A Comprehensive Strategy for Screening for Xenotransplantation-Relevant Viruses in a Second Isolated Population of Göttingen Minipigs


Abstract
:1. Introduction
2. Materials and Methods
2.1. Pig Holding Including Absence of Vaccination and Food Additives
2.2. Blood and Isolation of PBMCs
2.3. General Screening Strategy
2.4. DNA and RNA Isolation
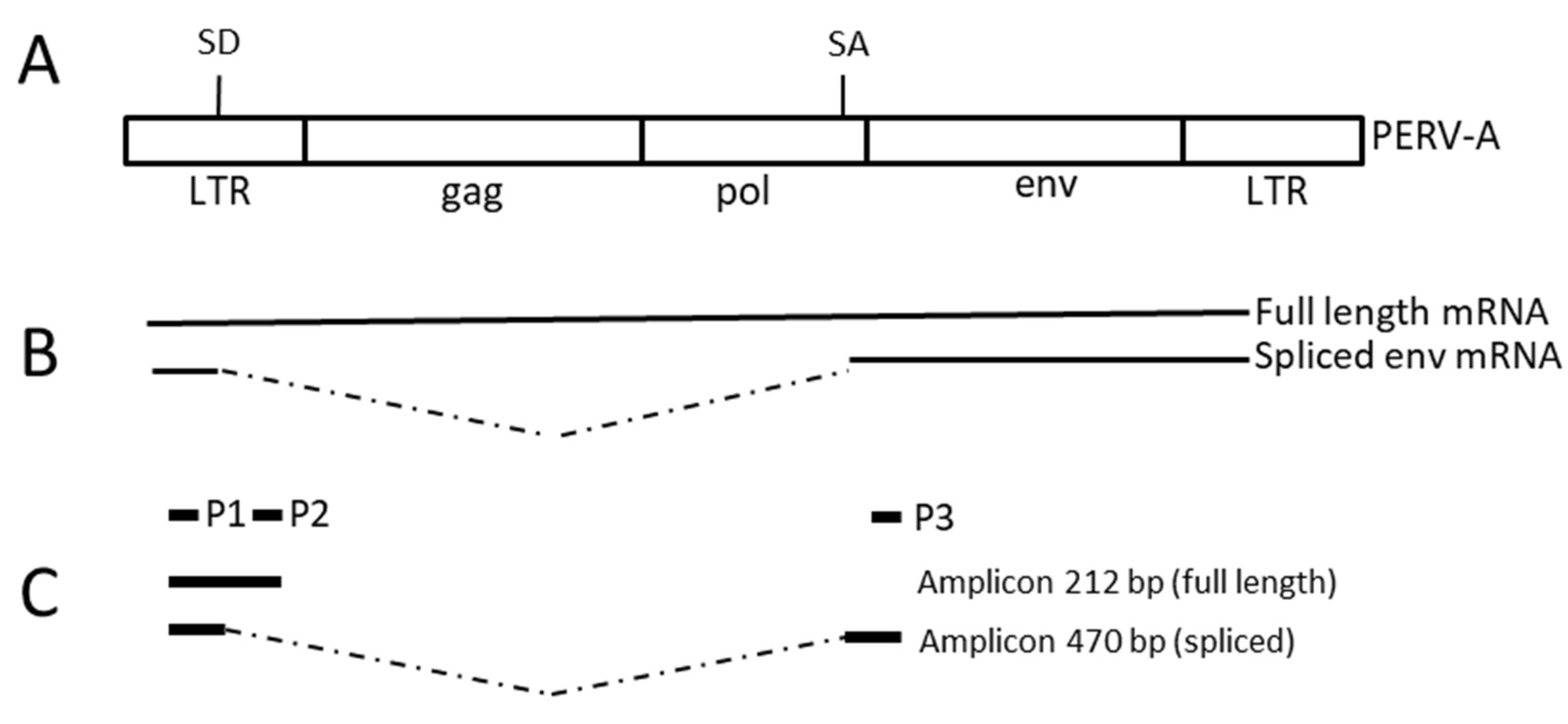
2.5. PCR, RT-PCR and Real-Time PCR
2.6. Mitogen Stimulation and Co-Culture
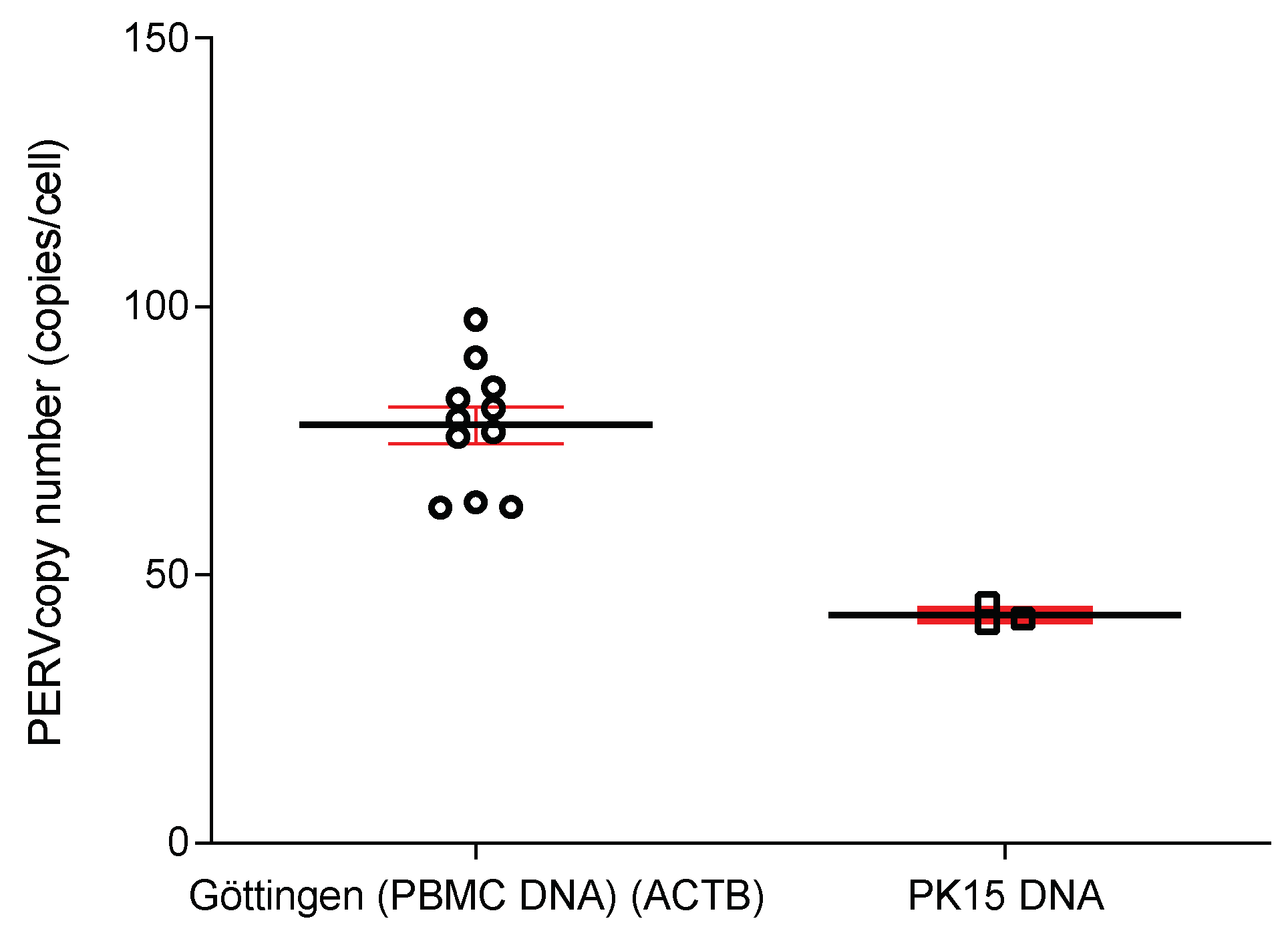
2.7. Determination of the PERV Copy Number
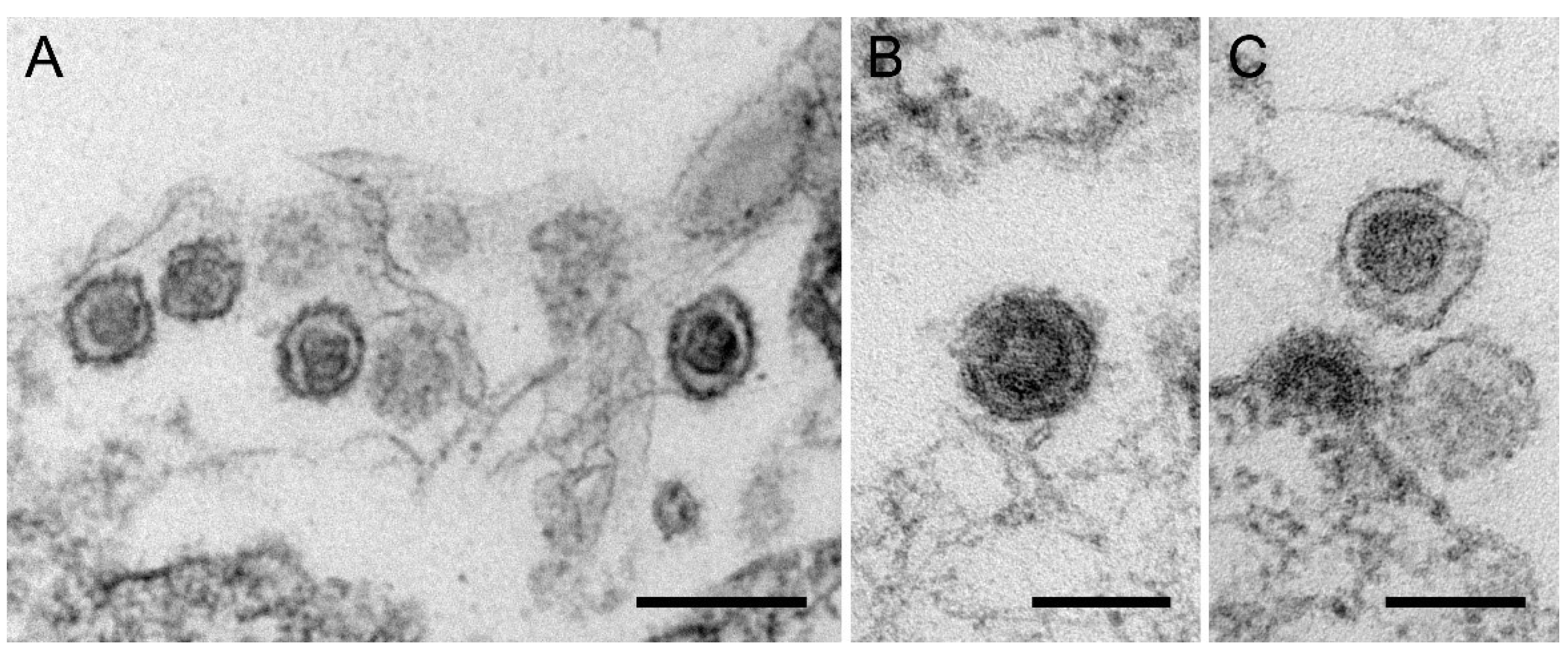
2.8. Transmission Electron Microscopy
3. Results
3.1. Brief History of Göttingen Minipgs
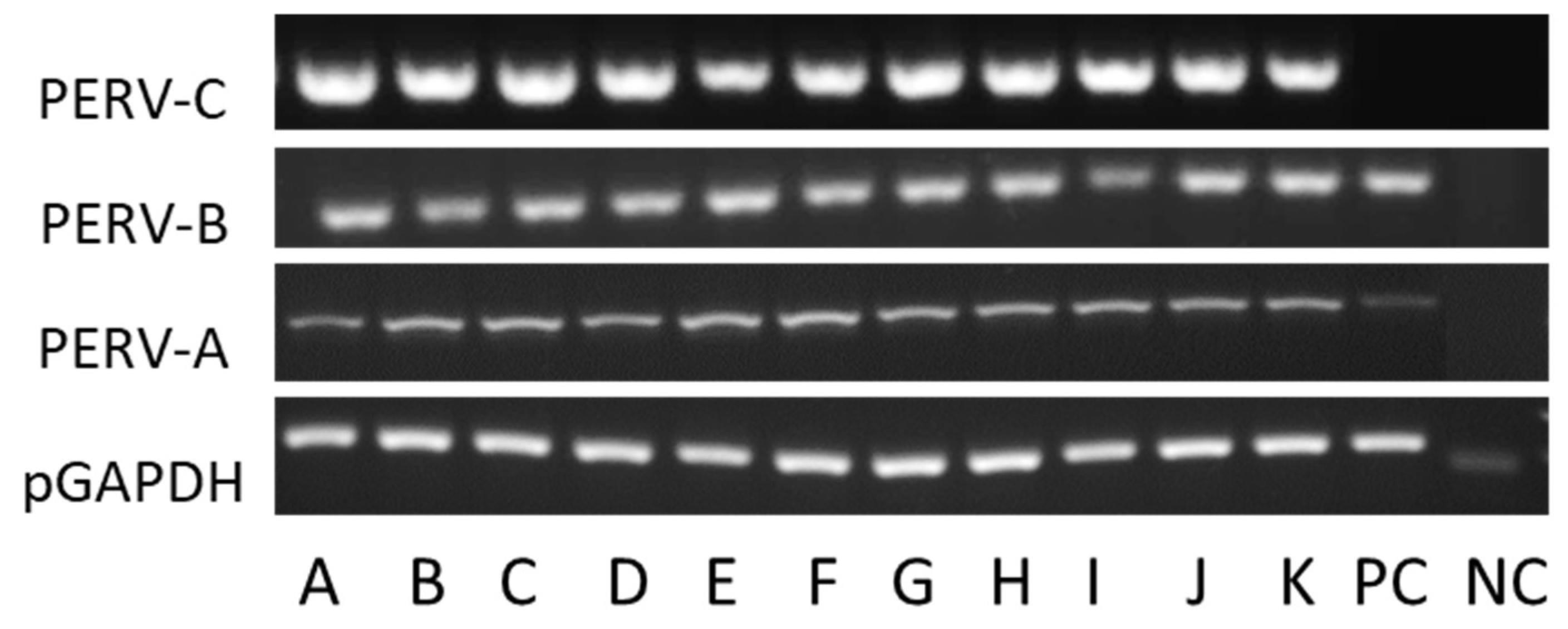
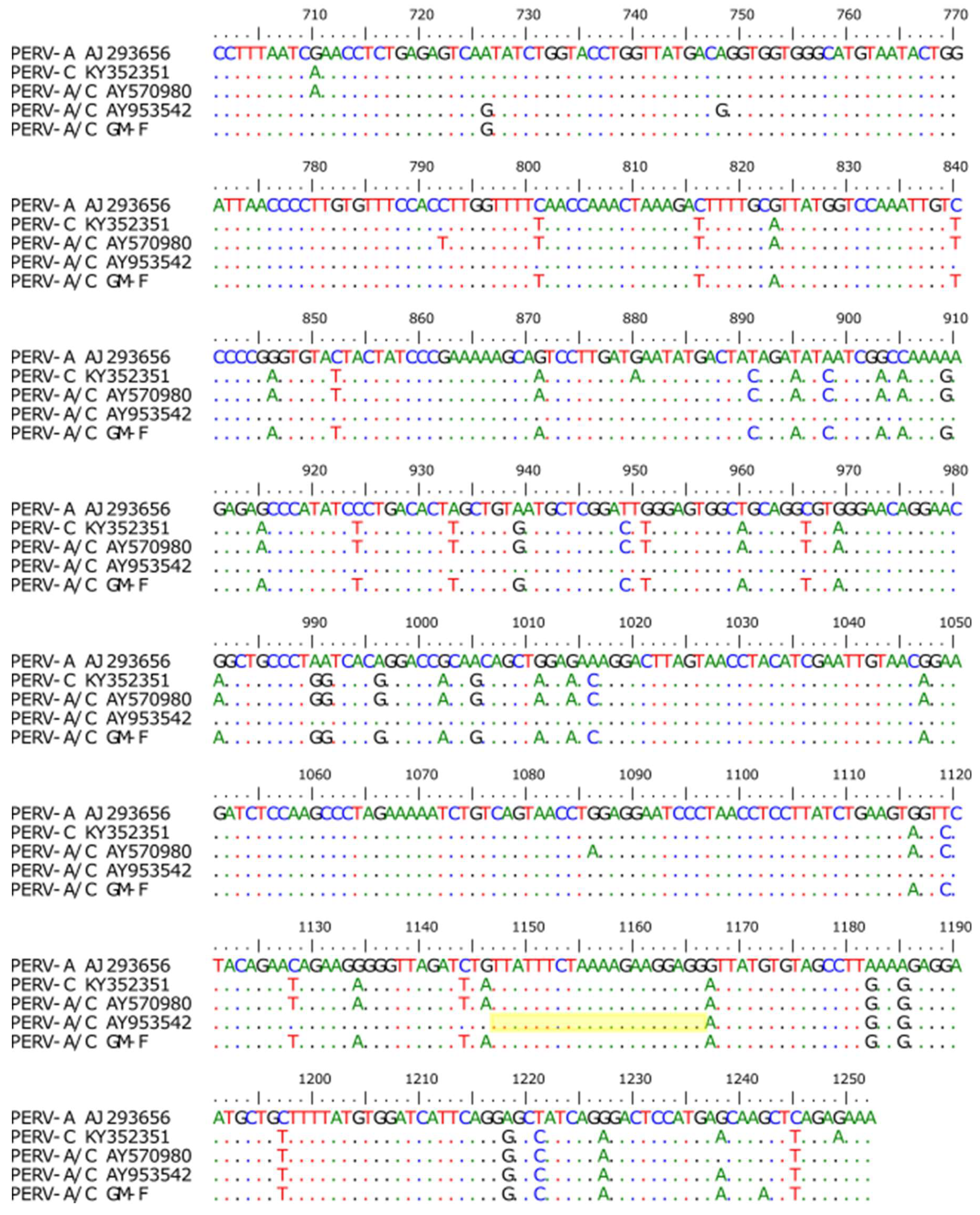
3.2. Screening for PERV-C
3.3. Screening for PERV-A/C
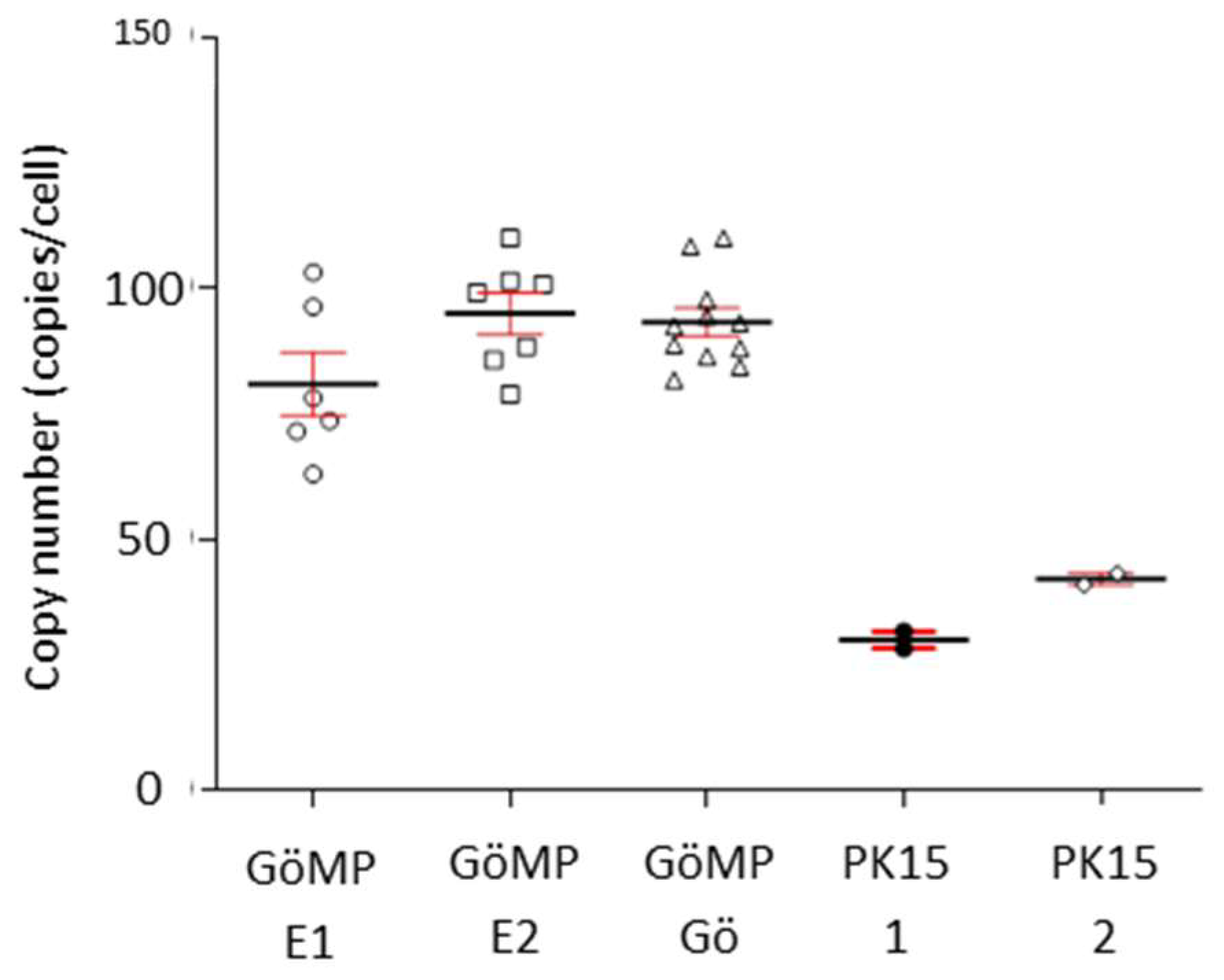
3.4. Copy Number of PERV in the Genome
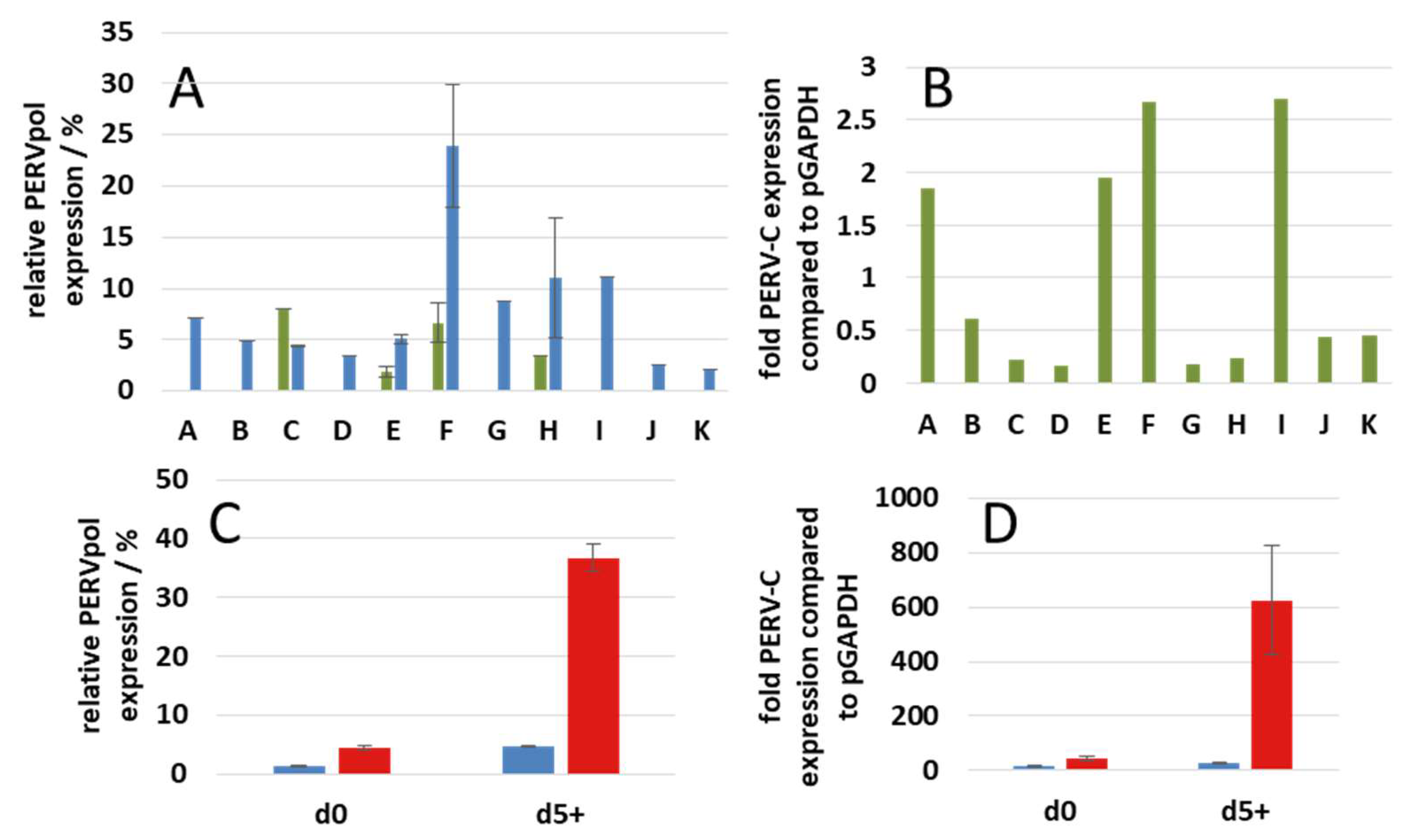
3.5. Expression of PERV
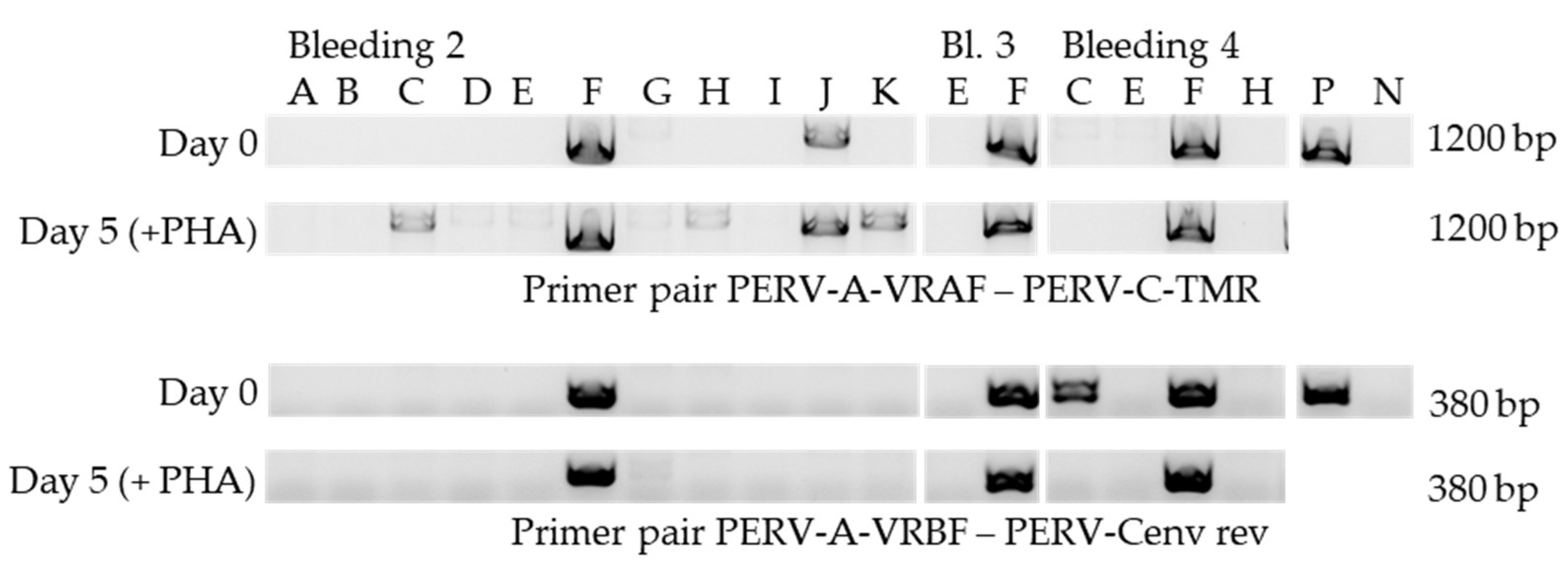
3.6. Release of Human-Tropic PERV
3.7. Screening for PCMV, HEV, PLHV, PCV2 and PCV3
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bollen, P.; Ellegaard, L. The Göttingen minipig in pharmacology and toxicology. Pharmacol. Toxicol. 1997, 80, 3–4. [Google Scholar] [CrossRef] [PubMed]
- McAnulty, P.A.; Hastings, K.L.; Ganderup, N.C. The Minipig in Biomedical Research; CRC Press/Taylor and Francis Group: Boca Raton, FL, USA, 2011. [Google Scholar]
- Available online: http//www.minipigs.dk/ (accessed on 9 July 2019).
