Enterovirus 71 VP1 Protein Regulates Viral Replication in SH-SY5Y Cells via the mTOR Autophagy Signaling Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies and Chemical Reagents
2.2. Cell Lines and Viruses
2.3. Virus Infection
2.4. Replication Kinetics
2.5. Lactate Dehydrogenase (LDH) Cytotoxicity Assay
2.6. Drugs Used to Induce or Inhibit Autophagy
2.7. Detection of Autophagosomes, Viruses, and mTOR in Cells by Immunofluorescent Assay
2.8. Detection of Autophagy-Related Protein and Virus VP1 Protein by Western Blot
3. Results
3.1. Construction and Rescue of Recombinant Virus SDLY107-VP1
3.2. Recombinant Virus Biological Characteristics
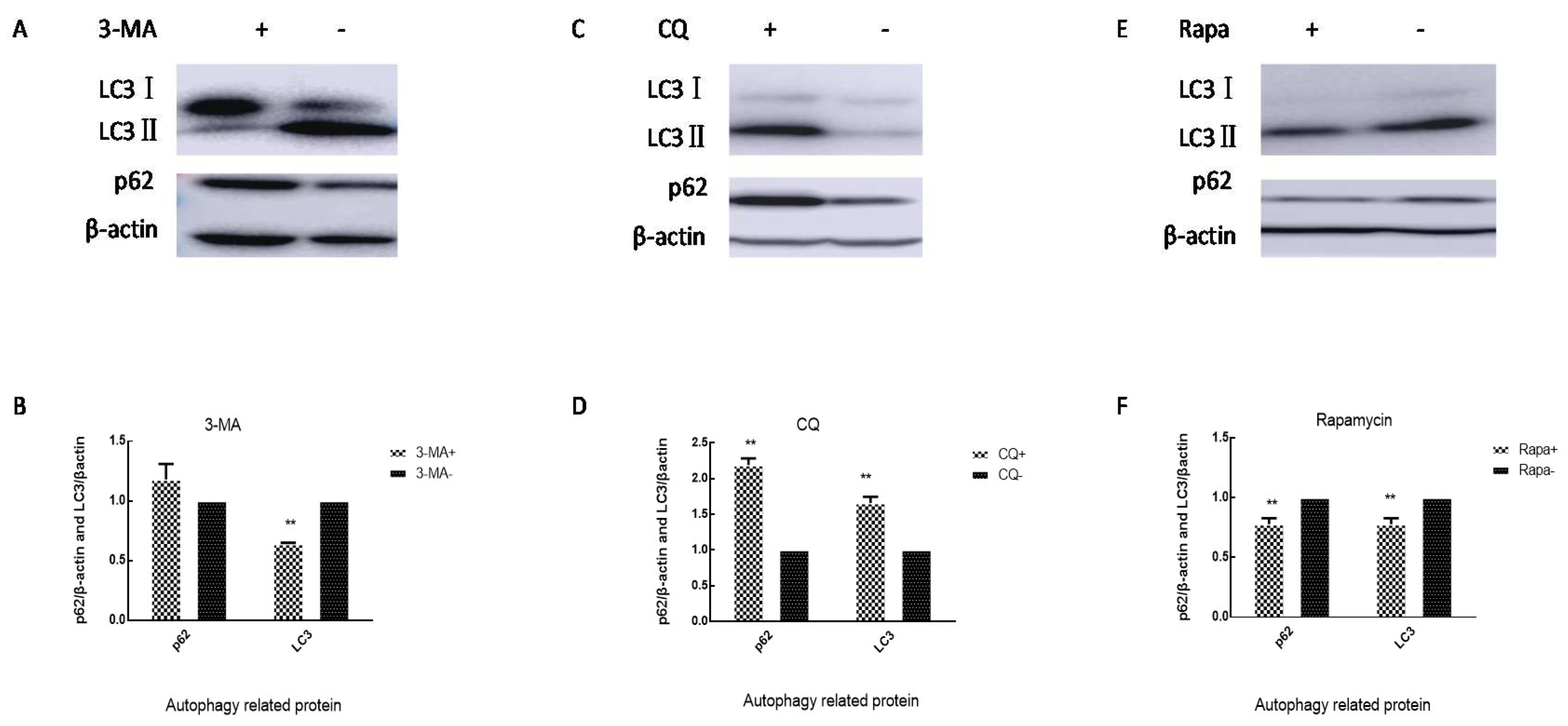
3.3. Autophagic Flux in SH-SY5Y Cells Is Inhibited by EV71 VP1 Protein
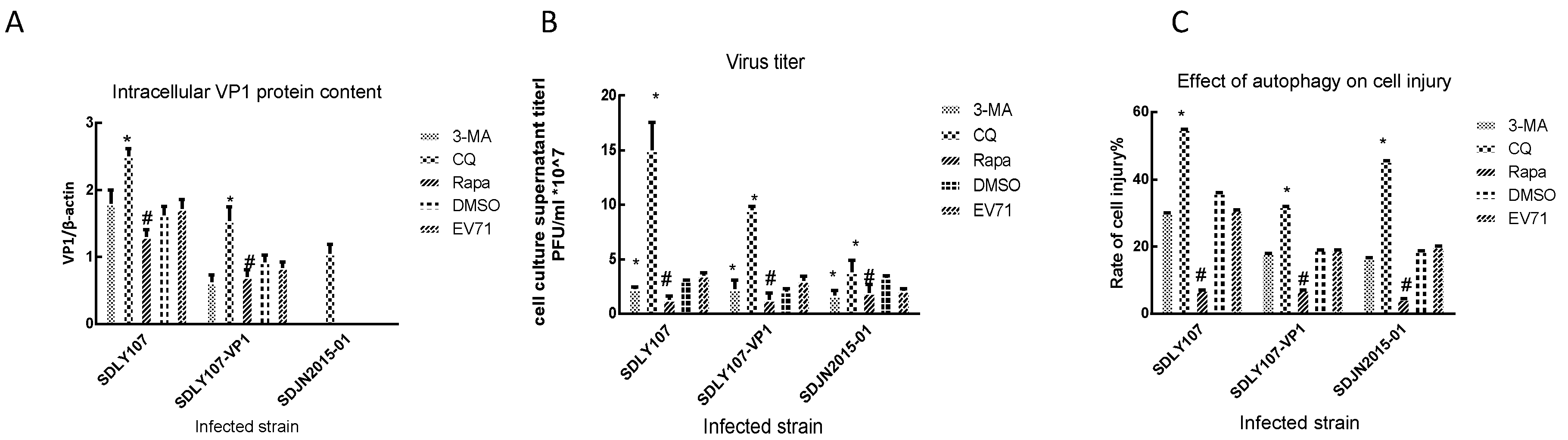
3.4. VP1 Protein Affects Virus Replication Associated with Cell Autophagy
3.5. Autophagy Affects the Survival of Nerve Cells Infected with EV71
3.6. mTOR Is a Key Molecule Affecting EV71 Replication in Nerve Cells
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ayukekbong, J.A.; Bergstrom, T. Polio will go, acute flaccid paralysis will stay. Lancet 2014, 383, 2209–2210. [Google Scholar] [CrossRef]
- Nassef, C.; Ziemer, C.; Morrell, D.S. Hand-foot-and-mouth disease: A new look at a classic viral rash. Curr. Opin. Pediatr. 2015, 27, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yeo, A.; Phoon, M.C.; Tan, E.L.; Poh, C.L.; Quak, S.H.; Chow, V.T. The largest outbreak of hand; foot and mouth disease in Singapore in 2008: The role of enterovirus 71 and coxsackievirus A strains. Int. J. Infect. Dis. 2010, 14, 1076–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, L.G.; Parashar, U.D.; Lye, M.S.; Ong, F.G.; Zaki, S.R.; Alexander, J.P.; Ho, K.K.; Han, L.L.; Pallansch, M.A.; Suleiman, A.B.; et al. Deaths of children during an outbreak of hand, foot, and mouth disease in sarawak, malaysia: Clinical and pathological characteristics of the disease. For the Outbreak Study Group. Clin. Infect. Dis. 2000, 31, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Van Tu, P.; Thao, N.T.T.; Perera, D.; Truong, K.H.; Tien, N.T.K.; Thuong, T.C.; How, O.M.; Cardosa, M.J.; McMinn, P.C. Epidemiologic and virologic investigation of hand, foot, and mouth disease, southern Vietnam, 2005. Emerg. Infect. Dis. 2007, 13, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.N.; Zhang, X.Z.; Wang, B.; Qiu, Y.T. Etiologic and epidemiologic analysis of hand, foot, and mouth disease in Guangzhou city: A review of 4,753 cases. Braz. J. Infect. Dis. 2012, 16, 457–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, G.L.; Dickson, K.E.; Waters, M.J.; Kennett, M.L.; Land, S.A.; Sneddon, M. Outbreak of enterovirus 71 infection in Victoria, Australia, with a high incidence of neurologic involvement. Pediatr. Infect. Dis. J. 1988, 7, 484–488. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Nakano, S.; Yamaoka, K.; Takami, S. Outbreaks of hand, foot, and mouth disease by enterovirus 71. High incidence of complication disorders of central nervous system. Arch. Dis. Child. 1980, 55, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Ng, Q.; He, F.; Kwang, J. Recent Progress towards Novel EV71 Anti-Therapeutics and Vaccines. Viruses 2015, 7, 6441–6457. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.I.; Song, J.H.; Kwon, B.E.; Kim, H.N.; Seo, M.D.; Park, K.; Lee, S.; Yeo, S.G.; Kweon, M.N.; Ko, H.J.; et al. Pros and cons of VP1-specific maternal IgG for the protection of Enterovirus 71 infection. Vaccine 2015, 33, 6604–6610. [Google Scholar] [CrossRef]
- Nishimura, Y.; Lee, H.; Hafenstein, S.; Kataoka, C.; Wakita, T.; Bergelson, J.M.; Shimizu, H. Enterovirus 71 binding to PSGL-1 on leukocytes: VP1-145 acts as a molecular switch to control receptor interaction. PLoS Pathog. 2013, 9, 1003511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Duo, J.; Liu, J.; Ma, C.; Zhang, L.; Wei, Q.; Qin, C. A mouse muscle-adapted enterovirus 71 strain with increased virulence in mice. Microbes Infect. 2011, 13, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Mizuta, K.; Aoki, Y.; Suto, A.; Ootani, K.; Katsushima, N.; Itagaki, T.; Ohmi, A.; Okamoto, M.; Nishimura, H.; Matsuzaki, Y.; et al. Cross-antigenicity among EV71 strains from different genogroups isolated in Yamagata, Japan, between 1990 and 2007. Vaccine 2009, 27, 3153–3158. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.W.; Hsu, Y.W.; Smith, D.J.; Kiang, D.; Tsai, H.P.; Lin, K.H.; Wang, S.M.; Liu, C.C.; Su, I.J.; Wang, J.R. Reemergence of enterovirus 71 in 2008 in taiwan: Dynamics of genetic and antigenic evolution from 1998 to 2008. J. Clin. Microbiol. 2009, 47, 3653–3662. [Google Scholar] [CrossRef] [Green Version]
