Modelling Ranavirus Transmission in Populations of Common Frogs (Rana temporaria) in the United Kingdom

 ,
,  ,
,
Abstract
:1. Introduction
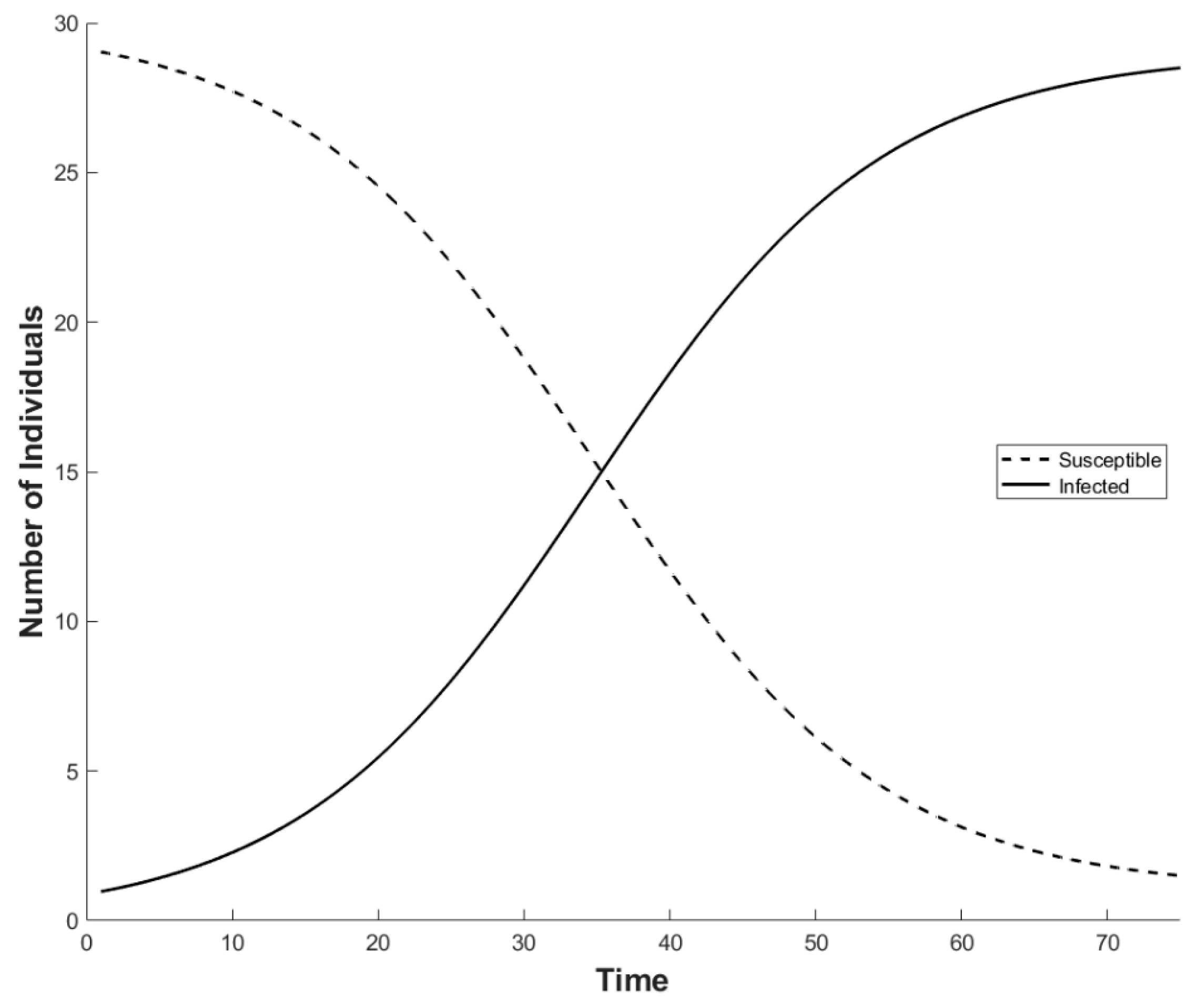
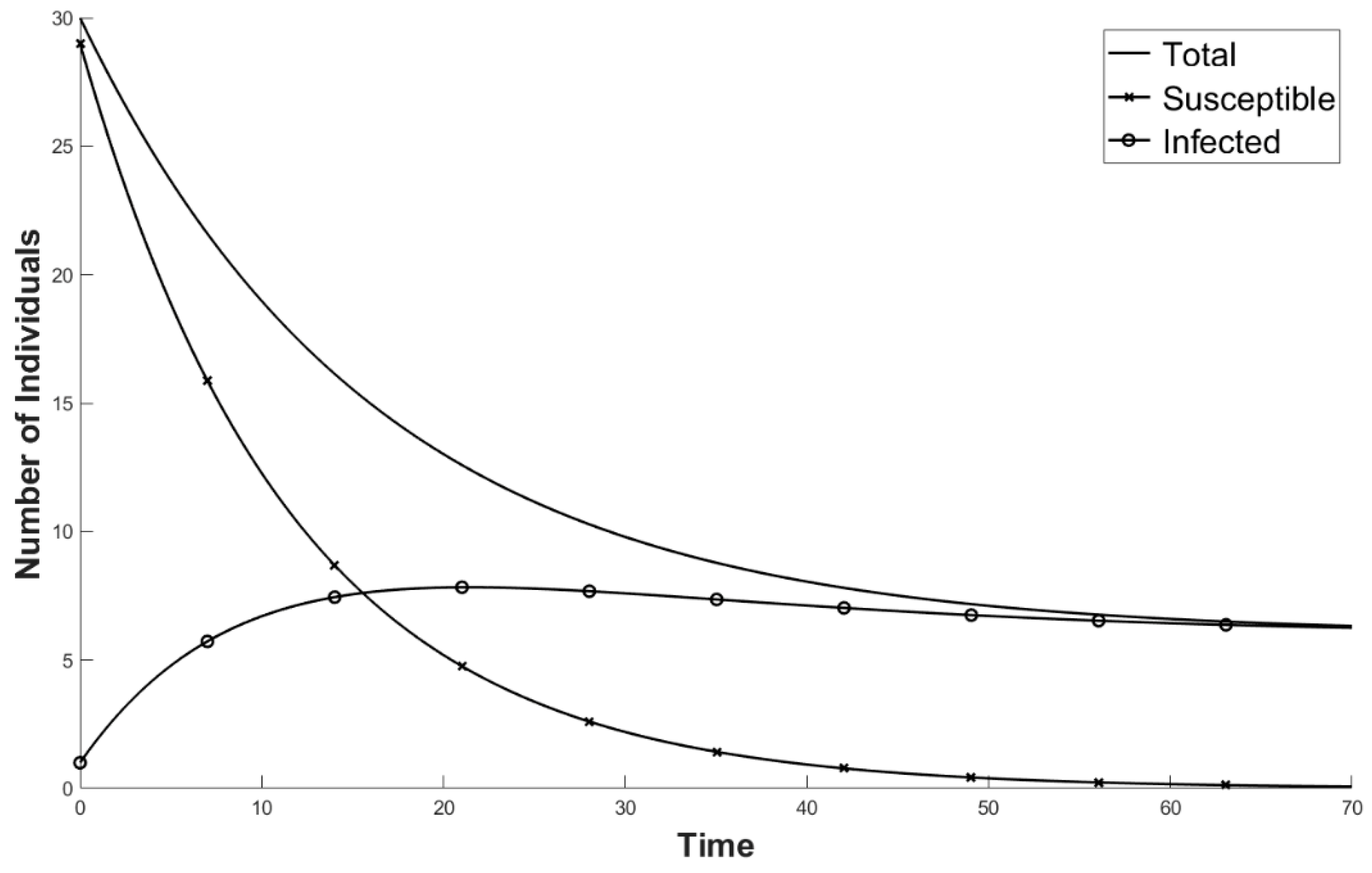
2. Basic Model Formulation
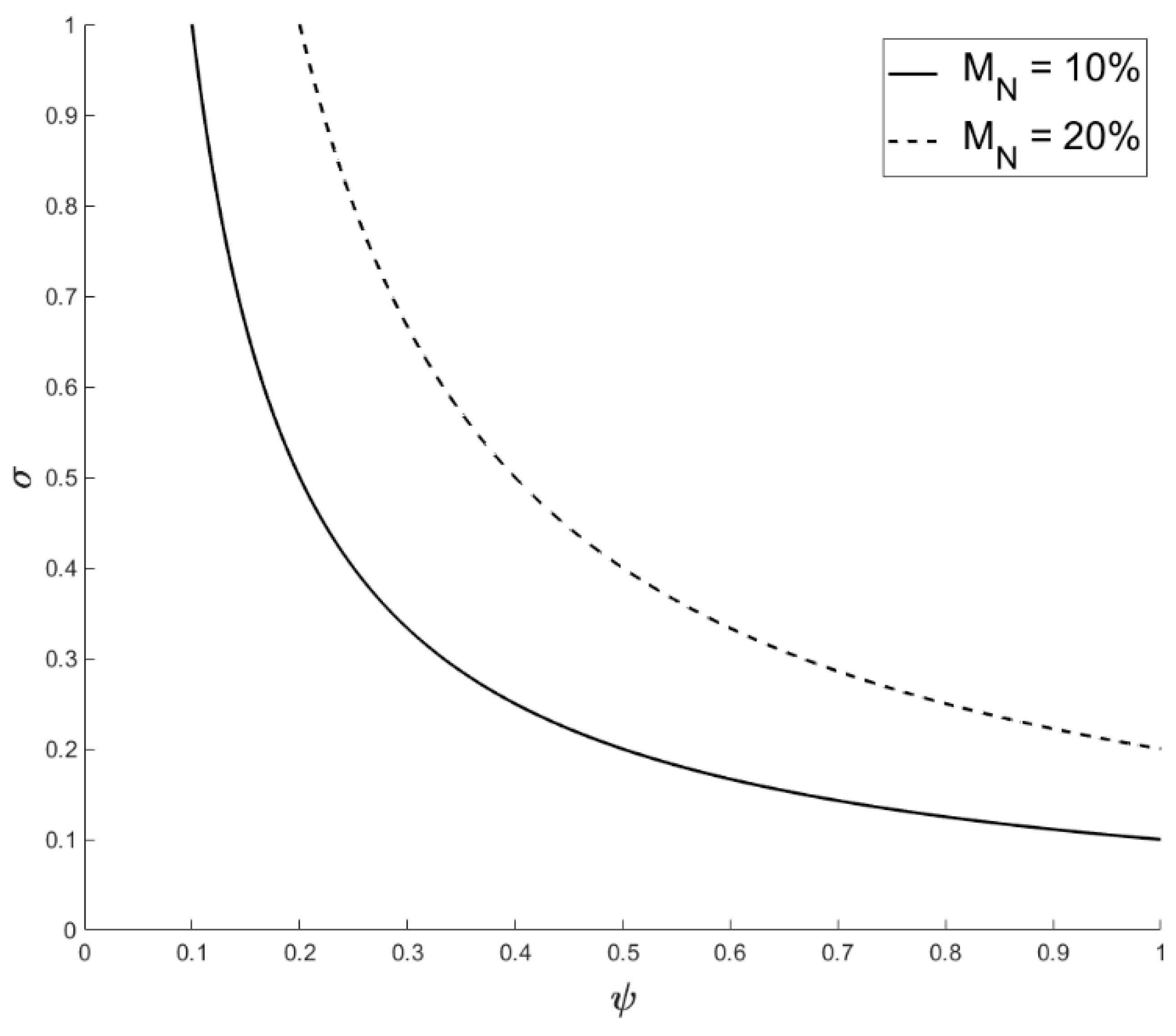
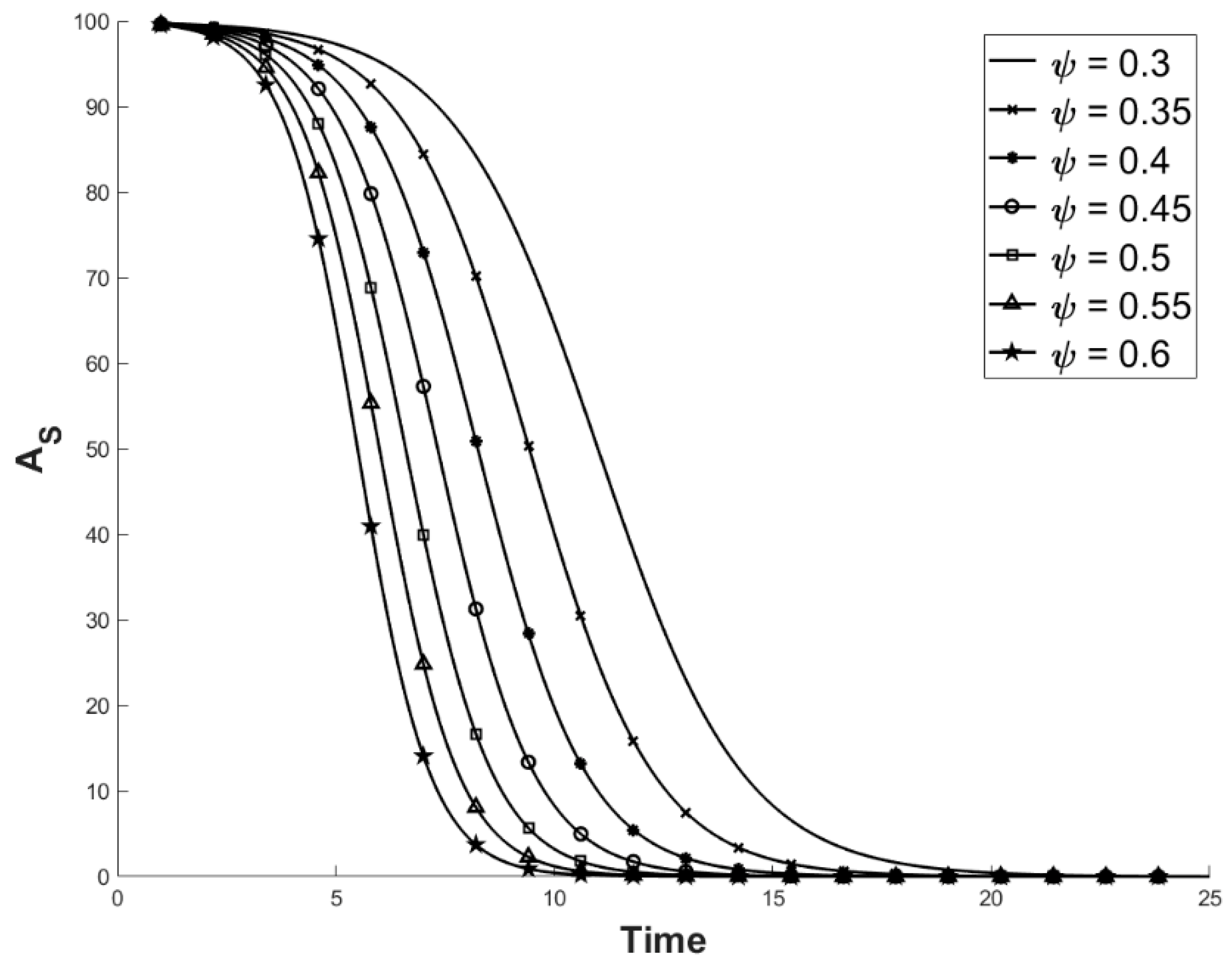
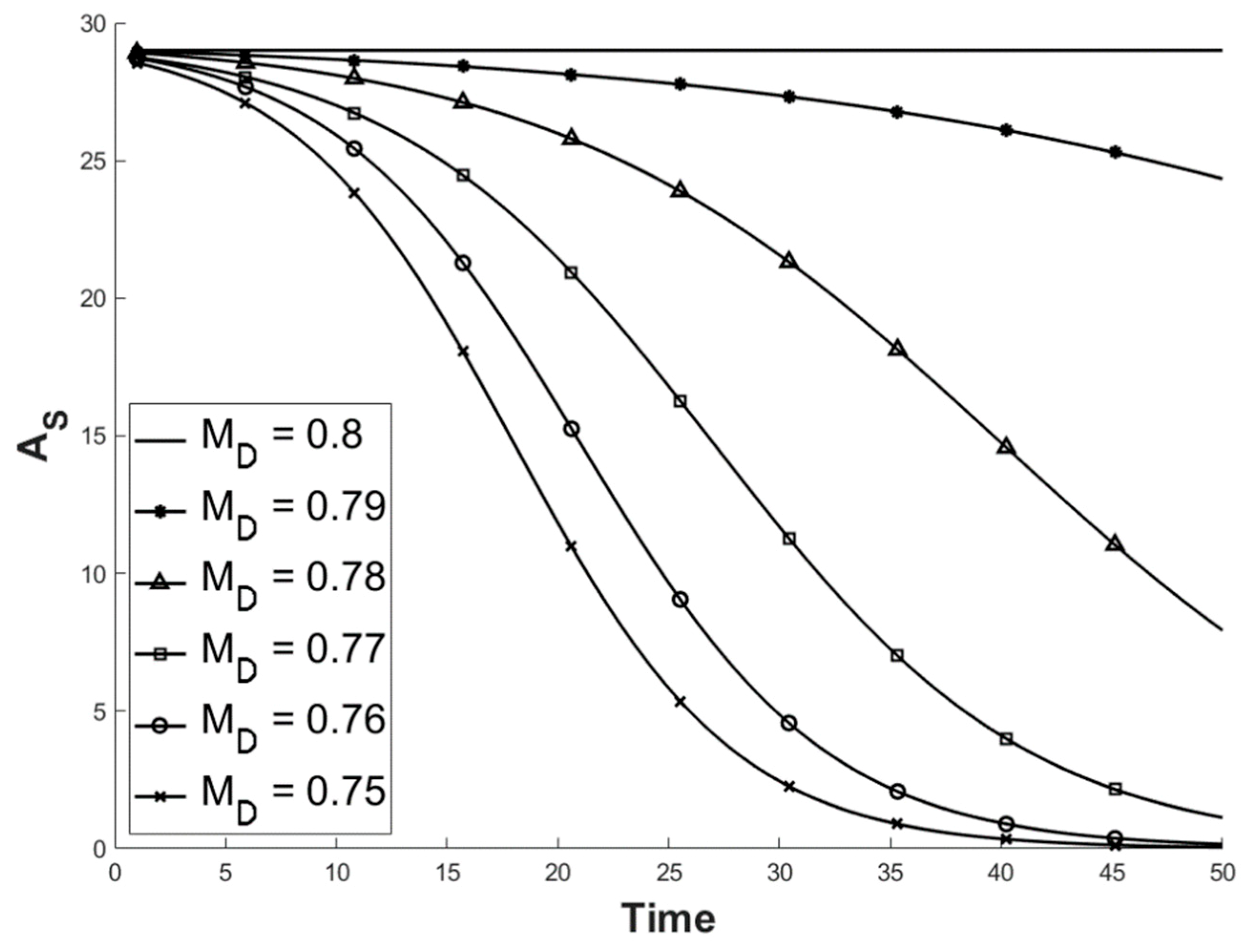
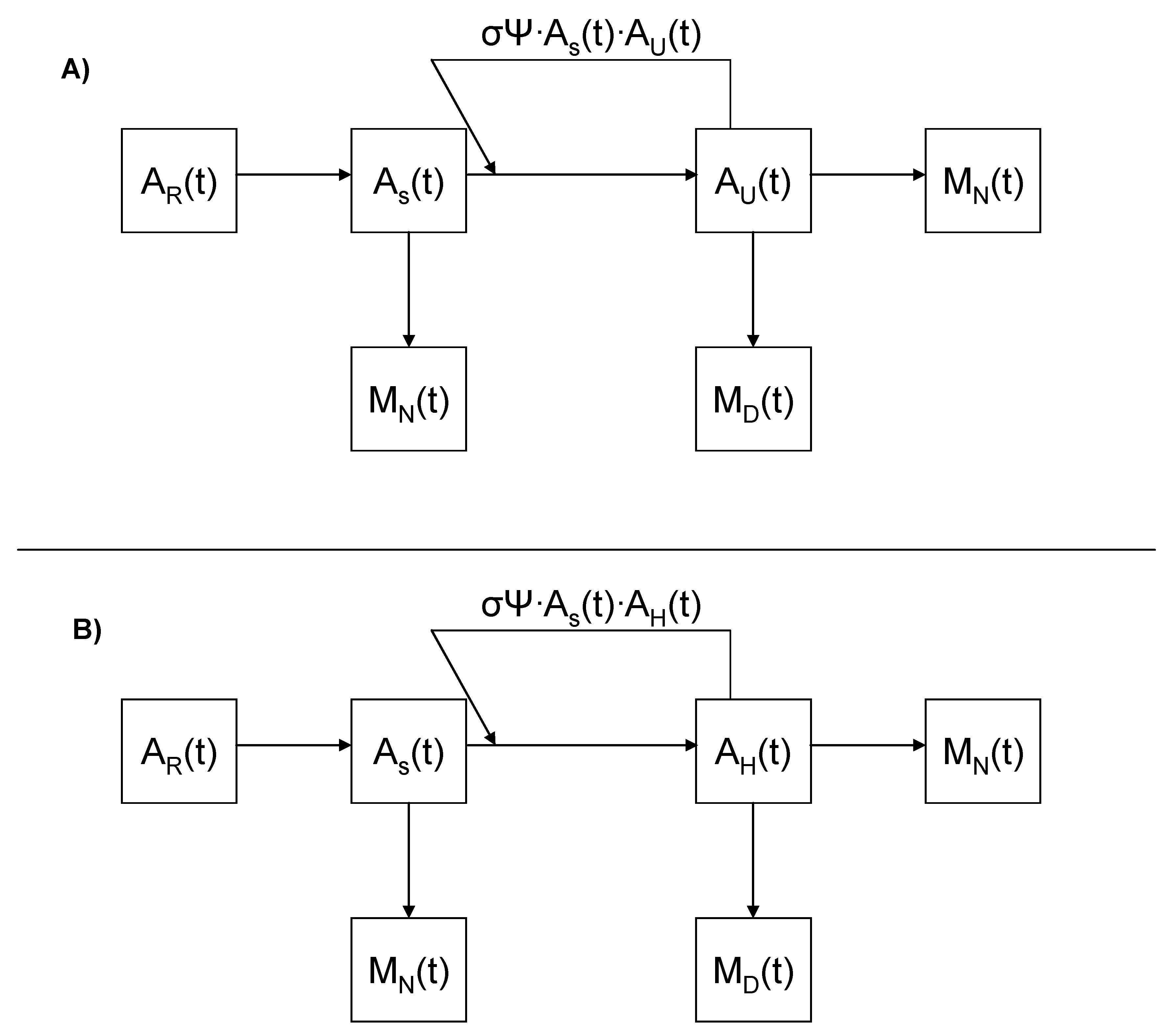
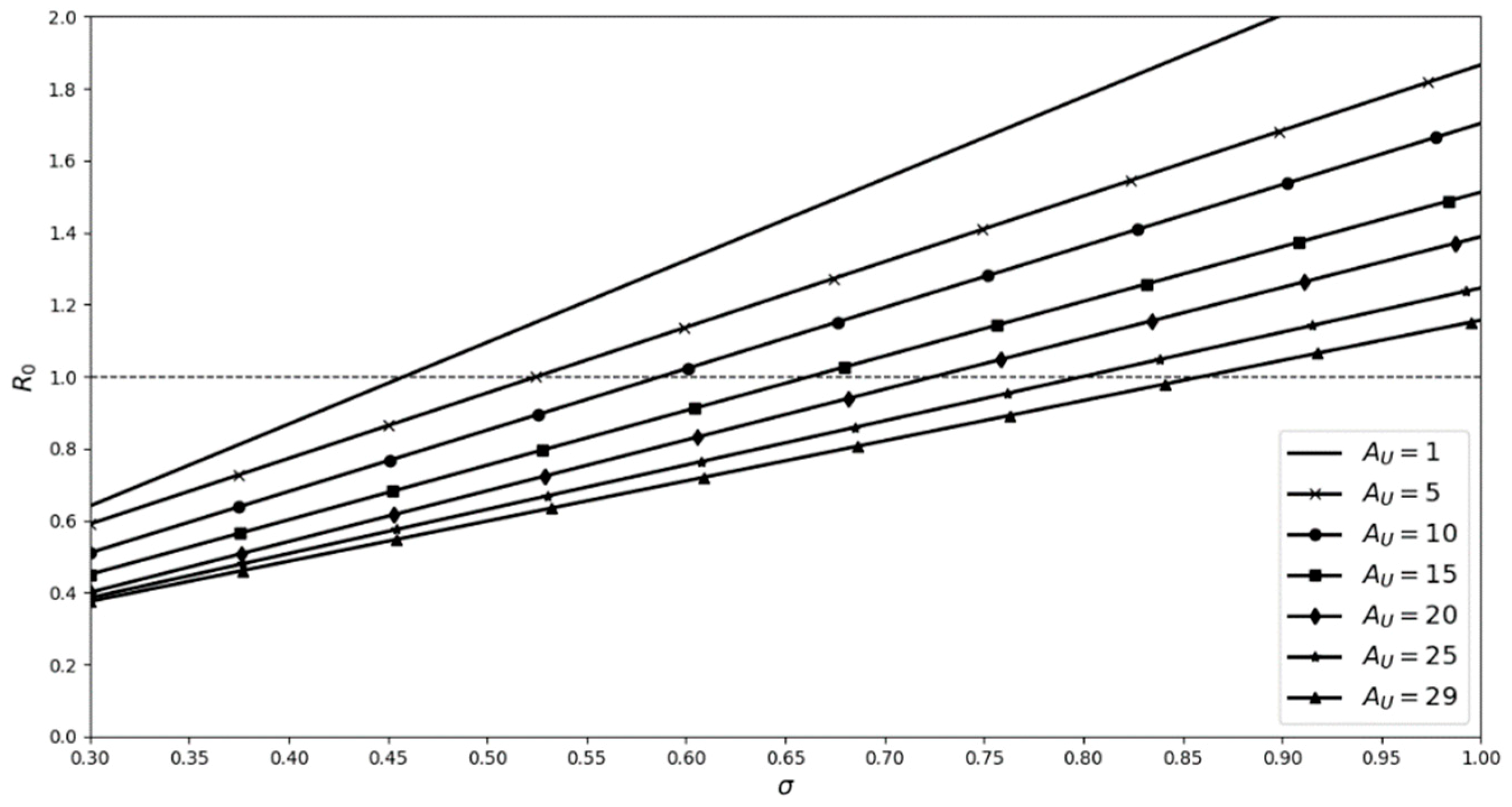