- Simianer, H.; Köhn, F. Genetic management of the Göttingen Minipig population. J. Pharmacol. Toxicol Methods 2010, 62, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Patience, C.; Takeuchi, Y.; Weiss, R.A. Infection of human cells by an endogenous retrovirus of pigs. Nat. Med. 1997, 3, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Semaan, M.; Rotem, A.; Barkai, U.; Bornstein, S.; Denner, J. Screening pigs for xenotransplantation: Prevalence and expression of porcine endogenous retroviruses in Göttingen minipigs. Xenotransplantation 2013, 20, 148–156. [Google Scholar] [CrossRef]
- Wood, J.C.; Quinn, G.; Suling, K.M.; Oldmixon, B.A.; Van Tine, B.A.; Cina, R.; Arn, S.; Huang, C.A.; Scobie, L.; Onions, D.E.; et al. Identification of exogenous forms of human-tropic porcine endogenous retrovirus in miniature Swine. J. Virol. 2004, 78, 2494–2501. [Google Scholar] [CrossRef] [Green Version]
- Harrison, I.; Takeuchi, Y.; Bartosch, B.; Stoye, J.P. Determinants of high titer in recombinant porcine endogenous retroviruses. J. Virol. 2004, 78, 13871–13879. [Google Scholar] [CrossRef] [Green Version]
- Denner, J.; Specke, V.; Thiesen, U.; Karlas, A.; Kurth, R. Genetic alterations of the long terminal repeat of an ecotropic porcine endogenous retrovirus during passage in human cells. Virology 2003, 314, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Fiebig, U.; Fischer, K.; Bähr, A.; Runge, C.; Schnieke, A.; Wolf, E.; Denner, J. Porcine endogenous retroviruses: Quantification of the copy number in cell lines, pig breeds, and organs. Xenotransplantation 2018, 25, e12445. [Google Scholar] [CrossRef]