- Haolong, C.; Du, N.; Hongchao, T.; Yang, Y.; Wei, Z.; Hua, Z.; Wenliang, Z.; Lei, S.; Po, T. Enterovirus 71 VP1 activates calmodulin-dependent protein kinase II and results in the rearrangement of vimentin in human astrocyte cells. PLoS ONE 2013, 8, 73900. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.L.; Chou, Y.T.; Wu, C.N.; Ho, M.S. Annexin II binds to capsid protein VP1 of enterovirus 71 and enhances viral infectivity. J. Virol. 2011, 85, 11809–11820. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.X.; Huang, Y.M.; Li, Q.J.; Li, X.Y.; Zhou, Y.D.; Guo, F.; Zhou, J.M.; Cen, S. A highly conserved amino acid in VP1 regulates maturation of enterovirus 71. PLoS Pathog. 2017, 13, 1006625. [Google Scholar] [CrossRef]
- Li, X.; Huang, Y.; Sun, M.; Ji, H.; Dou, H.; Hu, J.; Yan, Y.; Wang, X.; Chen, L. Honeysuckle-encoded microRNA2911 inhibits Enterovirus 71 replication via targeting VP1 gene. Antiviral Res. 2018, 152, 117–123. [Google Scholar] [CrossRef]
- Xi, X.; Zhang, X.; Wang, B.; Wang, T.; Wang, J.; Huang, H.; Wang, J.; Jin, Q.; Zhao, Z. The interplays between autophagy and apoptosis induced by enterovirus 71. PLoS ONE 2013, 8, 56966. [Google Scholar] [CrossRef] [Green Version]
- Too, I.H.; Yeo, H.; Sessions, O.M.; Yan, B.; Libau, E.A.; Howe, J.L.; Lim, Z.Q.; Suku-Maran, S.; Ong, W.Y.; Chua, K.B.; et al. Enterovirus 71 infection of motor neuron-like NSC-34 cells undergoes a non-lytic exit pathway. Sci. Rep. 2016, 6, 36983. [Google Scholar] [CrossRef]
- Li, P.; Yang, S.; Hu, D.; Wei, D.; Lu, J.; Zheng, H.; Nie, S.; Liu, G.; Yang, H. Enterovirus 71 VP1 promotes mouse Schwann cell autophagy via ER stressmediated PMP22 upregulation. Int. J. Mol. Med. 2019, 44, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhang, M.; Zhang, X.; Tan, X.; Guo, H.; Zeng, W.; Yan, G.; Memon, A.M.; Li, Z.; Zhu, Y.; et al. Porcine Epidemic Diarrhea Virus Induces Autophagy to Benefit Its Replication. Viruses 2017, 9, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.; Li, X.; Cai, Q.; Li, C.; Tian, L.; Chen, J.; Xing, X.; Gan, Y.; Ouyang, W.; Yang, Z.; et al. The PI3K/Akt/mTOR pathway is involved in CVB3-induced autophagy of HeLa cells. Int. J. Mol. Med. 2017, 40, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Q.; Li, J.; Sun, C.X.; Ma, Y.W.; Zhuang, Z.C.; Sun, L.L.; Zhao, L.; Wang, Z.Y.; Wen, H.L. Construction and identification of a chimeric cDNA infectious clone of enterovirus 71 strain. Chin. J. Microbiol. Immunol. 2015, 35, 87–93. [Google Scholar] [CrossRef]
