Dendritic Cell Targeting of Bovine Viral Diarrhea Virus E2 Protein Expressed by Lactobacillus casei Effectively Induces Antigen-Specific Immune Responses via Oral Vaccination

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Virus, Plasmid, and Animals
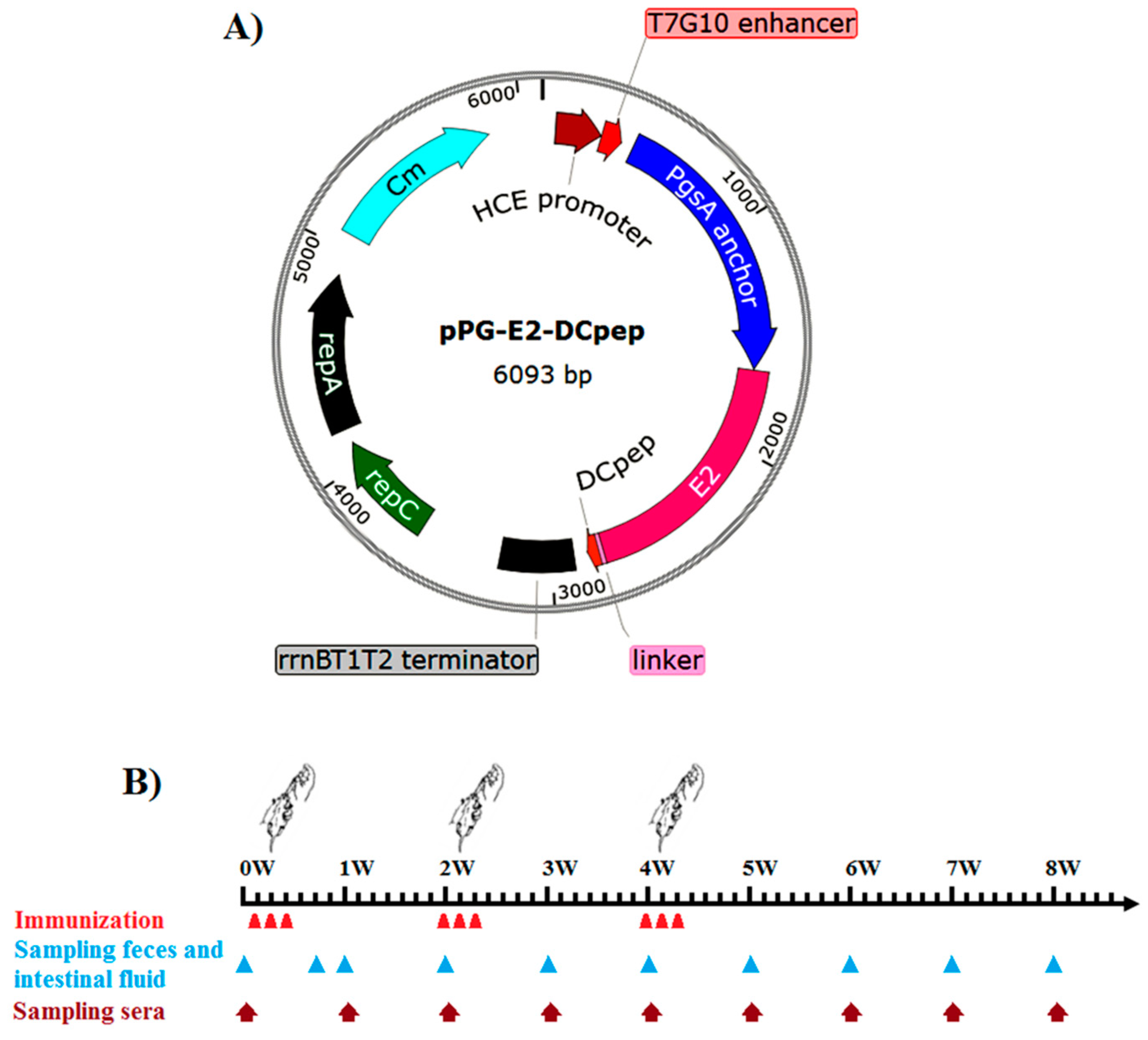
2.2. Construction of Recombinant Lactobacillus
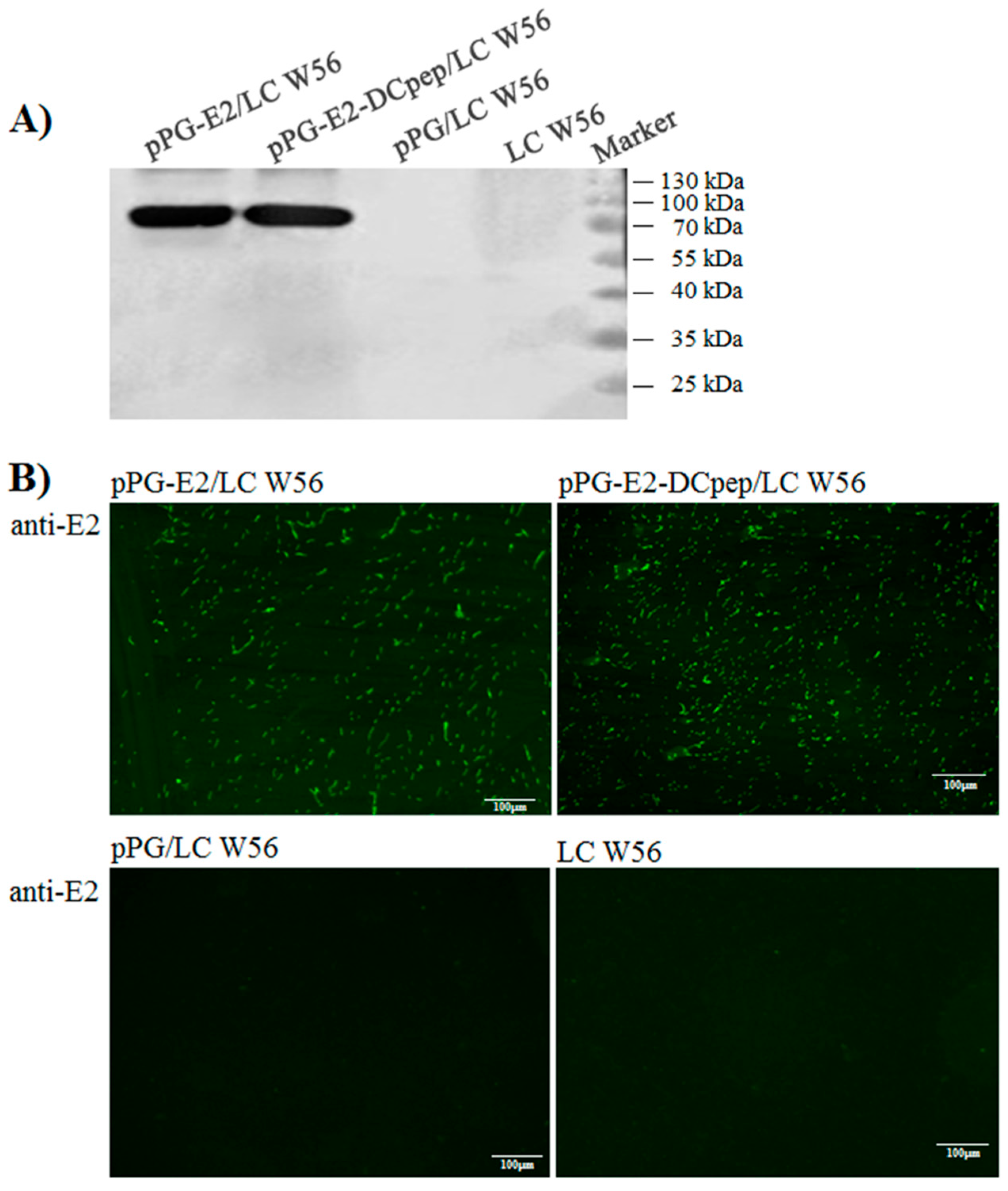
2.3. Expression and Identification of the Fusion Protein
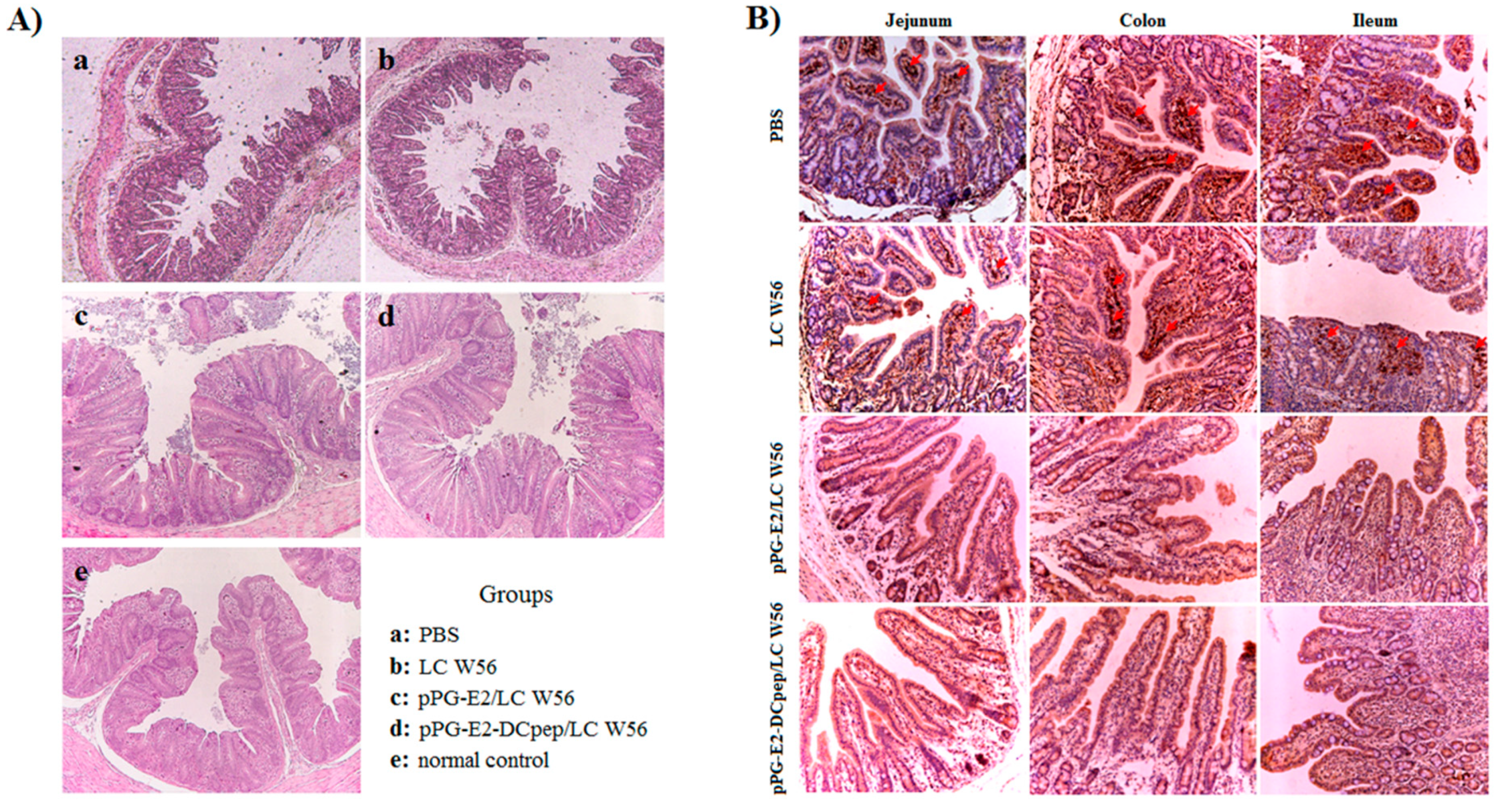
2.4. Determination of Adherence Ability of Recombinant Lactobacillus in Intestinal Tracts
2.5. Oral Immunization and Immune Sample Collection
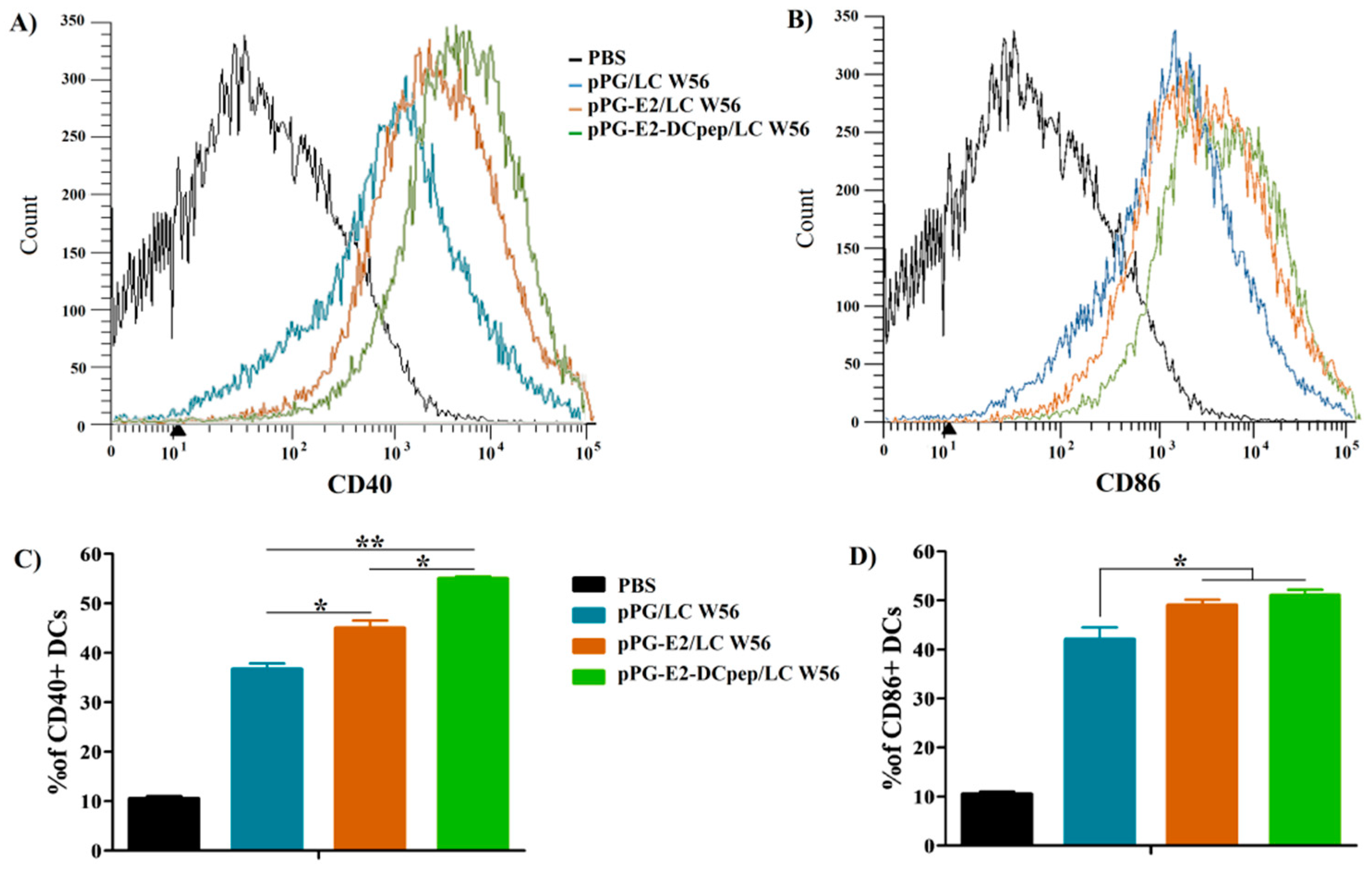
2.6. Activation of Dendritic Cells in Intestinal Peyer’s Patches by Recombinant Lactobacillus
2.7. ELISA of Antibody Levels Induced by Rrecombinant Lactobacillus
2.8. Determination of Neutralization Ability of Antibodies
2.9. Measurement of Lymphocyte Proliferation and Cytokine Levels
2.10. Challenge Experiment
2.11. Statistical Analysis
3. Results
3.1. Identification of the Protein of Interest Expressed by the Recombinant Lactobacilli
3.2. Colonization Ability of the Recombinant Lactobacillus in the Intestinal Tract
3.3. Activation of DCs in PPs Stimulated by the Recombinant Lactobacillus
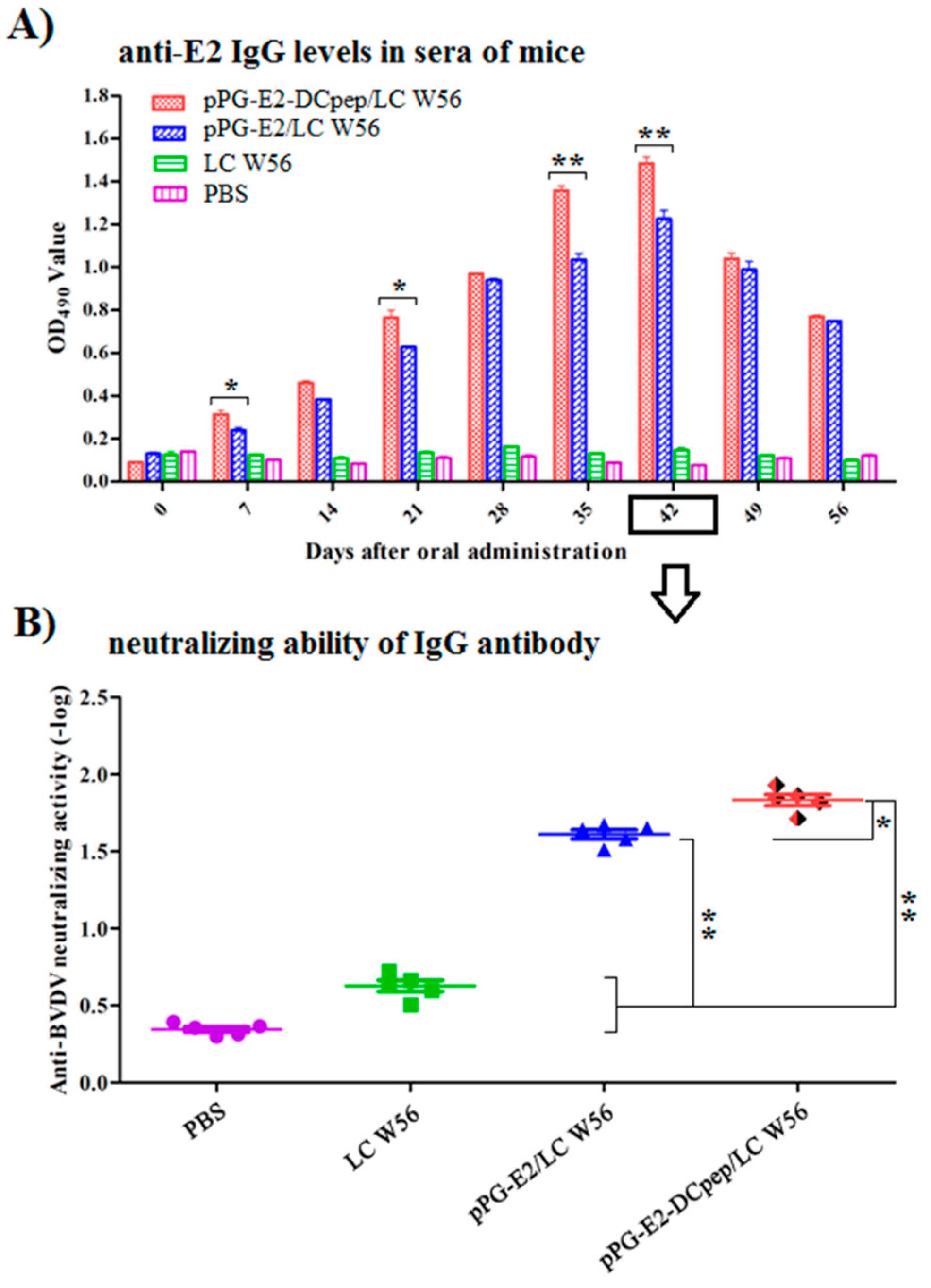
3.4. Antigen-Specific Serum IgG Antibody Levels Induced by Recombinant Lactobacillus
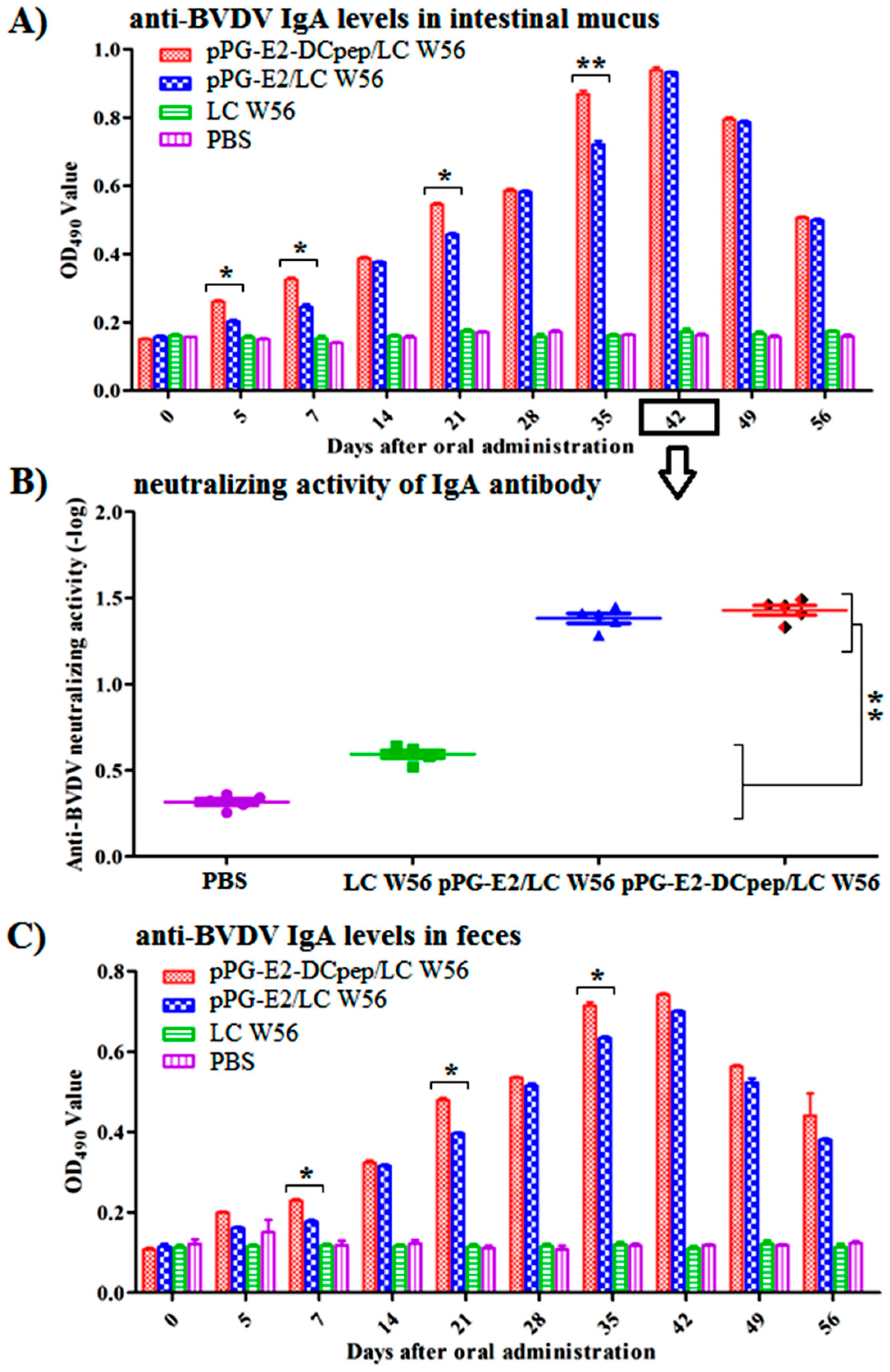
3.5. Antigen-Specific Intestinal Mucosal sIgA Antibody Levels Induced by Recombinant Lactobacillus
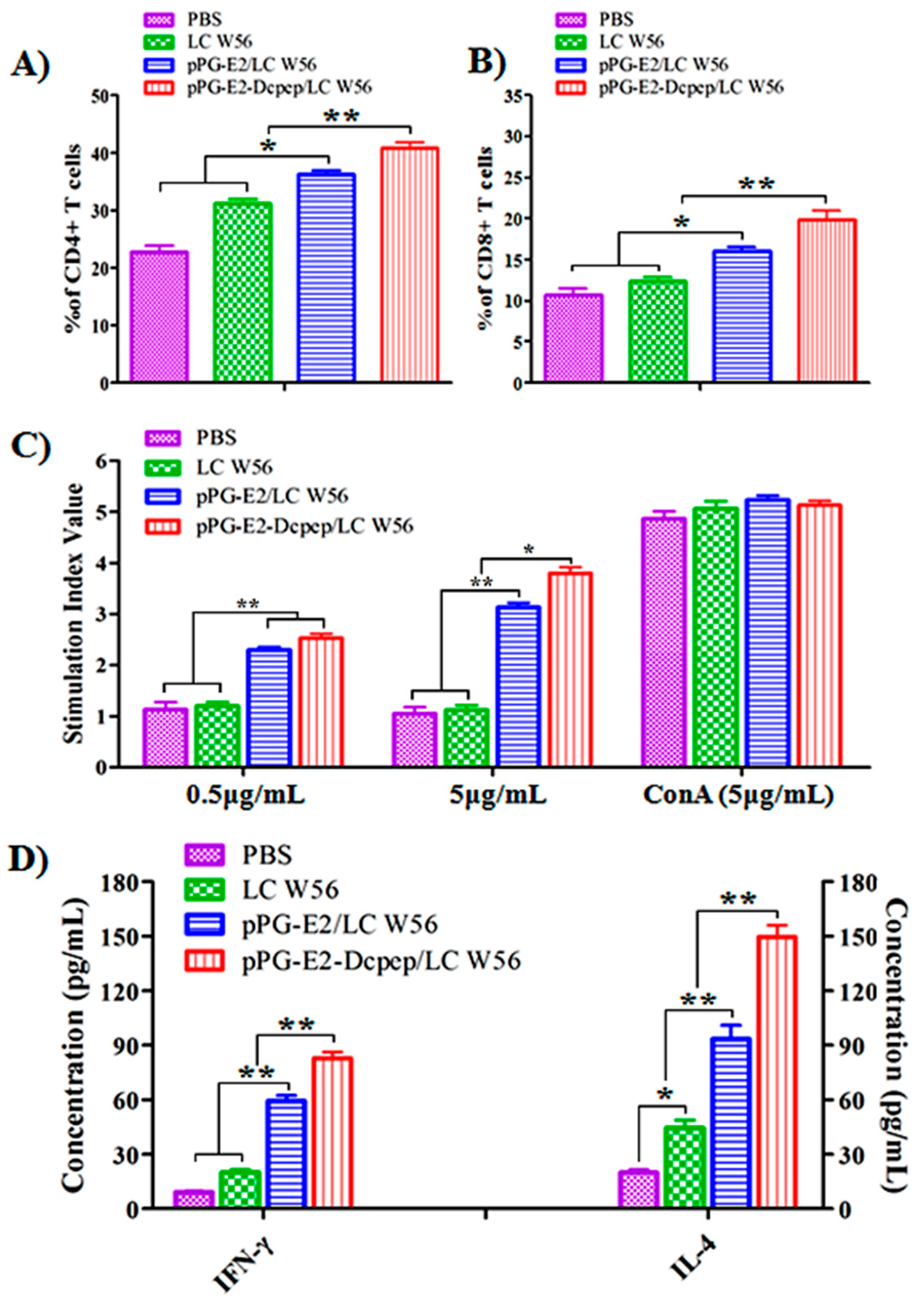
3.6. CD4+ and CD8+ T Cell Detection
3.7. Lymphocyte Proliferation
3.8. Challenge Experiment Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fray, M.D.; Paton, D.J.; Alenius, S. The effects of bovine viral diarrhoea virus on cattle reproduction in relation to disease control. Anim. Reprod. Sci. 2000, 60, 615–627. [Google Scholar] [CrossRef]