- Morozov, V.A.; Morozov, A.V.; Rotem, A.; Barkai, U.; Bornstein, S.; Denner, J. Extended Microbiological Characterization of Göttingen Minipigs in the Context of Xenotransplantation: Detection and Vertical Transmission of Hepatitis E Virus. PLoS ONE 2015, 10, e0139893. [Google Scholar] [CrossRef] [Green Version]
- Morozov, V.A.; Plotzki, E.; Rotem, A.; Barkai, U.; Denner, J. Extended microbiological characterization of Göttingen minipigs: Porcine cytomegalovirus and other viruses. Xenotransplantation 2016, 23, 490–496. [Google Scholar] [CrossRef]
- Webb, G.W.; Dalton, H.R. Hepatitis E: An underestimated emerging threat. Ther. Adv. Infect. Dis. 2019, 6, 2049936119837162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denner, J. Xenotransplantation and Hepatitis E virus. Xenotransplantation 2015, 22, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. Reduction of the survival time of pig xenotransplants by porcine cytomegalovirus. Virol. J. 2018, 15, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morozov, V.A.; Ludwig, S.; Ludwig, B.; Rotem, A.; Barkai, U.; Bornstein, S.R.; Denner, J. Islet cell transplantation from Göttingen minipigs to cynomolgus monkeys: Analysis of virus safety. Xenotransplantation 2016, 23, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one step real-time RT-PCR assay for rapid and sensitive detection of Hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.A.; Denner, J. New PCR diagnostic systems for the detection and quantification of the porcine cytomegalovirus (PCMV). Arch. Virol. 