- Wen, H.L.; Si, L.Y.; Yuan, X.J.; Hao, S.B.; Gao, F.; Chu, F.L.; Sun, C.X.; Wang, Z.Y. Complete genome sequencing and analysis of six enterovirus 71 strains with different clinical phenotypes. Virol. J. 2013, 10, 115. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Qiao, Q.; Hao, S.B.; Dong, Z.; Zhao, L.; Ji, J.; Wang, Z.Y.; Wen, H.L. Nonstructural protein 2A modulates replication and virulence of enterovirus 71. Virus Res. 2018, 244, 262–269. [Google Scholar] [CrossRef]
- Dong, Z.; Liu, Z.W.; Chen, R.; Wen, X.J.; Ji, J.; Zheng, X.X.; Zhao, L.; Wang, Z.Y.; Wen, H.L. The untranslated regions of EV-A71 contribute to its pathogenicity and virulence. Virus Res. 2019, 263, 55–63. [Google Scholar] [CrossRef]
- Ma, Y.W.; Hao, S.B.; Sun, L.L.; Li, J.; Qiao, Q.; Gao, F.; Zhao, L.; Yu, X.J.; Wang, Z.Y.; Wen, H.L.; et al. Construction and characterization of infectious cDNA clones of enterovirus 71 (EV71). Virol. Sin. 2015, 30, 305–308. [Google Scholar] [CrossRef]
- Sun, L.L.; Wang, J.K.; Cui, X.Q.; Hao, S.B.; Li, J.; Zhao, L.; Yuan, X.J.; Wen, H.L.; Yu, X.J.; Wang, Z.Y.; et al. Association of viral replication capacity with the pathogenicity of enterovirus 71. Virus Res. 2014, 189, 1–7. [Google Scholar] [CrossRef]
- Watanabe, R.; Wei, L.; Huang, J. mTOR signaling, function, novel inhibitors, and therapeutic targets. J. Nucl. Med. 2011, 52, 497–500. [Google Scholar] [CrossRef] [Green Version]
- Bruning, A.H.L.; Thomas, X.V.; van der Linden, L.; Wildenbeest, J.G.; Minnaar, R.P.; Jansen, R.R.; de Jong, M.D.; Sterk, P.J.; van der Schee, M.P.; Wolthers, K.C.; et al. Clinical, virological and epidemiological characteristics of rhinovirus infections in early childhood: A comparison between non-hospitalised and hospitalised children. J. Clin. Virol. 2015, 73, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.R.; Schinkel, J.; Dek, I.; Koekkoek, S.M.; Visser, C.E.; de Jong, M.D.; Molenkamp, R.; Pajkrt, D. Quantitation of respiratory viruses in relation to clinical course in children with acute respiratory tract infections. Pediatr. Infect. Dis. J. 2010, 29, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Kemball, C.C.; Alirezaei, M.; Flynn, C.T.; Wood, M.R.; Harkins, S.; Kiosses, W.B.; Whitton, J.L. Coxsackievirus infection induces autophagy-like vesicles and megaphagosomes in pancreatic acinar cells in vivo. J. Virol. 