2.1. Factoring in Population Decline
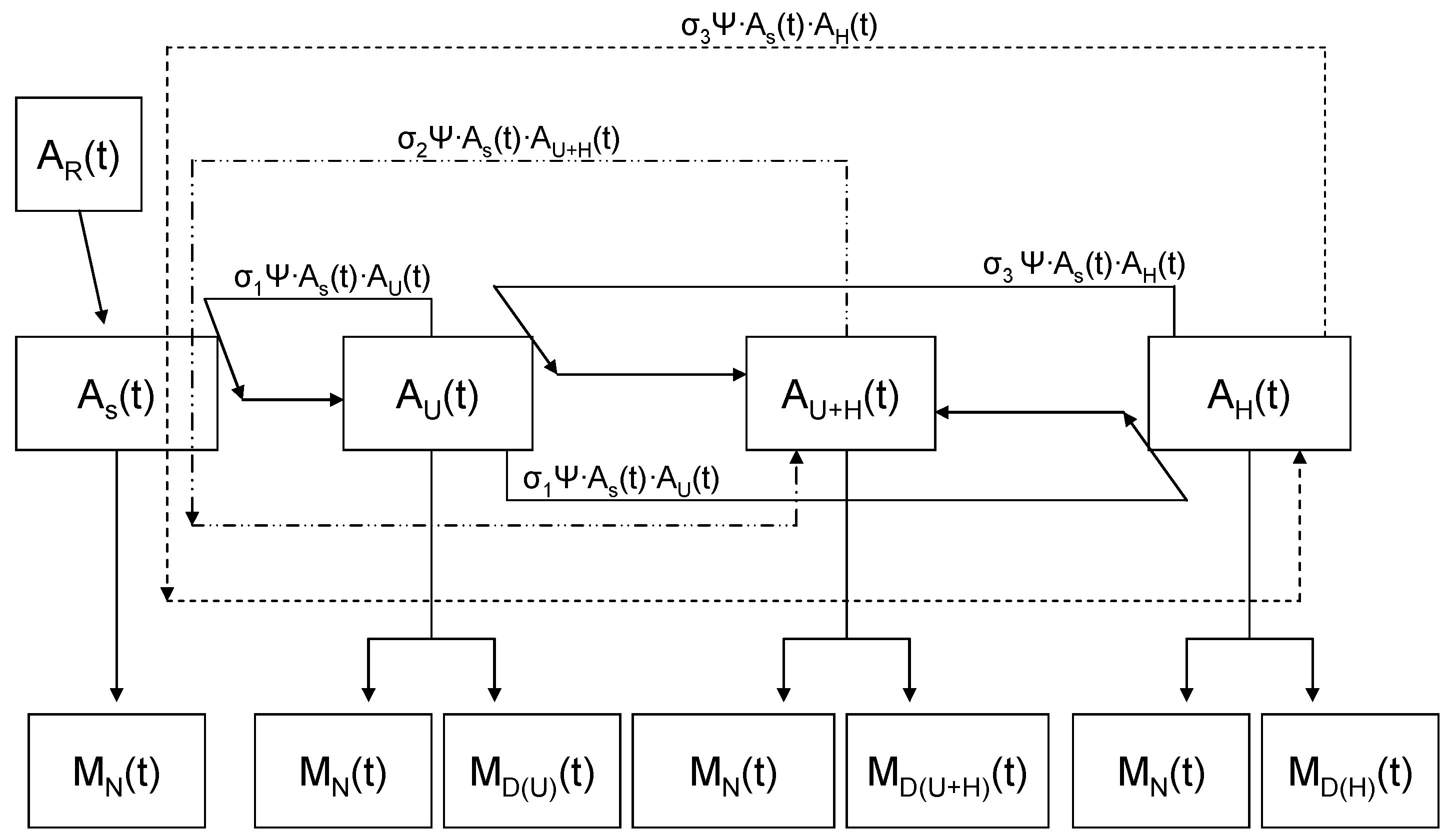
2.2. Accounting for Different Disease Syndromes
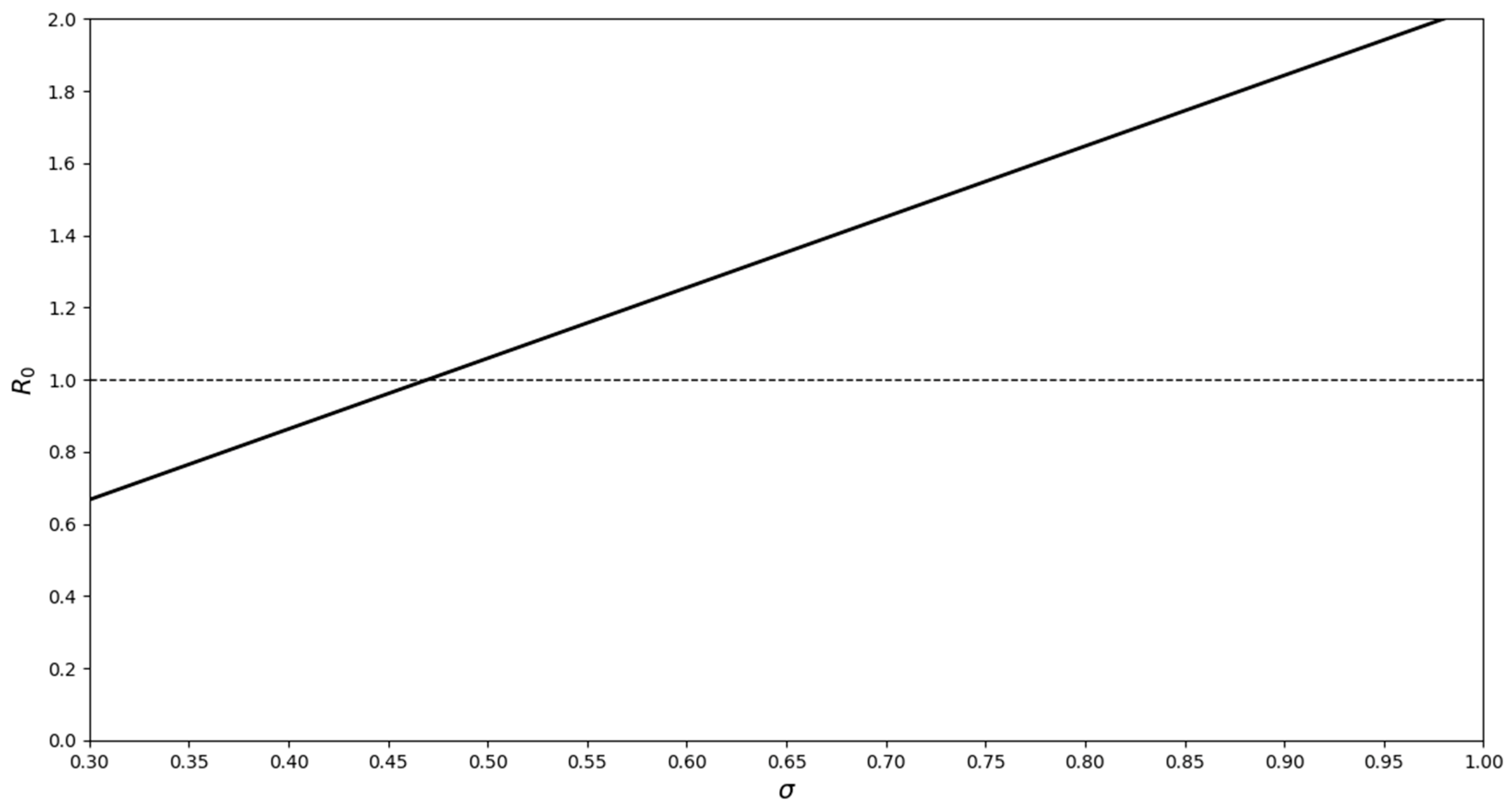
3. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gray, M.J.; Chinchar, V.G. Introduction: History and future of ranaviruses. In Ranaviruses: Lethal Pathogens of Ectothermic Vertebrates; Gray, M.J., Chinchar, V.G., Eds.; Springer: New York, NY, USA, 2015; pp. 1–7. [Google Scholar]