2016, 161, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Heinze, J.; Plotzki, E.; Denner, J. Virus Safety of Xenotransplantation: Prevalence of Porcine Cicrovirus 2 (PCV2) in Pigs. Ann. Virol. Res. 2016, 2, 1023. [Google Scholar]
- Prinz, C.; Stillfried, M.; Neubert, L.K.; Denner, J. Detection of PCV3 in German wild boars. Virol. J. 2019, 16, 25. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Güell, M.; Niu, D.; George, H.; Lesha, E.; Grishin, D.; Aach, J.; Shrock, E.; Xu, W.; Poci, J.; et al. Genome-wide inactivation of porcine endogenous retroviruses (PERVs). Science 2015, 350, 1101–1104. [Google Scholar] [CrossRef] [Green Version]
- Kaulitz, D.; Mihica, D.; Dorna, J.; Costa, M.R.; Petersen, B.; Niemann, H.; Tönjes, R.R.; Denner, J. Development of sensitive methods for detection of porcine endogenous retrovirus-C (PERV-C) in the genome of pigs. J. Virol. Methods 2011, 175, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Karlas, A.; Irgang, M.; Votteler, J.; Specke, V.; Ozel, M.; Kurth, R.; Denner, J. Characterisation of a human cell-adapted porcine endogenous retrovirus PERV-A/C. Ann. Transplant. 2010, 15, 45–54. [Google Scholar] [PubMed]
- Tacke, S.J.; Specke, V.; Denner, J. Differences in release and determination of subtype of porcine endogenous retroviruses produced by stimulated normal pig blood cells. Intervirology 2003, 46, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.A.; Wong, S.; VanBrocklin, M.; Federspiel, M.J. Extended analysis of the in vitro tropism of porcine endogenous retrovirus. J. Virol. 