2010, 84, 12110–12124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouna, L.; Hernandez, E.; Bonte, D.; Brost, R.; Amazit, L.; Delgui, L.R.; Brune, W.; Geballe, A.P.; Beau, I.; Esclatine, A.; et al. Analysis of the role of autophagy inhibition by two complementary human cytomegalovirus BECN1/Beclin 1-binding proteins. Autophagy 2016, 12, 327–342. [Google Scholar] [CrossRef] [Green Version]
- Ke PY, Chen SS: Autophagy in hepatitis C virus-host interactions: Potential roles and therapeutic targets for liver-associated diseases. World J. Gastroenterol. 2014, 20, 5773–5793. [CrossRef]
- Talloczy, Z.; Jiang, W.; Virgin HWt Leib, D.A.; Scheuner, D.; Kaufman, R.J.; Eskelinen, E.L.; Levine, B. Regulation of starvation- and virus-induced autophagy by the eIF2alpha kinase signaling pathway. Proc. Natl. Acad. Sci. USA 2002, 99, 190–195. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.; Zhang, J.; Si, X.; Gao, G.; Mao, I.; McManus, B.M.; Luo, H. Autophagosome supports coxsackievirus B3 replication in host cells. J. Virol. 2008, 82, 9143–9153. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Jiang, X.; Liu, D.; Fan, Z.; Hu, X.; Yan, J.; Wang, M.; Gao, G.F. Autophagy is involved in influenza A virus replication. Autophagy 2009, 5, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Espert, L.; Denizot, M.; Grimaldi, M.; Robert-Hebmann, V.; Gay, B.; Varbanov, M.; Codogno, P.; Biard-Piechaczyk, M. Autophagy is involved in T cell death after binding of HIV-1 envelope proteins to CXCR4. J. Clin. Investig. 2006, 116, 2161–2172. [Google Scholar] [CrossRef]
- Fu, Y.; Xu, W.; Chen, D.; Feng, C.; Zhang, L.; Wang, X.; Lv, X.; Zheng, N.; Jin, Y.; Wu, Z. Enterovirus 71 induces autophagy by regulating has-miR-30a expression to promote viral replication. Antiviral Res. 2015, 124, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.R.; Wang, P.S.; Wang, J.R.; Liu, H.S. Enterovirus 71-induced autophagy increases viral replication and pathogenesis in a suckling mouse model. J. Biomed. Sci. 2014, 21, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henault, J.; Martinez, J.; Riggs, J.M.; Tian, J.; Mehta, P.; Clarke, L.; Sasai, M.; Latz, E.; Brinkmann, M.M.; Iwasaki, A.; et al. Noncanonical autophagy is required for type I interferon secretion in response to