- Sarrazin, S.; Veldhuis, A.; Méroc, E.; Vangeel, I.; Laureyns, J.; Dewulf, J.; Caij, A.B.; Piepers, S.; Hooyberghs, J.; Ribbens, S.; et al. Serological and virological BVDV prevalence and risk factor analysis for herds to be BVDV seropositive in Belgian cattle herds. Prev. Vet. Med. 2013, 108, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Szabára, Á.; Lang, Z.; Földi, J.; Hornyák, Á.; Abonyi, T.; Ózsvári, L. Prevalence of bovine viral diarrhoea virus in cattle farms in hungary. Acta. Vet. Hung. 2016, 64, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.G.; Cong, W.; Zhang, F.H.; Feng, S.Y.; Zhou, D.H.; Wang, Y.M.; Zhu, X.Q.; Yin, H.; Hu, G.X. Seroprevalence and risk factors of bovine viral diarrhoea virus (BVDV) infection in yaks (Bos grunniens) in northwest China. Trop. Anim. Health Prod. 2016, 48, 1747–1750. [Google Scholar] [CrossRef] [PubMed]
- Scharnböck, B.; Roch, F.F.; Richter, V.; Funke, C.; Firth, C.; Obritzhauser, W.; Baumgartner, W.; Käsbohrer, A.; Pinior, B. A meta-analysis of bovine viral diarrhoea virus (BVDV) prevalences in the global cattle population. Sci. Rep. 2018, 8, 14420. [Google Scholar] [CrossRef] [PubMed]
- Neill, J.D. Molecular biology of bovine viral diarrhea virus. Biologicals 2013, 41, 2–7. [Google Scholar] [CrossRef]
- Ridpath, J.F.; Bolin, S.R.; Dubovi, E.J. Segregation of bovine viral diarrhea virus into genotypes. Virology 1994, 205, 66–74. [Google Scholar] [CrossRef]
- Giammarioli, M.; Ceglie, L.; Rossi, E.; Bazzucchi, M.; Casciari, C.; Petrini, S.; de Mia, G.M. Increased genetic diversity of BVDV-1: Recent findings and implications thereof. Virus Genes 2015, 50, 147–151. [Google Scholar] [CrossRef]
- Decaro, N.; Lucente, M.S.; Lanave, G.; Gargano, P.; Larocca, V.; Losurdo, M.; Ciambrone, L.; Marino, P.A.; Parisi, A.; Casalinuovo, F. Evidence for circulation of bovine viral diarrhoea virus type 2c in ruminants in southern Italy. Transbound. Emerg. Dis. 2017, 64, 1935–1944. [Google Scholar] [CrossRef]
- Brodersen, B.W. Bovine viral diarrhea virus infections: Manifestations of infection and recent advances in understanding pathogenesis and control. Vet. Pathol. 2014, 51, 453–464. [Google Scholar] [CrossRef]