2000, 74, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieckhoff, B.; Kessler, B.; Jobst, D.; Kues, W.; Petersen, B.; Pfeifer, A.; Kurth, R.; Niemann, H.; Wolf, E.; Denner, J. Distribution and expression of porcine endogenous retroviruses in multi-transgenic pigs generated for xenotransplantation. Xenotransplantation 2009, 16, 64–73. [Google Scholar] [CrossRef]
- Piroozmand, A.; Yamamoto, Y.; Khamsri, B.; Fujita, M.; Uchiyama, T.; Adachi, A. Generation and characterization of APOBEC3G-positive 293T cells for HIV-1 Vif study. J. Med. Invest. 2007, 54, 154–158. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, B. Robert Koch Institute: Berlin, Germany, 2002; Unpublished work.
- Chmielewicz, B.; Goltz, M.; Franz, T.; Bauer, C.; Brema, S.; Ellerbrok, H.; Beckmann, S.; Rziha, H.J.; Lahrmann, K.H.; Romero, C.; et al. A novel porcine gammaherpesvirus. Virology 2003, 308, 317–329. [Google Scholar] [CrossRef] [Green Version]
- Mankertz, A.; Domingo, M.; Folch, J.M.; LeCann, P.; Jestin, A.; Segalés, J.; Chmielewicz, B.; Plana-Durán, J.; Soike, D. Characterisation of PCV-2 isolates from Spain, Germany and France. Virus Res. 2000, 66, 65–77. [Google Scholar] [CrossRef]
- Palinski, R.; Piñeyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A novel porcine circovirus distantly related to known circoviruses is associated with porcine dermatitis and nephropathy syndrome and reproductive failure. J. Virol. 2017, 91, e01879-16. [Google Scholar] [CrossRef] [Green Version]
