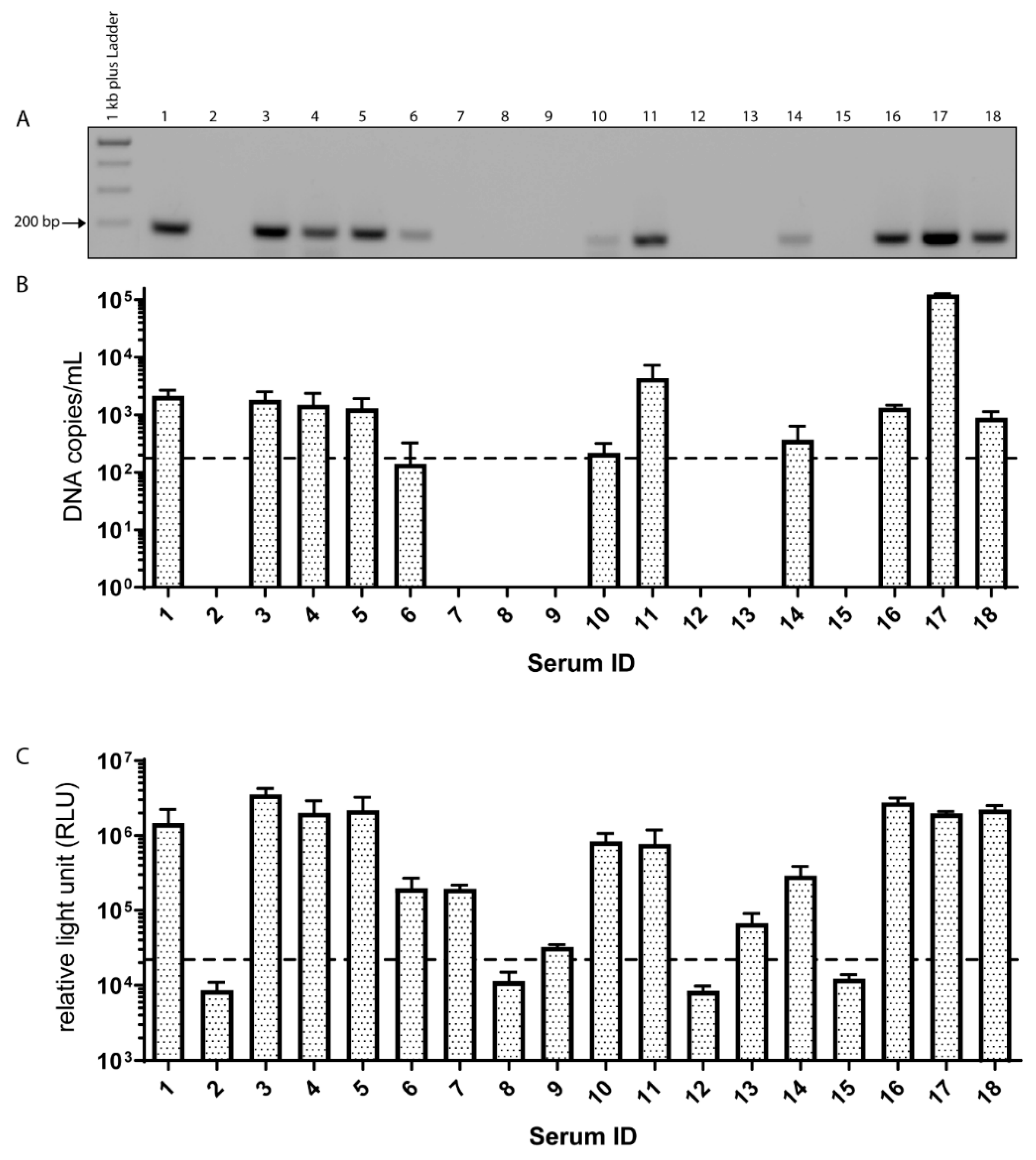
3.1. EqPV-H DNA is Frequently Detectable in Commercial Horse Sera
3.2. EqPV-H Can be Detected in Different Countries Worldwide
3.3. Sequence and Phylogenetic Analysis of EqPV-H in Commercial Serum Pools
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cotmore, S.F.; Agbandje-McKenna, M.; Chiorini, J.A.; Mukha, D.V.; Pintel, D.J.; Qiu, J.; Soderlund-Venermo, M.; Tattersall, P.; Tijssen, P.; Gatherer, D.; et al. The family Parvoviridae. Arch. Virol. 2014, 159, 1239–1247. [Google Scholar] [CrossRef]
- Kailasan, S.; Agbandje-McKenna, M.; Parrish, C.R. Parvovirus Family Conundrum, What Makes a Killer? Annu. Rev. Virol. 2015, 2, 425–450. [Google Scholar] [CrossRef]
- Palinski, R.M.; Mitra, N.; Hause, B.M. Discovery of a novel Parvovirinae virus, porcine parvovirus 7, by metagenomic sequencing of porcine rectal swabs. Virus Genes 2016, 52, 564–567. [Google Scholar] [CrossRef]
- Lau, S.K.P.; Woo, P.C.Y.; Yeung, H.C.; Teng, J.L.L.; Wu, Y.; Bai, R.; Fan, R.Y.Y.; Chan, K.-H.; Yuen, K.-Y. Identification and characterization of bocaviruses in cats and dogs reveals a novel feline bocavirus and a novel genetic group of canine bocavirus. J. Gen. Virol. 2012, 93, 1573–1582. [Google Scholar] [CrossRef] [Green Version]
- Divers, T.J.; Tennant, B.C.; Kumar, A.; McDonough, S.; Cullen, J.; Bhuva, N.; Jain, K.; Chauhan, L.S.; Scheel, T.K.H.; Lipkin, W.I.; et al. New Parvovirus Associated with Serum Hepatitis in Horses after Inoculation of Common Biological Product. Emerg. Infect. Dis. 2018, 24, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Divers, T.J.; Tomlinson, J.E. Theiler’s disease. Equine Vet Educ. 2019, 19, 120. [Google Scholar] [CrossRef]
- Lu, G.; Sun, L.; Ou, J.; Xu, H.; Wu, L.; Li, S. Identification and genetic characterization of a novel parvovirus associated with serum hepatitis in horses in China. Emerg. Microbes & Infect. 2018, 7, 170. [Google Scholar]
- Theiler, A. Acute Liver-Atrophy and Parenchymatous Hepatitis in Horses; Reports of the Director of Veterinary Research; CAB Direct: Glasgow, UK, 1918; pp. 7–164. [Google Scholar]
- Aleman, M.; Nieto, J.E.; Carr, E.A.; Carlson, G.P. Serum Hepatitis Associated with Commercial Plasma Transfusion in Horses. J. Vet. Intern. Med. 2005, 19, 120–122. [Google Scholar] [CrossRef]
- Thomsett, L.R. Acute Hepatic Failure in the Horse. Equine Vet. J. 1971, 3, 15–19. [Google Scholar] [CrossRef]
- Chandriani, S.; Skewes-Cox, P.; Zhong, W.; Ganem, D.E.; Divers, T.J.; van Blaricum, A.J.; Tennant, B.C.; Kistler, A.L. Identification of a previously undescribed divergent virus from the Flaviviridae family in an outbreak of equine serum hepatitis. Proc. Natl. Acad. Sci. USA 2013, 110, E1407–E1415. [Google Scholar] [CrossRef] [Green Version]