- Ståhl, K.; Alenius, S. BVDV control and eradication in Europe-an update. JPN. J. Vet. Res. 2012, 60, S31–S39. [Google Scholar] [PubMed]
- Lindberg, A.L.; Alenius, S. Principles for eradication of bovine viral diarrhoea virus (BVDV) infections in cattle populations. Vet. Microbiol. 1999, 64, 197–222. [Google Scholar] [CrossRef]
- Houe, H.; Lindberg, A.; Moennig, V. Test strategies in bovine viral diarrhea virus control and eradication campaigns in Europe. J. Vet. Diagn. Investig. 2006, 18, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, A.; Brownlie, J.; Gunn, G.J.; Houe, H.; Moennig, V.; Saatkamp, H.W.; Sandvik, T.; Valle, P.S. The control of bovine viral diarrhoea virus in Europe: Today and in the future. Rev. Sci. Tech. 2006, 25, 961–979. [Google Scholar] [CrossRef] [PubMed]
- Stott, A.W.; Humphry, R.W.; Gunn, G.J.; Higgins, I.; Hennessy, T.; O’Flaherty, J.; Graham, D.A. Predicted costs and benefits of eradicating BVDV from Ireland. Irish Vet. J. 2012, 65, 12. [Google Scholar] [CrossRef] [PubMed]
- Lanyon, S.; Reichel, M. Bovine viral diarrhoea virus (‘pestivirus’) in Australia: To control or not to control? Aust. Vet. J. 2014, 92, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Falcone, E.; Cordioli, P.; Tarantino, M.; Muscillo, M.; Sala, G.; Rosa, G.L.; Archetti, I.L.; Marianelli, C.; Lombardi, G.; Tollis, M. Experimental infection of calves with bovine viral diarrhoea virus type-2 (bvdv-2) isolated from a contaminated vaccine. Vet. Res. Commun. 2003, 27, 577–589. [Google Scholar] [CrossRef]
- Frink, S.; Grummer, B.; Pohlenz, J.F.; Liebler-Tenorio, E.M. Changes in distribution and numbers of CD4+ and CD8+ T-lymphocytes in lymphoid tissues and intestinal mucosa in the early phase of experimentally induced early onset mucosal disease in cattle. J. Vet. Med. B Infect. Dis. Vet. Public Health 2002, 49, 476–483. [Google Scholar] [CrossRef]
- Xu, Y.; Li, Y. Construction of recombinant Lactobacillus casei efficiently surface displayed and secreted porcine parvovirus VP2 protein and comparison of the immune responses induced by oral immunization. Immunology 2008, 124, 68–75. [Google Scholar]