- Duvigneau, J.C.; Hartl, R.T.; Groiss, S.; Gemeiner, M. Quantitative simultaneous multiplex real-time PCR for the detection of porcine cytokines. J. Immunol. Methods 2005, 306, 16–27. [Google Scholar] [CrossRef]
- Mueller, N.J.; Barth, R.N.; Yamamoto, S.; Kitamura, H.; Patience, C.; Yamada, K.; Cooper, D.K.; Sachs, D.H.; Kaur, A.; Fishman, J.A. Activation of cytomegalovirus in pig-to-primate organ xenotransplantation. J. Virol. 2002, 76, 4734–4740. [Google Scholar] [CrossRef] [Green Version]
- Dettmers, A. Die Zucht eines neuen Versuchstieres, des Miniaturschweines in Amerika. Tierzüchtung und Züchtungsbiologie 1956, 68, 37–41. [Google Scholar] [CrossRef]
- Glodek, P.; Oldigs, B. Das Göttinger Miniaturschwein; Paul Parey Verlag: Berlin/Hamburg, Germany, 1981. [Google Scholar]
- Reimer, C.; Ha, N.-T.; Sharifi,, A.R.; Weigend, S.; Geibel, J.; Simianer, H. Analyses of the breed integrity of the Goettingen Minipig using pool-sequencing. In Proceedings of the 11th World Congress of Genetics Applied to Livestock Production, Auckland, New Zealand, 11–16 February 2018. [Google Scholar]
- Bartosch, B.; Stefanidis, D.; Myers, R.; Weiss, R.; Patience, C.; Takeuchi, Y. Evidence and consequence of porcine endogenous retrovirus recombination. J. Virol. 2004, 78, 13880–13890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristiansen, Y.; Denner, J. Robert Koch Institute: Berlin, Germany, 2019; Unpublished work.