DNA-immune complexes. Immunity 2012, 37, 986–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paludan, C.; Schmid, D.; Landthaler, M.; Vockerodt, M.; Kube, D.; Tuschl, T.; Munz, C. Endogenous MHC class II processing of a viral nuclear antigen after autophagy. Science 2005, 307, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Buffen, K.; Oosting, M.; Quintin, J.; Ng, A.; Kleinnijenhuis, J.; Kumar, V.; van de Vosse, E.; Wijmenga, C.; van Crevel, R.; Oosterwijk, E.; et al. Autophagy controls BCG-induced trained immunity and the response to intravesical BCG therapy for bladder cancer. PLoS Pathog. 2014, 10, 1004485. [Google Scholar] [CrossRef] [PubMed]
- Kyei, G.B.; Dinkins, C.; Davis, A.S.; Roberts, E.; Singh, S.B.; Dong, C.; Wu, L.; Kominami, E.; Ueno, T.; Yamamoto, A.; et al. Autophagy pathway intersects with HIV-1 biosynthesis and regulates viral yields in macrophages. J. Cell. Biol. 2009, 186, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Heras-Sandoval, D.; Perez-Rojas, J.M.; Hernandez-Damian, J.; Pedraza-Chaverri, J. The role of PI3K/AKT/mTOR pathway in the modulation of autophagy and the clearance of protein aggregates in neurodegeneration. Cell Signal. 2014, 26, 2694–2701. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.-W.; Zhuang, Z.-C.; Chen, R.; Wang, X.-R.; Zhang, H.-L.; Li, S.-H.; Wang, Z.-Y.; Wen, H.-L. Enterovirus 71 VP1 Protein Regulates Viral Replication in SH-SY5Y Cells via the mTOR Autophagy Signaling Pathway. Viruses 2020, 12, 11. https://doi.org/10.3390/v12010011
Liu Z-W, Zhuang Z-C, Chen R, Wang X-R, Zhang H-L, Li S-H, Wang Z-Y, Wen H-L. Enterovirus 71 VP1 Protein Regulates Viral Replication in SH-SY5Y Cells via the mTOR Autophagy Signaling Pathway. Viruses. 2020; 12(1):11. https://doi.org/10.3390/v12010011
Chicago/Turabian StyleLiu, Zi-Wei, Zhi-Chao Zhuang, Rui Chen, Xiao-Rui Wang, Hai-Lu Zhang, Shu-Han Li, Zhi-Yu Wang, and Hong-Ling Wen. 2020. "Enterovirus 71 VP1 Protein Regulates Viral Replication in SH-SY5Y Cells via the mTOR Autophagy Signaling Pathway" Viruses 12, no. 1: 11. https://doi.org/10.3390/v12010011
APA StyleLiu, Z.-W., Zhuang, Z.-C., Chen, R., Wang, X.-R., Zhang, H.-L., Li, S.-H., Wang, Z.-Y., & Wen, H.-L. (2020). Enterovirus 71 VP1 Protein Regulates Viral Replication in SH-SY5Y Cells via the mTOR Autophagy Signaling Pathway. Viruses, 12(1), 11. https://doi.org/10.3390/v12010011