- Xu, Y.G.; Guan, X.T.; Liu, Z.M.; Tian, C.Y.; Cui, L.C. Immunogenicity in swine of orally administered recombinant Lactobacillus plantarum expressing classical swine fever virus E2 protein in conjunction with thymosin α-1 as an adjuvant. Appl. Environ. Microbiol. 2015, 81, 3745–3752. [Google Scholar] [CrossRef]
- Yu, M.; Qi, R.; Chen, C.; Yin, J.; Ma, S.; Shi, W.; Wu, Y.; Ge, J.; Jiang, Y.; Tang, L.; et al. Immunogenicity of recombinant Lactobacillus casei expressing F4 (K88) fimbrial adhesin FaeG in conjunction with a heat-labile enterotoxin A (LTAK 63) and heat-labile enterotoxin B (LTB) of enterotoxigenic Escherichia coli as an oral adjuvant in mice. J. Appl. Microbiol. 2017, 122, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Wang, L.; Ma, S.; Wang, X.; Wang, Y.; Xiao, Y.; Jiang, Y.; Qiao, X.; Tang, L.; Xu, Y.; et al. Immunogenicity of eGFP-marked recombinant Lactobacillus casei against transmissible gastroenteritis virus and porcine epidemic diarrhea virus. Viruses 2017, 9, 274. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, L.; Huang, X.; Ma, S.; Yu, M.; Shi, W.; Qiao, X.; Tang, L.; Xu, Y.; Li, Y. Oral delivery of probiotics expressing dendritic cell-targeting peptide fused with porcine epidemic diarrhea virus COE antigen: A promising vaccine strategy against PEDV. Viruses 2017, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Wang, L.; Huang, X.; Wang, X.; Chen, S.; Shi, W.; Qiao, X.; Jiang, Y.; Tang, L.; Xu, Y.; et al. Oral recombinant Lactobacillus vaccine targeting the intestinal microfold cells and dendritic cells for delivering the core neutralizing epitope of porcine epidemic diarrhea virus. Microb. Cell Fact. 2018, 17, 20. [Google Scholar] [CrossRef] [PubMed]
- Wells, J. Mucosal vaccination and therapy with genetically modified lactic acid bacteria. Annu. Rev. Food Sci. Technol. 2011, 2, 423–445. [Google Scholar] [CrossRef]
- Owen, J.L.; Sahay, B.; Mohamadzadeh, M. New generation of oral mucosal vaccines targeting dendritic cells. Curr. Opin. Chem. Biol. 2013, 17, 918–924. [Google Scholar] [CrossRef] [Green Version]