- Denner, J. Porcine endogenous retrovirus infection of human peripheral blood mononuclear cells. Xenotransplantation 2015, 22, 151–152. [Google Scholar] [CrossRef] [PubMed]
- Scobie, L.; Taylor, S.; Wood, J.C.; Suling, K.M.; Quinn, G.; Meikle, S.; Patience, C.; Schuurman, H.J.; Onions, D.E. Absence of replication-competent human-tropic porcine endogenous retroviruses in the germ line DNA of inbred miniature Swine. J. Virol. 2004, 78, 2502–2525. [Google Scholar] [CrossRef] [Green Version]
- Denner, J.; Scobie, L. Are there better assays to evaluate the risk of transmission of porcine endogenous retroviruses (PERVs) to human cells? Xenotransplantation 2019, 1, e12510. [Google Scholar] [CrossRef]
- Pal, N.; Baker, R.; Schalk, S.; Scobie, L.; Tucker, A.W.; Opriessnig, T. Detection of porcine endogenous retrovirus (PERV) viremia in diseased versus healthy US pigs by qualitative and quantitative real-time RT-PCR. Transbound Emerg. Dis. 2011, 58, 344–351. [Google Scholar] [CrossRef]
- Denner, J. Why was PERV not transmitted during preclinical and clinical xenotransplantation trials and after inoculation of animals? Retrovirology 2018, 15, 28. [Google Scholar] [CrossRef] [Green Version]
- Wynyard, S.; Nathu, D.; Garkavenko, O.; Denner, J.; Elliott, R. Microbiological safety of the first clinical pig islet xenotransplantation trial in New Zealand. Xenotransplantation 2014, 21, 309–323. [Google Scholar] [CrossRef]
- Morozov, V.A.; Wynyard, S.; Matsumoto, S.; Abalovich, A.; Denner, J.; Elliott, R. No PERV transmission during a clinical trial of pig islet cell transplantation. Virus Res. 2017, 227, 34–40. [Google Scholar] [CrossRef]
- Denner, J.; Scobie, L.; Schuurman, H.J. Is it currently possible to evaluate the risk posed by PERVs for clinical xenotransplantation? Xenotransplantation 2018, 25, e12403. [Google Scholar] [CrossRef] [Green Version]
- Karlas, A.; Kurth, R.; Denner, J. Inhibition of porcine endogenous retroviruses by RNA interference: Increasing the safety of xenotransplantation. Virology 2004, 325, 18–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyagawa, S.; Nakatsu, S.; Nakagawa, T.; Kondo, A.; Matsunami, K.; Hazama, K.; Yamada, J.; Tomonaga, K.; Miyazawa, T.; Shirakura, R. Prevention of PERV infections in pig to human xenotransplantation by the RNA interference silences gene. J. Biochem. 2005, 137, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Dieckhoff, B.; Karlas, A.; Hofmann, A.; Kues, W.A.; Petersen, B.; Pfeifer, A.; Niemann, H.; Kurth, R.; Denner, J. Inhibition of porcine endogenous retroviruses (PERVs) in primary porcine cells by RNA interference using lentiviral vectors. Arch. Virol. 2007, 152, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Dieckhoff, B.; Petersen, B.; Kues, W.A.; Kurth, R.; Niemann, H.; Denner, J. Knockdown of porcine endogenous retrovirus (PERV) expression by PERV-specific shRNA in transgenic pigs. Xenotransplantation 2008, 15, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Ramsoondar, J.; Vaught, T.; Ball, S.; Mendicino, M.; Monahan, J.; Jobst, P.; Vance, A.; Duncan, J.; Wells, K.; Ayares, D. Production of transgenic pigs that express porcine endogenous retrovirus small interfering RNAs. Xenotransplantation 2009, 16, 164–180. [Google Scholar] [CrossRef]
- Semaan, M.; Kaulitz, D.; Petersen, B.; Niemann, H.; Denner, J. Long-term effects of PERV-specific RNA interference in transgenic pigs. Xenotransplantation 2012, 19, 112–121. [Google Scholar] [CrossRef]
- Fiebig, U.; Stephan, O.; Kurth, R.; Denner, J. Neutralizing antibodies against conserved domains of p15E of porcine endogenous retroviruses: Basis for a vaccine for xenotransplantation? Virology 2003, 307, 406–413. [Google Scholar] [CrossRef] [Green Version]
- Kaulitz, D.; Fiebig, U.; Eschricht, M.; Wurzbacher, C.; Kurth, R.; Denner, J. Generation of neutralising antibodies against porcine endogenous retroviruses (PERVs). Virology 2011, 411, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Denner, J.; Mihica, D.; Kaulitz, D.; Schmidt, C.M. Increased titers of neutralizing antibodies after immunization with both envelope proteins of the porcine endogenous retroviruses (PERVs). Virol. J. 2012, 9, 260. [Google Scholar] [CrossRef] [Green Version]
- Waechter, A.; Denner, J. Novel neutralising antibodies targeting the N-terminal helical region of the transmembrane envelope protein p15E of the porcine endogenous retrovirus (PERV). Immunol. Res. 2014, 58, 9–19. [Google Scholar] [CrossRef]
- Denner, J. Can Antiretroviral Drugs Be Used to Treat Porcine Endogenous Retrovirus (PERV) Infection after Xenotransplantation? Viruses 2017, 9, E213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semaan, M.; Ivanusic, D.; Denner, J. Cytotoxic Effects during Knock Out of Multiple Porcine Endogenous Retrovirus (PERV) Sequences in the Pig Genome by Zinc Finger Nucleases (ZFN). PLoS ONE 2015, 10, e0122059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, D.; Wei, H.J.; Lin, L.; George, H.; Wang, T.; Lee, I.H.; Zhao, H.Y.; Wang, Y.; Kan, Y.; Shrock, E.; et al. Inactivation of porcine endogenous retrovirus in pigs using CRISPR-Cas9. Science 2017, 357, 1303–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denner, J.; Tönjes, R.R.; Takeuchi, Y.; Fishman, J.; Scobie, L. First update of the International Xenotransplantation Association consensus statement on conditions for undertaking clinical trials of porcine islet products in type 1 diabetes--Chapter 5: Recipient monitoring and response plan for preventing disease transmission. Xenotransplantation 2016, 23, 53–59. [Google Scholar] [PubMed]
- Denner, J. Advances in organ transplant from pigs. Science 2017, 357, 1238–1239. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. Paving the Path toward Porcine Organs for Transplantation. N. Engl. J. Med. 2017, 377, 1891–1893. [Google Scholar] [CrossRef]