- Duffus, A.L.J.; Waltzek, T.B.; Stöhr, A.C.; Allender, M.C.; Gotesman, M.; Whittington, R.J.; Hick, P.; Hines, M.K.; Marschang, R.E. Distribution and host range of ranviruses. In Ranaviruses: Lethal Pathogens of Ectothermic Vertebrates; Gray, M.J., Chinchar, V.G., Eds.; Springer: Secaucus, NJ, USA, 2015. [Google Scholar]
- Miller, D.L.; Pessier, A.P.; Hick, P.; Whittington, R.J. Comparative pathology of ranaviruses and diagnostic techniques. In Ranaviruses: Lethal Pathogens of Ectothermic Vertebrates; Gray, M.J., Chinchar, V.G., Eds.; Springer: Secaucus, NJ, USA, 2015. [Google Scholar]
- Hoverman, J.T.; Gray, M.J.; Haislip, N.A.; Miller, D.L. Phylogeny, life history, and ecology contribute to differences in amphibian susceptibility to ranaviruses. EcoHealth 2011, 8, 301–319. [Google Scholar] [CrossRef] [PubMed]
- North, A.C.; Hodgson, D.J.; Price, S.J.; Griffiths, A.G.F. Anthropogenic and ecological drivers of amphibian disease (ranavirosis). PLoS ONE 2015, 10, e0127037. [Google Scholar] [CrossRef] [PubMed]
- Balseiro, A.; Dalton, K.P.; del Cerro, A.; Marquez, I.; Cunningham, A.A.; Parra, F.; Prieto, J.M.; Casais, R. Pathology, isolation and molecular characterisation of a ranavirus from the common midwife toad Alytes obstetricans on the iberian peninsula. Dis. Aquat. Org. 2009, 84, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Green, D.E.; Converse, K.A.; Schrader, A.K. Epizootiology of sixty-four amphibian morbidity and mortality events in the USA, 1996–2001. Ann. N. Y. Acad. Sci. 2002, 969, 323–339. [Google Scholar] [CrossRef] [PubMed]
- Muths, E.; Gallant, A.L.; Grant, E.H.C.; Battaglin, W.A.; Green, D.E.; Staiger, J.S.; Walls, S.C.; Gunzburger, M.S.; Kearney, R.F. The Amphibian Research and Monitoring Iniative (ARMI): 5-Year Report; US Geological Survey Scientific Investigations Report 2006-5224; USDOI, USGS, Eds.; US Geological Survey: Reston, VA, USA, 2006; p. 77.