- Mohamadzadeh, M.; Duong, T.; Sandwick, S.J.; Hoover, T.; Klaenhammer, T.R. Dendritic cell targeting of Bacillus anthracis protective antigen expressed by Lactobacillus acidophilus protects mice from lethal challenge. Proc. Natl. Acad. Sci. USA 2009, 106, 4331–4336. [Google Scholar] [CrossRef]
- Bruschke, C.J.; Moormann, R.J.; van Oirschot, J.T.; van Rijn, P.A. A subunit vaccine based on glycoprotein E2 of bovine virus diarrhea virus induces fetal protection in sheep against homologous challenge. Vaccine 1997, 15, 1940–1945. [Google Scholar] [CrossRef]
- Harpin, S.; Hurley, D.J.; Mbikay, M.; Talbot, B.; Elazhary, Y. Vaccination of cattle with a DNA plasmid encoding the bovine viral diarrhoea virus major glycoprotein E2. J. Gen. Virol. 1999, 80, 3137–3144. [Google Scholar] [CrossRef]
- Toth, R.L.; Nettleton, P.F.; McCrae, M.A. Expression of the E2 envelope glycoprotein of bovine viral diarrhoea virus (BVDV) elicits virus-type specific neutralising antibodies. Vet. Microbiol. 1999, 65, 87–101. [Google Scholar] [CrossRef]
- Nobiron, I.; Thompson, I.; Brownlie, J.; Collins, M.E. Cytokine adjuvancy of BVDV DNA vaccine enhances both humoral and cellular immune responses in mice. Vaccine 2001, 19, 4226–4235. [Google Scholar] [CrossRef]
- Xu, Y.X.; Ayala, A.; Monfils, B.; Cioffi, W.G.; Chaudry, I.H. Mechanism of intestinal mucosal immune dysfunction following trauma-hemorrhage: Increased apoptosis associated with elevated Fas expression in Peyer’s patches. J. Surg. Res. 1997, 70, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Xie, Y.; Li, X.; Guo, G.; Meng, Q.; Xiu, Y.; Li, T.; Feng, W.; Ma, L. The influence of Peyer’s patch apoptosis on intestinal mucosal immunity in burned mice. Burns 2009, 35, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Walz, P.H.; Grooms, D.L.; Passler, T.; Ridpath, J.F.; Tremblay, R.; Step, D.L.; Callan, R.J.; Givens, M.D. Control of bovine viral diarrhea virus in ruminants. J. Vet. Intern. Med. 2010, 24, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Newcomer, B.W.; Chamorro, M.F.; Walz, P.H. Vaccination of cattle against bovine viral diarrhea virus. Vet. Microbiol. 