- Rose, J.A.; Immenschuh, R.D.; Rose, E.M. Serum hepatitis in the horse, Proceedings of the Twentieth Annual Conference of the American Association of Equine Practitioners. Am. Assoc. Equine Pract. 1974, 175–185. [Google Scholar]
- Tomlinson, J.E.; Kapoor, A.; Kumar, A.; Tennant, B.C.; Laverack, M.A.; Beard, L.; Delph, K.; Davis, E.; Schott Ii, H.; Lascola, K.; et al. Viral testing of 18 consecutive cases of equine serum hepatitis, A prospective study (2014–2018). J. Vet. Intern. Med. 2019, 33, 251–257. [Google Scholar] [CrossRef]
- Tomlinson, J.E.; Tennant, B.C.; Struzyna, A.; Mrad, D.; Browne, N.; Whelchel, D.; Johnson, P.J.; Jamieson, C.; Löhr, C.V.; Bildfell, R.; et al. Viral testing of 10 cases of Theiler’s disease and 37 in-contact horses in the absence of equine biologic product administration, A prospective study (2014–2018). J. Vet. Intern. Med. 2019, 33, 258–265. [Google Scholar] [CrossRef]
- Burbelo, P.D.; Ching, K.H.; Klimavicz, C.M.; Iadarola, M.J. Antibody profiling by Luciferase Immunoprecipitation Systems (LIPS). J. Vis. Exp. 2009, 32, e1549. [Google Scholar] [CrossRef] [PubMed]
- Burbelo, P.D.; Dubovi, E.J.; Simmonds, P.; Medina, J.L.; Henriquez, J.A.; Mishra, N.; Wagner, J.; Tokarz, R.; Cullen, J.M.; Iadarola, M.J.; et al. Serology-enabled discovery of genetically diverse hepaciviruses in a new host. J. Virol. 2012, 86, 6171–6178. [Google Scholar] [CrossRef] [PubMed]
- Pfaender, S.; Cavalleri, J.M.V.; Walter, S.; Doerrbecker, J.; Campana, B.; Brown, R.J.P.; Burbelo, P.D.; Postel, A.; Hahn, K.; Anggakusuma, R.N.; et al. Clinical course of infection and viral tissue tropism of hepatitis C virus-like nonprimate hepaciviruses in horses. Hepatology (Baltimore, Md.) 2015, 61, 447–459. [Google Scholar] [CrossRef] [Green Version]
- Waddell, P.J.; Steel, M.A. General time-reversible distances with unequal rates across sites, Mixing gamma and inverse Gaussian distributions with invariant sites. Mol. Phylogenetics Evol. 1997, 8, 398–414. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences, A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X, Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Stang, A.; Korn, K.; Wildner, O.; Uberla, K. Characterization of virus isolates by particle-associated nucleic acid PCR. J. Clin. Microbiol. 2005, 43, 716–720. [Google Scholar] [CrossRef]
- Postel, A.; Cavalleri, J.-M.V.; Pfaender, S.; Walter, S.; Steinmann, E.; Fischer, N.; Feige, K.; Haas, L.; Becher, P. Frequent presence of hepaci and pegiviruses in commercial equine serum pools. Vet. Microbiol. 2016, 182, 8–14. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| PCR | Primer | Sequence |
|---|---|---|
| I | EPVf1 | GGGTGGTAAATGCCTTCG |
| EPVseqr01 | TGGTTGGTGACGCCTGTC | |
| II | EPVseqf01 | GACAGGCGTCACCAACCA |
| 1104R1 | GGGAATGTCATTGAACGGGAA |
| Serum ID | Serum Type | Origin | Number of Individuals |
|---|---|---|---|
| 1 | Horse Serum | New Zealand | donor herd |