- Teacher, A.G.F.; Cunningham, A.A.; Garner, T.W.J. Assessing the long-term impact of ranavirus infection in wild common frog populations. Anim. Conserv. 2010, 13, 514–522. [Google Scholar] [CrossRef]
- Price, S.J.; Garner, T.W.J.; Nichols, R.A.; Balloux, F.; Ayres, C.; Mora-Cabello de Alba, A.; Bosch, J. Collapse of amphibian communities due to an introduced ranavirus. Curr. Biol. 2014, 24, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Earl, J.E.; Gray, M.J. Introduction of ranavirus to isolated wood frog populations could cause local extinction. EcoHealth 2014, 11, 581–592. [Google Scholar] [CrossRef]
- Earl, J.E.; Chaney, J.C.; Sutton, W.B.; Lillard, C.E.; Kouba, A.J.; Langhorne, C.; Krebs, J.; Wilkes, R.P.; Hill, R.D.; Miller, D.L.; et al. Ranavirus could facilitate local extinction of rare amphibian species. Oecologia 2016, 182, 611–623. [Google Scholar] [CrossRef]
- Farrell, A.P.; Collins, J.P.; Greer, A.L.; Thieme, H.R. Times from infection to disease-induced death and their influence on final population sizes after epidemic outbreaks. Bull. Math. Biol. 2018, 80, 1937–1961. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Population biology of infectious disease. Nature 1979, 280, 361–461. [Google Scholar] [CrossRef]
- May, R.M.; Anderson, R.M. Population biology of infectious diseases: Part II. Nature 1979, 280, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Brunner, J.L.; Yarber, C.M. Evaluating the importance of environmental persistence for Ranavirus transmission and epidemiology. Adv. Virus Res. 2018, 101, 129–148. [Google Scholar] [PubMed]
- Brunner, J.L.; Schock, D.M.; Davidson, E.W.; Collins, J.P. Intraspecific reservoirs: Complex life history and the persistence of a lethal ranavirus. Ecology 2004, 85, 560–566. [Google Scholar] [CrossRef]
- Duffus, A.L.J.; Pauli, B.D.; Wozney, K.; Brunetti, C.R.; Berrill, M. Frog virus 3-like infections in aquatic amphibian communities. J. Wildl. Dis. 2008, 44, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Duffus, A.L.J.; Nichols, R.A.; Garner, T.W.J. Experimental evidence in support of single host maintenance of a multihost pathogen. Ecosphere 2014, 5, 1–11. [Google Scholar] [CrossRef]
- Duffus, A.L.J.; Nichols, R.A.; Garner, T.W.J. Investigations into the life history stages of the common frog (Rana temporaria) affected by an amphibian ranavirus in the United Kingdom. Herpetol. Rev. 2013, 44, 260–263. [Google Scholar]
- Cunningham, A.A.; Langton, T.E.S.; Bennett, P.M.; Lewin, J.F.; Drury, S.E.N.; Gough, R.E.; MacGregor, S.K. Pathological and microbiological findings from incidents of unusual mortality of the common frog (Rana temporaria). Philos. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 1539–1557. [Google Scholar]
- Otto, S.P.; Day, T. A Biologist’s Guide to Mathematical Modeling in Ecology and Evolution; Princeton University Press: Princeton, NJ, USA, 2007; p. 732. [Google Scholar]
- Miaud, C.; Guyetant, R.; Elmberg, J. Variations in life-history traits in the common frog Rana temporaria (Amphibia: Anura): A literature review and new data from the French Alps. J. Zool. 1999, 249, 61–73. [Google Scholar] [CrossRef]
- Gibbons, M.M.; McCarthy, T.K. Growth, maturation and survival of frogs Rana temporaria L. Holarct. Ecol. 1984, 7, 419–427. [Google Scholar] [CrossRef]
- Cunningham, A.A.; Hyatt, A.D.; Russell, P.; Bennett, P.M. Emerging epidemic diseases of frogs in britain are dependant on the source of ranavirus agent and the route of exposure. Epidemiol. Infect. 2007, 135, 1200–1212. [Google Scholar] [CrossRef] [PubMed]
- Vieites, D.R.; Nieto-Román, S.; Barluenga, M.; Palanca, A.; Vences, M.; Meyer, A. Post-mating clutch piracy in an amphibian. Nature 2004, 431, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Price, S.J.; Wadia, A.; Wright, O.N.; Leung, W.T.; Cunningham, A.A.; Lawson, B. Screening of a long-term sample set reveals two ranavirus lineages in british herpetofauna. PLoS ONE 2017, 12, e0184768. [Google Scholar] [CrossRef] [PubMed]
- Price, S.J.; Leung, W.T.M.; Owen, C.; Puschendorf, R.; Sergeant, C.; Cunningham, A.A.; Balloux, F.; Garner, T.W.J.; Nichols, R.A. Effects of historic and projected climate change on the range and impacts of an emerging wildlife disease. Glob. Chang. Biol. 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- Teacher, A.G.F.; Garner, T.W.J.; Nichols, R.A. Evidence for directional selection at a novel major histocompatability class i marker in wild common frogs (Rana temporaria) exposed to a viral pathogen (Ranavirus). PLoS ONE 2009, 4, e4616. [Google Scholar] [CrossRef]