2017, 206, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Presi, P.; Struchen, R.; Knight-Jones, T.; Scholl, S.; Heim, D. Bovine viral diarrhea (BVD) eradication in Switzerland-Experiences of the first two years. Prev. Vet. Med. 2011, 99, 112–121. [Google Scholar] [CrossRef]
- Graham, D.A.; Lynch, M.; Coughlan, S.; Doherty, M.L.; O’Neill, R.; Sammin, D.; O’Flaherty, J. Development and review of the voluntary phase of a national BVD eradication programme in Ireland. Vet. Rec. 2014, 174, 67. [Google Scholar] [CrossRef]
- Duncan, A.J.; Gunn, G.J.; Humphry, R.W. Difficulties arising from the variety of testing schemes used for bovine viral diarrhoea virus (BVDV). Vet. Rec. 2016, 178, 292. [Google Scholar] [CrossRef]
- Wang, W.; Shi, X.; Tong, Q.; Wu, Y.; Xia, M.Q.; Ji, Y.; Xue, W.; Wu, H. A bovine viral diarrhea virus type 1a strain in China: Isolation, identification, and experimental infection in calves. Virol. J. 2014, 11, 8. [Google Scholar] [CrossRef]
- Cano-Garrido, O.; Seras-Franzoso, J.; Garcia-Fruitós, E. Lactic acid bacteria: Reviewing the potential of a promising delivery live vector for biomedical purposes. Microb. Cell Fact. 2015, 14, 137. [Google Scholar] [CrossRef]
- Heiss, S.; Hörmann, A.; Tauer, C.; Sonnleitner, M.; Egger, E.; Grabherr, R.; Heinl, S. Evaluation of novel inducible promoter/repressor systems for recombinant protein expression in Lactobacillus plantarum. Microb. Cell Fact. 2016, 15, 50. [Google Scholar] [CrossRef] [PubMed]
- Alander, M.; Satokari, R.; Korpela, R.; Saxelin, M.; Vilpponen-Salmela, T.; Mattila-Sandholm, T. Persistence of colonization of human colonic mucosa by a probiotic strain, Lactobacillus rhamnosus GG, after oral consumption. Appl. Environ. Microbiol. 1999, 65, 351–354. [Google Scholar] [PubMed]
- Xu, Y.; Li, Y. Induction of immune responses in mice after intragastric administration of Lactobacillus casei producing porcine parvovirus VP2 protein. Appl. Environ. Microbiol. 2007, 73, 7041–7047. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Asai, Y.; Yasuda, K.; Sakamoto, H. Oral immunoadjuvant activity of a newsymbiotic Lactobacillus casei subsp. casei in conjunction with dextran in BALB/cmice. Nutr. Res. 2005, 25, 295–304. [Google Scholar] [CrossRef]
