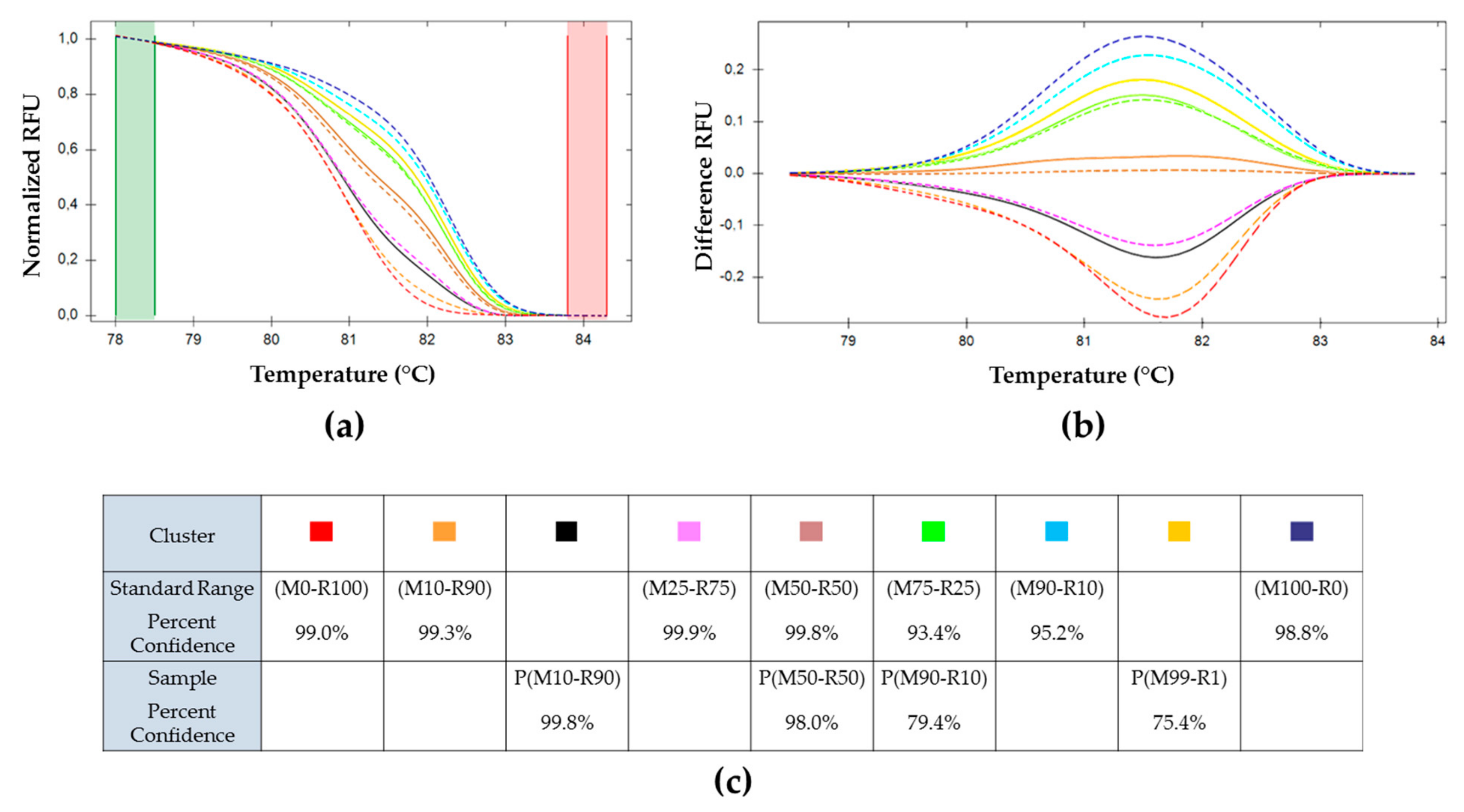
A Combination of Real-Time PCR and High-Resolution Melting Analysis to Detect and Identify CpGV Genotypes Involved in Type I Resistance



{kind=link}
{kind=link}
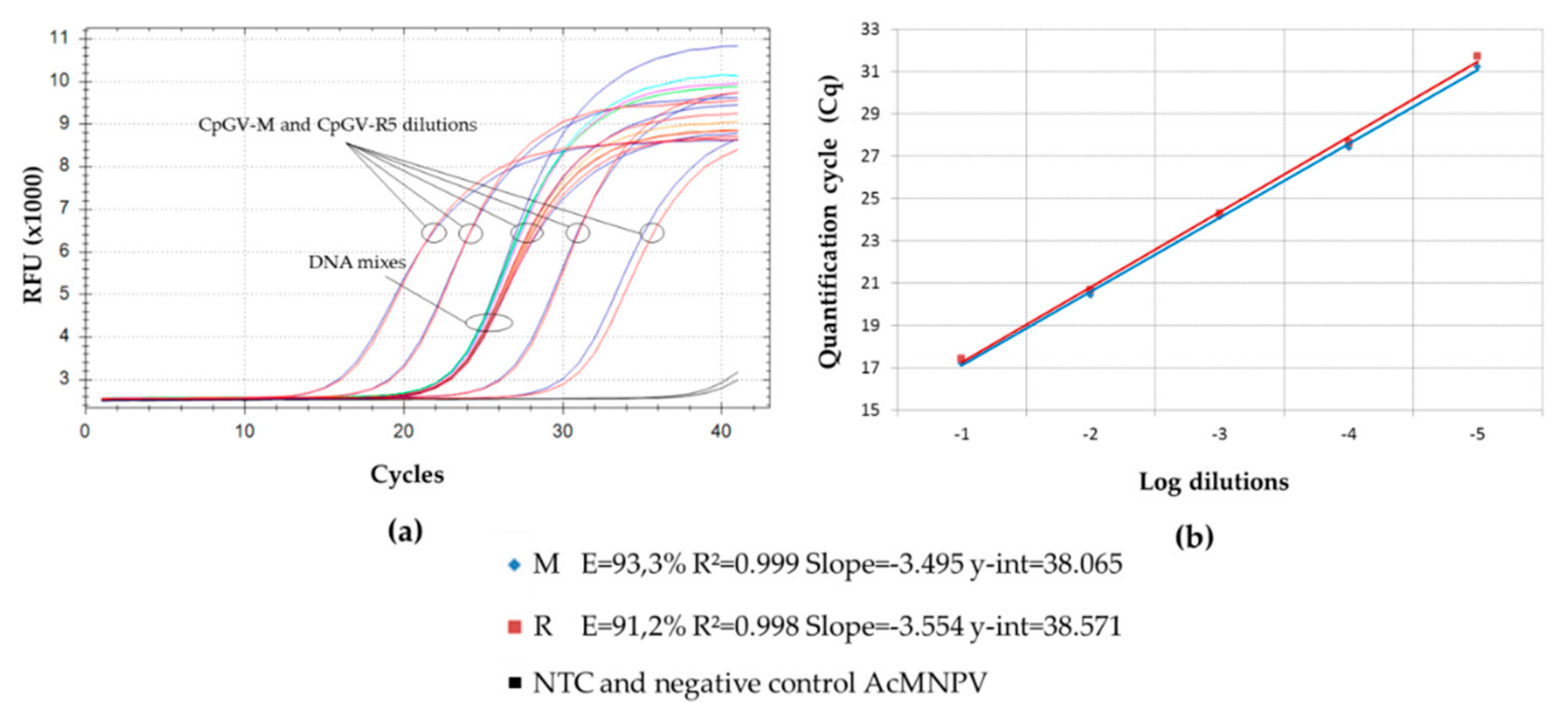
{kind=link}
{kind=link}
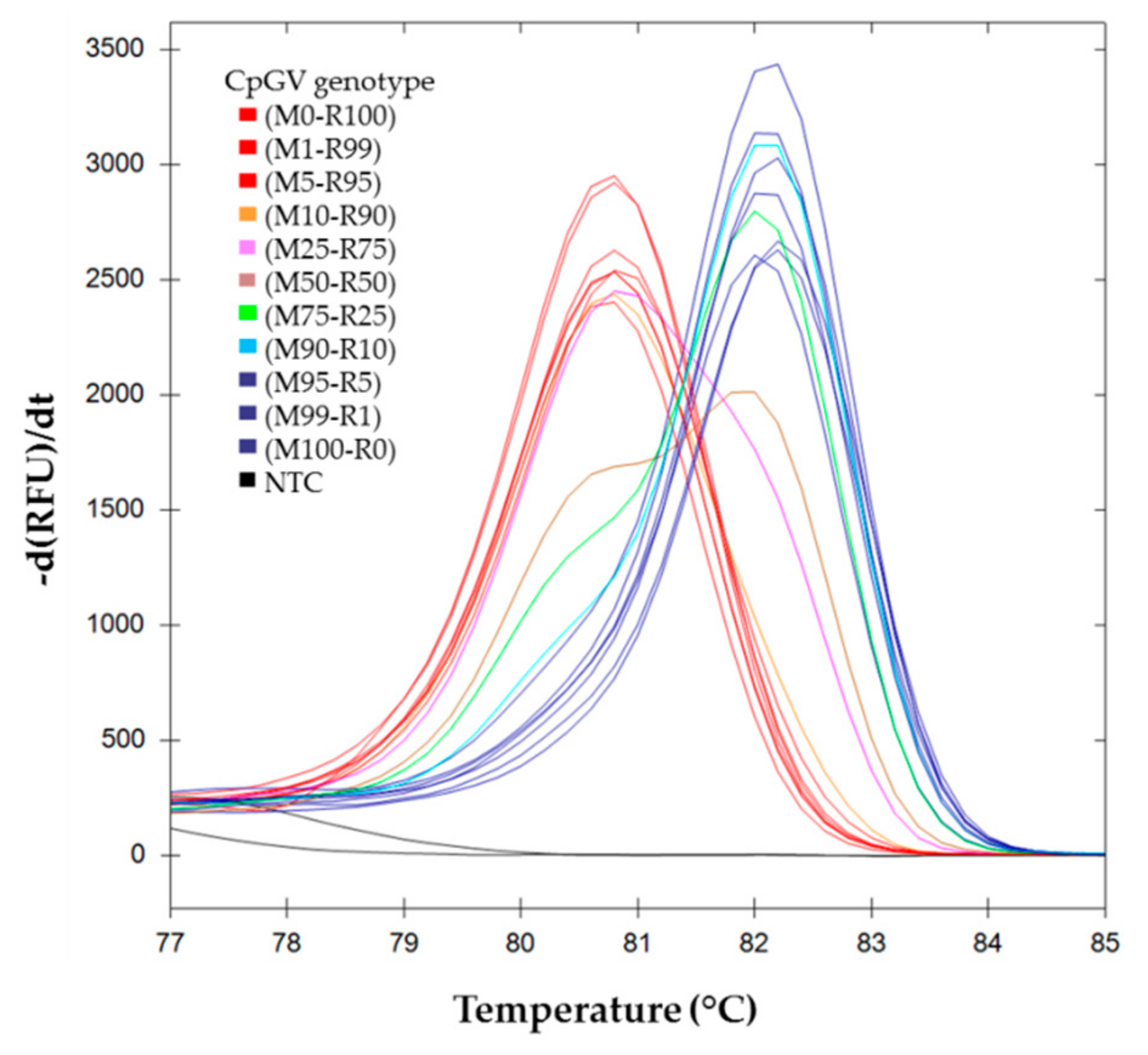
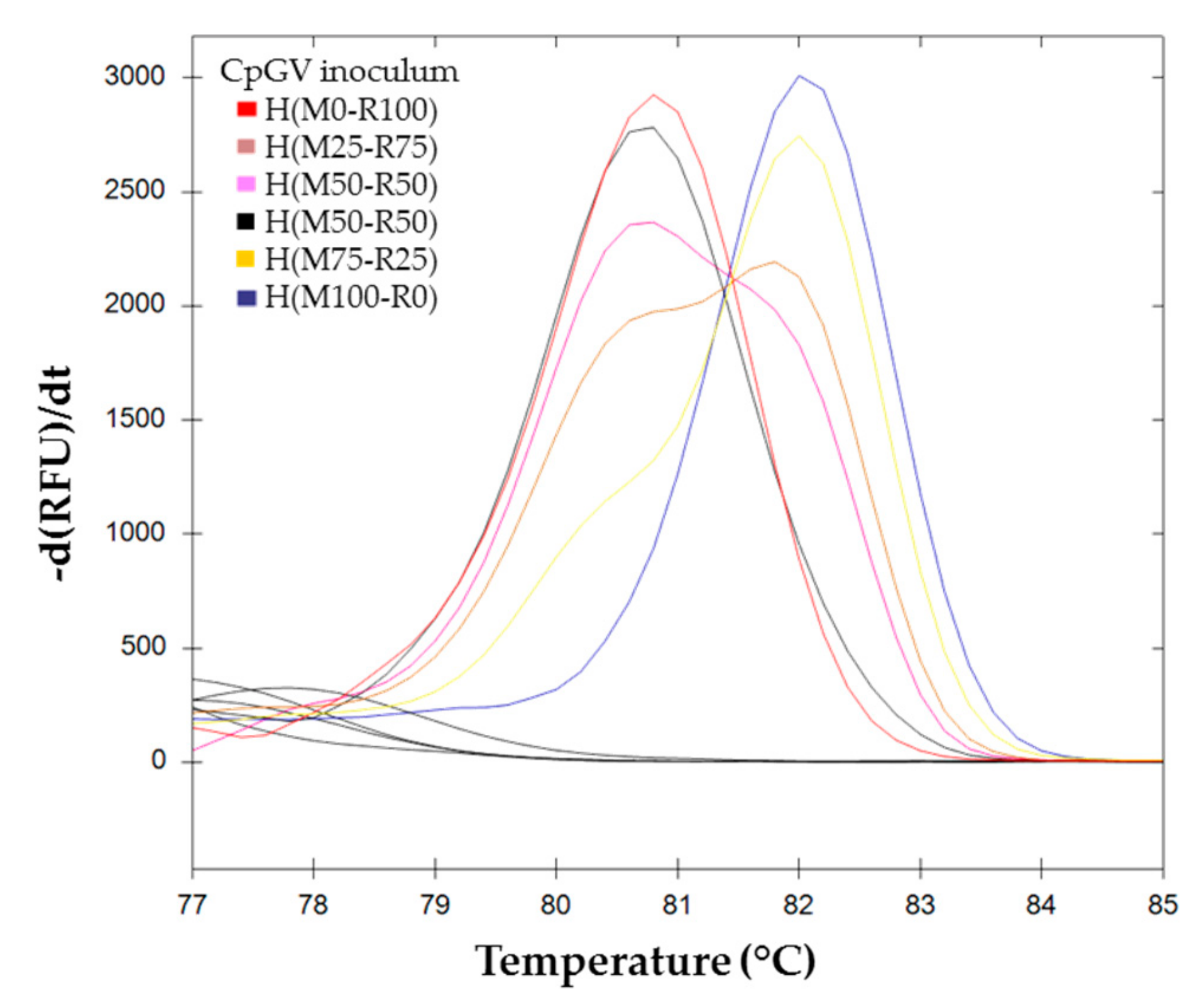