- Campbell, L.J.; Garner, T.W.J.; Tessa, G.; Scheele, B.C.; Griffiths, A.G.F.; Harrison, X.A. An emerging viral pathogen truncates population age structure in a european amphibian and may reduce population viability. PeerJ 2018, 6, e5949. [Google Scholar] [CrossRef] [PubMed]
- Kik, M.; Martel, A.; Spitzen-van der Sluijs, A.; Pasmans, F.; Wohlsein, P.; Gröne, A.; Rijks, J.M. Ranavirus-associated mass mortality in wild amphibians, the netherlands, 2010: A first report. Vet. J. 2011, 190, 284–286. [Google Scholar] [CrossRef]
- Duffus, A.L.J.; Nichols, R.A.; Garner, T.W.J. Detection of a frog virus 3-like ranavirus in native and introduced amphibians in the United Kingdom in 2007 and 2008. Herpetol. Rev. 2014, 45, 608–610. [Google Scholar]
- Mihaljevic, J.R.; Hoverman, J.T.; Johnson, P.T.J. Co-exposure to multiple ranavirus types enhances viral infectivity and replication in a larval amphibian system. Dis. Aquat. Org. 2018, 132, 23–35. [Google Scholar] [CrossRef]
- Duffus, A.L.J.; Garner, T.W.J.; Davis, A.R.; Dean, A.W.; Nichols, R.A. Phylogentic analysis of 24 ranavirus isolates from English amphibians using 2 partial loci. J. Emerg. Dis. Virol. 2017, 3. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Development of Disease | Disease Prevalence | Type of Experiment/Exposure Type | Estimate of σ | TCID50 | |
|---|---|---|---|---|---|
| No. with Disease | Total No. Exposed | ||||
| 3 | 20 | 15% | Immersion with virus from naturally disease tissue, with and without bacteria | 0.15 | 101/mL |
| 9 | 20 | 45% | Immersion with virus from naturally disease tissue homogenate to animals with skin wounds, with and without bacteria | 0.45 | 102/mL HS 101.5/mL US |
| 9 | 10 | 90% | Immersion in virus from culture | 0.90 | 102/mL HS 101.5/mL US |
| 5 | 5 | 100% | Immersion in virus from virus culture to animals with wounded skin | 1 | 105.6 to 106.2/mL |
| 2 | 5 | 40% | Immersion in virus from tissue homogenate from naturally diseased animals to animals with wounded skin | 0.40 | 103/mL |
| Development of Disease | Type of Experiment/Exposure Type | Estimate of σ | Average Estimate of σ | |||
|---|---|---|---|---|---|---|
| No. with U | No. with H | No. with U & H | Total Exp. | |||
| 2 | 0 | 0 | 5 | Immersion with virus from naturally disease tissue with bacteria (Ulcerative) | 0.4 | 0.36 |
| 1 | 0 | 0 | 5 | Immersion with virus from naturally disease tissue without bacteria (Ulcerative) | 0.2 | |
| 2 | 0 | 0 | 5 | Immersion with virus from naturally disease tissue to animals with skin wounds with bacteria (Ulcerative) | 0.4 | |
| 0 | 0 | 0 | 5 | Immersion with virus from naturally disease tissue to animals with skin wounds without bacteria (Ulcerative) | 0 | |
| 2 | 2 | 0 | 5 | Immersion in virus isolated from naturally diseased animals from virus culture (RUK 13, Ulcerative) | 0.8 | |
| 0 | 0 | 0 | 5 | Immersion with virus from naturally disease tissue without bacteria (Hemorrhagic) | 0 | 0.44 |
| 0 | 0 | 0 | 5 | Immersion with virus from naturally disease tissue with bacteria (Hemorrhagic) | 0 | |
| 1 | 1 | 1 | 5 | Immersion with virus from naturally disease tissue to animals with skin wounds with bacteria (Hemorrhagic) | 0.6 | |
| 0 | 3 | 1 | 5 | Immersion with virus from naturally disease tissue to animals with skin wounds without bacteria (Hemorrhagic) | 0.8 | |
| 1 | 2 | 1 | 5 | Immersion in virus isolated from naturally diseased animals from virus culture (RUK 11, Hemorrhagic) | 0.8 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duffus, A.L.J.; Garner, T.W.J.; Nichols, R.A.; Standridge, J.P.; Earl, J.E. Modelling Ranavirus Transmission in Populations of Common Frogs (Rana temporaria) in the United Kingdom. Viruses 2019, 11, 556. https://doi.org/10.3390/v11060556
Duffus ALJ, Garner TWJ, Nichols RA, Standridge JP, Earl JE. Modelling Ranavirus Transmission in Populations of Common Frogs (Rana temporaria) in the United Kingdom. Viruses. 2019; 11(6):556. https://doi.org/10.3390/v11060556
Chicago/Turabian StyleDuffus, Amanda L.J., Trenton W.J. Garner, Richard A. Nichols, Joshua P. Standridge, and Julia E. Earl. 2019. "Modelling Ranavirus Transmission in Populations of Common Frogs (Rana temporaria) in the United Kingdom" Viruses 11, no. 6: 556. https://doi.org/10.3390/v11060556
APA StyleDuffus, A. L. J., Garner, T. W. J., Nichols, R. A., Standridge, J. P., & Earl, J. E. (2019). Modelling Ranavirus Transmission in Populations of Common Frogs (Rana temporaria) in the United Kingdom. Viruses, 11(6), 556. https://doi.org/10.3390/v11060556