- Remus, D.M.; van Kranenburg, R.; van Swam, I.I.; Taverne, N.; Bongers, R.S.; Wels, M.; Wells, J.M.; Bron, P.A.; Kleerebezem, M. Impact of 4 Lactobacillus plantarum capsular polysaccharide clusters on surface glycan composition and host cell signaling. Microb. Cell Fact. 2012, 11, 149. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Guo, Z.; Liu, J.; Wang, Z.; Wang, R.; Li, Y.; Wang, L.; Xu, Y.; Tang, L.; Qiao, X. Recombinant Lactobacillus casei expressing Clostridium perfringens toxoids α, β2, ε and β1 gives protection against Clostridium perfringens in rabbits. Vaccine 2017, 35, 4010–4021. [Google Scholar] [CrossRef]
- Hoang, V.V.; Ochi, T.; Kurata, K.; Arita, Y.; Ogasahara, Y.; Enomoto, K. Nisin-induced expression of recombinant T cell epitopes of major Japanese cedar pollen allergens in Lactococcus lactis. Appl. Microbiol. Biotechnol. 2018, 102, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Ma, Y.; Wang, Z.; Bai, J.; Jia, S.; Feng, B.; Jiang, Y.; Cui, W.; Tang, L.; Li, Y.; et al. Oral immunization of mice with a probiotic Lactobacillus casei constitutively expressing the α-toxoid induces protective immunity against Clostridium perfringens α-toxin. Virulence 2019, 10, 166–179. [Google Scholar] [CrossRef]
- Seong, G.; Oem, J.K.; Lee, K.H.; Choi, K.S. Experimental infection of mice with bovine viral diarrhea virus. Arch. Virol. 2015, 160, 1565–1571. [Google Scholar] [CrossRef]
- Lee, K.H.; Han, D.G.; Kim, S.; Choi, E.J.; Choi, K.S. Experimental infection of mice with noncytopathic bovine viral diarrhea virus 2 increases the number of megakaryocytes in bone marrow. Virol. J. 2018, 15, 115. [Google Scholar] [CrossRef]