{kind=link}
{kind=link}
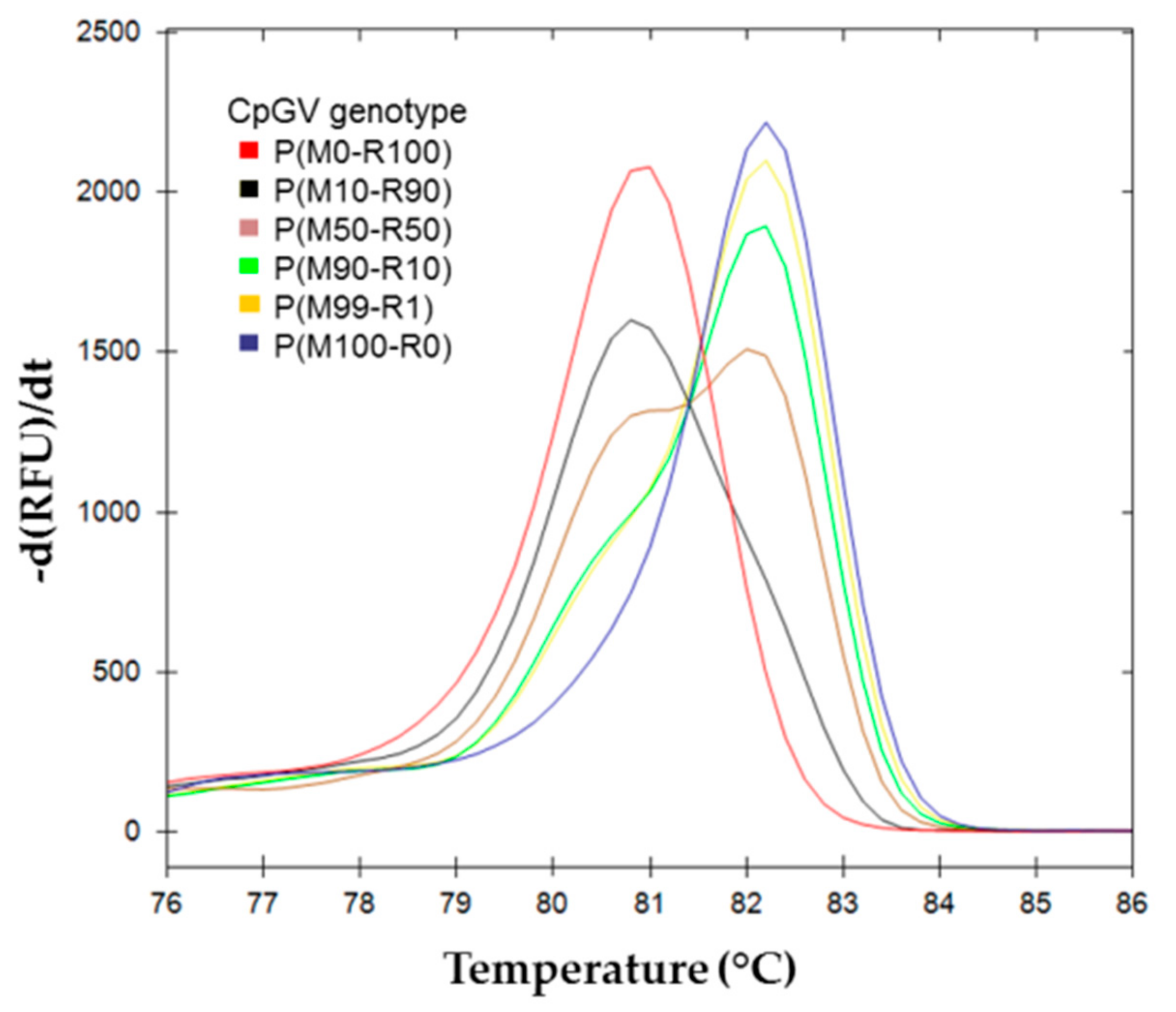
{kind=link}
Abstract
Share and Cite
Hinsberger, A.; Theulier Saint Germain, S.; Guerrero, P.; Blachère-López, C.; López-Ferber, M.; Bayle, S. A Combination of Real-Time PCR and High-Resolution Melting Analysis to Detect and Identify CpGV Genotypes Involved in Type I Resistance. Viruses 2019, 11, 723. https://doi.org/10.3390/v11080723
Hinsberger A, Theulier Saint Germain S, Guerrero P, Blachère-López C, López-Ferber M, Bayle S. A Combination of Real-Time PCR and High-Resolution Melting Analysis to Detect and Identify CpGV Genotypes Involved in Type I Resistance. Viruses. 2019; 11(8):723. https://doi.org/10.3390/v11080723
Chicago/Turabian StyleHinsberger, Aurélie, Stéphane Theulier Saint Germain, Patrice Guerrero, Christine Blachère-López, Miguel López-Ferber, and Sandrine Bayle. 2019. "A Combination of Real-Time PCR and High-Resolution Melting Analysis to Detect and Identify CpGV Genotypes Involved in Type I Resistance" Viruses 11, no. 8: 723. https://doi.org/10.3390/v11080723
APA StyleHinsberger, A., Theulier Saint Germain, S., Guerrero, P., Blachère-López, C., López-Ferber, M., & Bayle, S. (2019). A Combination of Real-Time PCR and High-Resolution Melting Analysis to Detect and Identify CpGV Genotypes Involved in Type I Resistance. Viruses, 11(8), 723. https://doi.org/10.3390/v11080723