| 2 | Fetal Horse Serum | Mexico | unknown |
| 3 | Donor Horse Serum | USA | pooled |
| 4 | Horse Serum | USA | donor herd |
| 5 | Horse Serum | USA | donor herd |
| 6 | Donor Horse Serum | Italy | pooled |
| 7 | Horse Serum | France | pooled |
| 8 | Donor Horse Serum | Canada | unknown |
| 9 | Donor Horse Serum | Chile | unknown |
| 10 | Donor Horse Serum | Germany | unknown |
| 11 | Donor Horse Serum | Italy | unknown |
| 12 | Fetal Horse Serum | Brazil | unknown |
| 13 | Donor Horse Serum (heat inactivated) | Chile | unknown |
| 14 | Horse Serum | Europe | pooled |
| 15 | Donor Horse Serum | Europe | pooled |
| 16 | Donor Horse Serum | USA | unknown |
| 17 | Horse Serum | New Zealand | unknown |
| 18 | Donor Horse Serum | Canada | unknown |
| Serum ID | EqPV Sequence Name | NCBI Accession Number |
|---|---|---|
| 1 | Equine Parvovirus H/seq1_NZ_518-1104nt | MK792430 |
| 3 | Equine Parvovirus H/seq3_USA_518-1104nt | MK792431 |
| 4 | Equine Parvovirus H/seq4_USA_518-1104nt | MK792432 |
| 5 | Equine Parvovirus H/seq5_USA_518-1104nt | MK792433 |
| 6 | Equine Parvovirus H/seq6_IT_518-1104nt | MK792429 |
| 11 | Equine Parvovirus H/seq11_IT_518-1104nt | MK792434 |
| 16 | Equine Parvovirus H/seq16_USA_518-1104nt | MK792435 |
| 17 | Equine Parvovirus H/seq17_NZ_518-1104nt | MK792436 |
| 18 | Equine Parvovirus H/seq18_CN_518-1104nt | MK792437 |
| 1 | Equine Parvovirus H/seq1_NZ_20-535nt | MK792438 |
| 3 | Equine Parvovirus H/seq3_USA_20-535nt | MK792439 |
| 4 | Equine Parvovirus H/seq4_USA_20-535nt | MK792440 |
| 11 | Equine Parvovirus H/seq11_IT_20-535nt | MK792441 |
| 16 | Equine Parvovirus H/seq16_USA_20-535nt | MK792442 |
| 17 | Equine Parvovirus H/seq17_NZ_20-535nt | MK792443 |
| 18 | Equine Parvovirus H/seq18_CN_20-535nt | MK792444 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meister, T.L.; Tegtmeyer, B.; Postel, A.; Cavalleri, J.-M.V.; Todt, D.; Stang, A.; Steinmann, E. Equine Parvovirus-Hepatitis Frequently Detectable in Commercial Equine Serum Pools. Viruses 2019, 11, 461. https://doi.org/10.3390/v11050461
Meister TL, Tegtmeyer B, Postel A, Cavalleri J-MV, Todt D, Stang A, Steinmann E. Equine Parvovirus-Hepatitis Frequently Detectable in Commercial Equine Serum Pools. Viruses. 2019; 11(5):461. https://doi.org/10.3390/v11050461
Chicago/Turabian StyleMeister, Toni Luise, Birthe Tegtmeyer, Alexander Postel, Jessika-M.V. Cavalleri, Daniel Todt, Alexander Stang, and Eike Steinmann. 2019. "Equine Parvovirus-Hepatitis Frequently Detectable in Commercial Equine Serum Pools" Viruses 11, no. 5: 461. https://doi.org/10.3390/v11050461
APA StyleMeister, T. L., Tegtmeyer, B., Postel, A., Cavalleri, J.-M. V., Todt, D., Stang, A., & Steinmann, E. (2019). Equine Parvovirus-Hepatitis Frequently Detectable in Commercial Equine Serum Pools. Viruses, 11(5), 461. https://doi.org/10.3390/v11050461