- Xue, W.; Mattick, D.; Smith, L.; Umbaugh, J.; Trigo, E. Vaccination with a modified-live bovine viral diarrhea virus (BVDV) type 1a vaccine completely protected calves against challenge with BVDV type 1b strains. Vaccine 2010, 29, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Palomares, R.A.; Givens, M.D.; Wright, J.C.; Walz, P.H.; Brock, K.V. Valuation of the onset of protection induced by a modified-live virus vaccine in calves challenge inoculated with type 1b bovine viral diarrhea virus. Am. J. Vet. Res. 2012, 73, 567–574. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′–3′) | Production Size |
|---|---|---|
| BVDV-E2-F | CGAGCTCaATGCTCCCAGCCTGTAAACC | 1122 bp |
| BVDV-E2-R | TTTGGGCCCaTTAACCTAAGGTCGTTTGTTCTGAT | |
| E2-DCpep | GGGCCCa TGGACGTTGTGGAGTTGAATGATATGATGGATAAAAbTGAGCCACCGCCACCcTTAACCTAAGGTCGTTTGTTC | |
| BVDV-F | GGTAGCAACAGTGGTGAG | 221 bp |
| BVDV-R | GTAGCAATACAGTGGGCC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Feng, B.; Niu, C.; Jia, S.; Sun, C.; Wang, Z.; Jiang, Y.; Cui, W.; Wang, L.; Xu, Y. Dendritic Cell Targeting of Bovine Viral Diarrhea Virus E2 Protein Expressed by Lactobacillus casei Effectively Induces Antigen-Specific Immune Responses via Oral Vaccination. Viruses 2019, 11, 575. https://doi.org/10.3390/v11060575
Wang Y, Feng B, Niu C, Jia S, Sun C, Wang Z, Jiang Y, Cui W, Wang L, Xu Y. Dendritic Cell Targeting of Bovine Viral Diarrhea Virus E2 Protein Expressed by Lactobacillus casei Effectively Induces Antigen-Specific Immune Responses via Oral Vaccination. Viruses. 2019; 11(6):575. https://doi.org/10.3390/v11060575
Chicago/Turabian StyleWang, Yixin, Baohua Feng, Chao Niu, Shuo Jia, Chao Sun, Zhuo Wang, Yanping Jiang, Wen Cui, Li Wang, and Yigang Xu. 2019. "Dendritic Cell Targeting of Bovine Viral Diarrhea Virus E2 Protein Expressed by Lactobacillus casei Effectively Induces Antigen-Specific Immune Responses via Oral Vaccination" Viruses 11, no. 6: 575. https://doi.org/10.3390/v11060575
APA StyleWang, Y., Feng, B., Niu, C., Jia, S., Sun, C., Wang, Z., Jiang, Y., Cui, W., Wang, L., & Xu, Y. (2019). Dendritic Cell Targeting of Bovine Viral Diarrhea Virus E2 Protein Expressed by Lactobacillus casei Effectively Induces Antigen-Specific Immune Responses via Oral Vaccination. Viruses, 11(6), 575. https://doi.org/10.3390/v11060575