- Scobie, L.; Denner, J.; Schuurman, H.J. Inactivation of porcine endogenous retrovirus in pigs using CRISPR-Cas9, editorial commentary. Xenotransplantation 2017, 24. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Sex | Date of Birth | Remarks | Virus | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PCMV | PERV | PCV2 | PCV3 | PLHV-1 | PLHV-2 | PLHV-3 | HEV | |||||||
| Real-Time PCR | PERV-C Real-Time PCR | PERV-C EnvC2 PCR | PERVpol PCR | PCR | Real-Time PCR | PCR | PCR | PCR | ELISA | Real-Time PCR | ||||
| A | M | 8 June 2014 | Father (sire) | - | + | + | + | - | - | - | - | - | - | - |
| B | F | 14 May 2015 | - | + | + | + | - | - | - | - | - | - | - | |
| C | F | 17 October 2015 | Mother (dam) | - | + | + | + | - | - | - | - | - | - | - |
| D | F | 17 October 2015 | - | + | + | + | - | - | - | - | - | - | - | |
| E | F | 29 May 2016 | - | + | + | + | - | - | - | - | - | - | - | |
| F | F | 14 October 2026 | - | + | + | + | - | - | - | - | - | - | - | |
| G | F | 7 July 2017 | Daughter (female offspring) | - | + | + | + | - | - | - | - | - | - | - |
| H | M | 25 October 2017 | - | + | + | + | - | - | - | - | - | - | - | |
| I | M | 14. January 2018 | - | + | + | + | - | - | - | - | - | - | - | |
| J | F | 31 May 2018 | - | + | + | + | - | - | - | - | - | - | - | |
| K | F | 23 June2018 | - | + | + | + | - | - | - | - | - | - | - | |
| Primers Used | Sequence 5′-3′ | Nucleotide Position | Accession Number | Reference |
|---|---|---|---|---|
| Primers used for PCR | ||||
| PLHV-1,-2 (747) fw PLHV-1,-2 (747) rev | CAYGGTAGTATTTATTCAGACA GATATCCTGGTACATTGGAAAG | 21,146–21,167 21,488–21,467 | AY170317.1 | Ehlers B., 2002 [28] |
| PLHV-3 (905) fw PLHV-3 (905) rev | ACAAGAGCCTTAGGGTTCCAAACT GTGTCCAGTGTTGTAATGGATGCC | 13,472–13,495 13,727–13,704 | AY170316.1 | Chmielewicz et al., 2003 [29] |
| PCV2 fw (F66) PCV2 rev (B67) | GGTTTGTAGCCTCAGCCAAAGC GCACCTTCGGATATACTGTCAAGG | 567–546 152–175 | KT868491.1 | Mankertz et al., 2000 [30] |
| PERV env C.2 fw PERV env C.2 rev | GATTAGAACTGGAAGCCCCAAGTGCTCT TCTGATCCAGAAGTTATGTTAGAGGATGGT | 9362–9389 9649–9620 | AM229312 | Dieckhoff et al., 2009 [26] |
| Primers and probes used to detect PERV-A/C | ||||
| PERV-A-VRBF PERV-C-TMR PERV-C rev | CCTACCAGTTATAATCAATTTAATTATGGC C TCAAACCACCCTTGAGTAGTTTCC TATGTTAGAGGATGGTCCTGGTC | Wood et al., 2004 [7] Dieckhoff et al., 2009 [26] | ||
| Primers and probes used for real-time PCR | ||||
| PCV3 fw PCV3 rev PCV3 probe | AGTGCTCCCCATTGAACG ACACAGCCGTTACTTCAC FAM-ACCCCATGG-Zen-CTCAACACATATGACC-BHQ | 1427–1444 1561–1544 1473–1449 | KT869077 | Palinski et al., 2016 [31] |
| pGAPDH fw pGAPDH rev pGAPDH probe | GATCGAGTTGGGGCTGTGACT ACATGGCCTCCAAGGAGTAAGAHEX-CCACCAACCCCAGCAAGAGCACGC-BHQ | 1083–1104 1188–1168 1114–1137 | NM_001206359.1 | Duvigneau et al., 2005 [32] |
| HEV fw HEV rev HEV probe | GGTGGTTTCTGGGGTGAC AGGGGTTGGTTGGATGAA FAM-TGATTCTCAGCCCTTCGC-BHQ | 5261–5278 5330–5313 5284–5301 | M73218. | Jothikumar et al., 2006 [17] |
| PCMV fw PCMV rev PCMV probe | ACTTCGTCGCAGCTCATCTGA GTTCTGGGATTCCGAGGTTG FAM-CAGGGCGGCGGTCGAGCTC-BHQ | Mueller et al., 2002 [33] | ||
| PERV pol fw PERV pol rev PERV pol probe | CGACTGCCCCAAGGGTTCAA TCTCTCCTGCAAATCTGGGCC FAM-CACGTACTGGAGGAGGGTCACCTG-BHQ | 3568–3587 3803–3783 3655–3678 | HM159246 | Yang et al., 2015 [21] |
| Primers used to detect spliced mRNA | ||||
| P1 P2 P3 | TGCTGTTTGCATCAAGACCGC ACAGACACTCAGAACAGAGAC ATGGAGGCGAAGCTTAAGGGGA | Karlas et al. [23] | ||
| Primers and probes used for dd PCR | ||||
| pGAPDH fw pGAPDH rev pGAPDH probe | TTCACTCCGACCTTCACCA CCGCGATCTAATGTTCTCTTTC HEX-CAGCCGCGTCCCTGAGACAC-BHQ | 3951–3970 4022–4001 3991–3972 bp | 396823 | Yang et al., 2015 [21] |
| ID | PERV Expression | Release of Virus 3 | ||||||
|---|---|---|---|---|---|---|---|---|
| Non-Stimulated, Non-Cultured | Non-Stimulated, Cultured | Stimulated, Cultured | ||||||
| FL 1 | S 2 | FL 1 | S 2 | FL 1 | S 2 | |||
| A | + | - | + | - | + | - | - | |
| B | + | - | + | + | + | + | - | |
| C | + | - | + | + | -+ | + | - | |
| D | + | - | + | + | -+ | + | - | |
| E | + | - | + | + | + | + | - | |
| F | + | - | + | + | + | + | + | |
| G | + | - | + | + | + | + | - | |
| H | + | - | + | + | + | + | - | |
| I | + | - | + | + | + | + | - | |
| J | + | - | -+ | + | + | + | - | |
| K | + | - | + | + | + | + | - | |
| Virus | Ellegaard 1 | University Göttingen |
|---|---|---|
| PERV-A, PERV-B | 40/40 (100%) | 11/11 (100%) |
| PERV-C | 28/28 (100%) | 11/11 (100%) |
| HEV | 9/40 (22.5%) | 0/11 (0%) |
| PCMV | 10/22 (45%) | 0/11 (0%) |
| PLHV-1 | 1/10 (10%) | 0/11 (0%) |
| PLHV-2 | n.t. | 0/11 (0%) |
| PLHV-3 | n.t. | 0/11 (0%) |
| PCV2 | 3/21 (14%) | 2/11 (18%) |
| PCV3 | 0/10 (0%) | 0/10 (0%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krüger, L.; Kristiansen, Y.; Reuber, E.; Möller, L.; Laue, M.; Reimer, C.; Denner, J. A Comprehensive Strategy for Screening for Xenotransplantation-Relevant Viruses in a Second Isolated Population of Göttingen Minipigs. Viruses 2020, 12, 38. https://doi.org/10.3390/v12010038
Krüger L, Kristiansen Y, Reuber E, Möller L, Laue M, Reimer C, Denner J. A Comprehensive Strategy for Screening for Xenotransplantation-Relevant Viruses in a Second Isolated Population of Göttingen Minipigs. Viruses. 2020; 12(1):38. https://doi.org/10.3390/v12010038
Chicago/Turabian StyleKrüger, Luise, Yannick Kristiansen, Emelie Reuber, Lars Möller, Michael Laue, Christian Reimer, and Joachim Denner. 2020. "A Comprehensive Strategy for Screening for Xenotransplantation-Relevant Viruses in a Second Isolated Population of Göttingen Minipigs" Viruses 12, no. 1: 38. https://doi.org/10.3390/v12010038
APA StyleKrüger, L., Kristiansen, Y., Reuber, E., Möller, L., Laue, M., Reimer, C., & Denner, J. (2020). A Comprehensive Strategy for Screening for Xenotransplantation-Relevant Viruses in a Second Isolated Population of Göttingen Minipigs. Viruses, 12(1), 38. https://doi.org/10.3390/v12010038