Characterization of a Novel Rat Hepatitis E Virus Isolated from an Asian Musk Shrew (Suncus murinus)

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. A Serum Sample from a Shrew
2.2. Inoculation of Rats and Sample Collection
2.3. Cell Culture and Virus Inoculation
2.4. Detection of Rat HEV RNA
2.5. Detection of IgG Antibodies and Rat HEV Antigen
2.6. Next-Generation Sequencing Analyses (NGS)
2.7. Phylogenetic Analyses
2.8. Examination of Alanine Aminotransferase (ALT)
3. Results
3.1. Isolation of a Rat HEV Strain from a Shrew
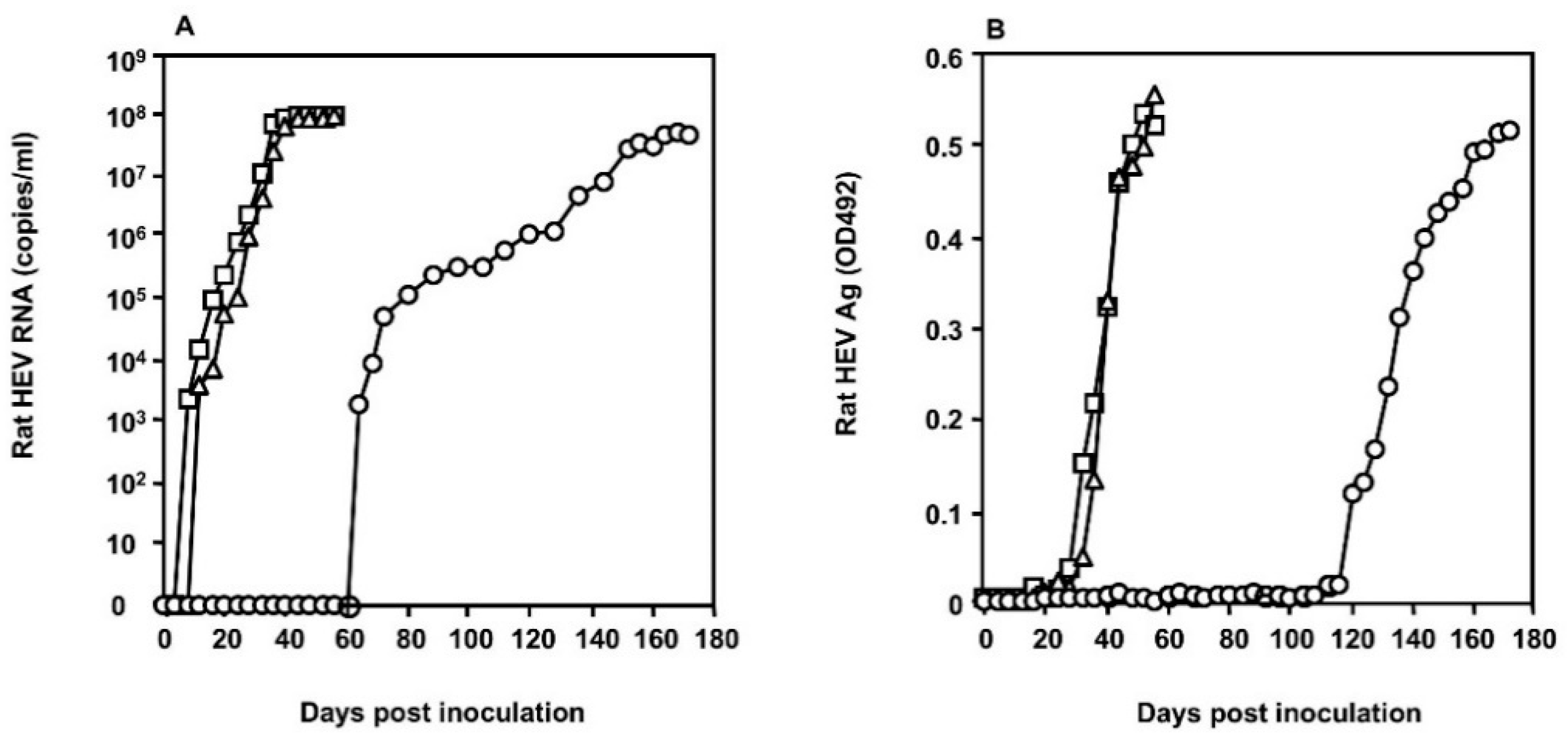
3.2. Growth of the S1129 Strain in PLC/PRF/5 Cells
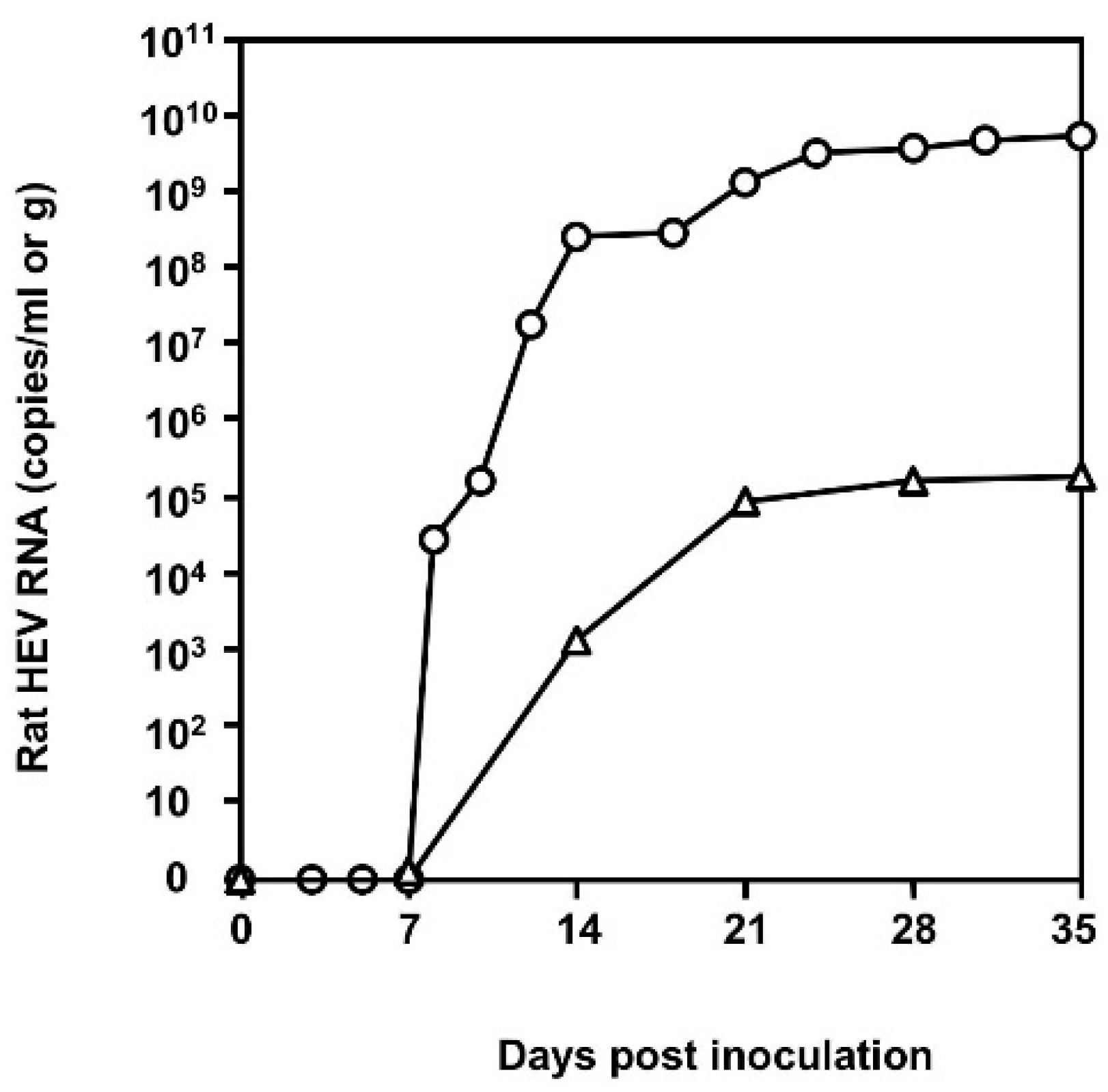
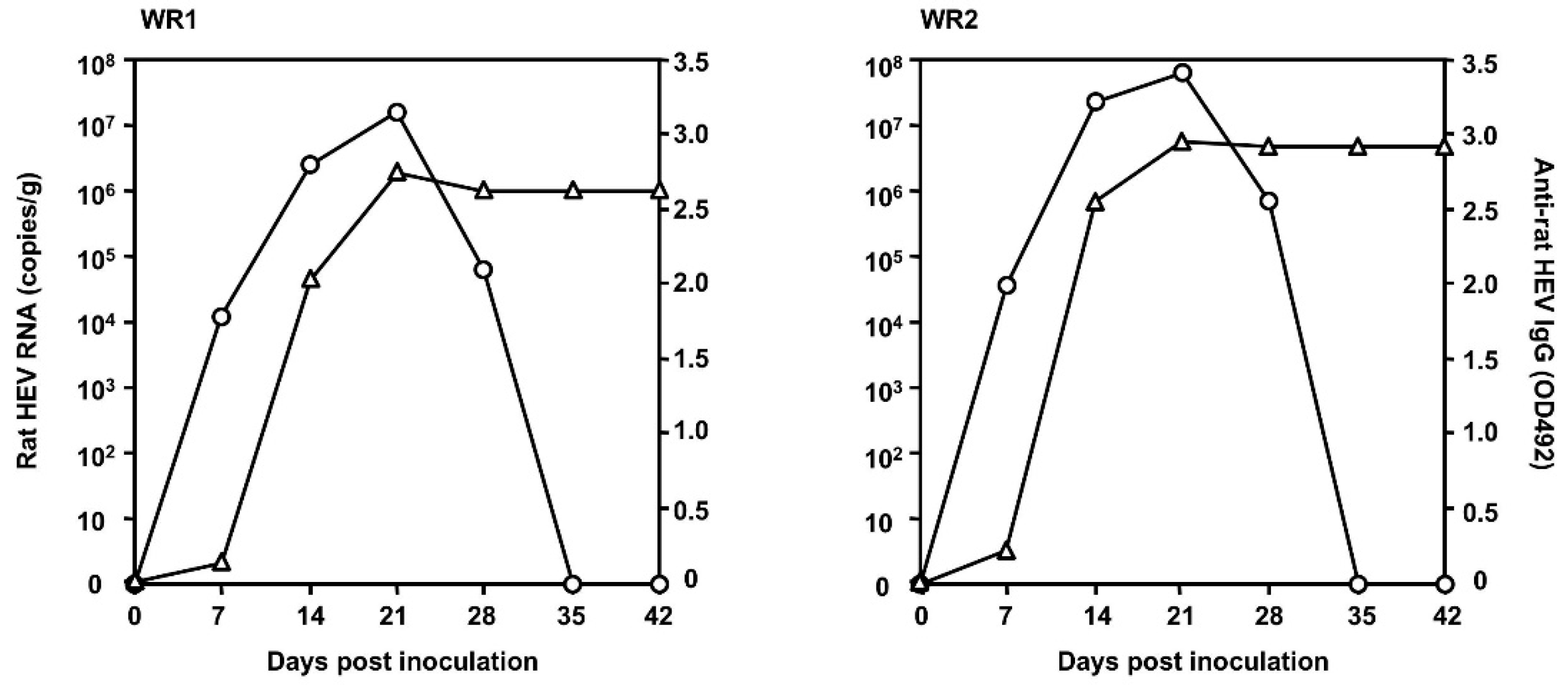
3.3. Infectivity of Rat HEV In Vivo
3.4. Characterization of Full Genome Sequences of S1129 and S1129c1
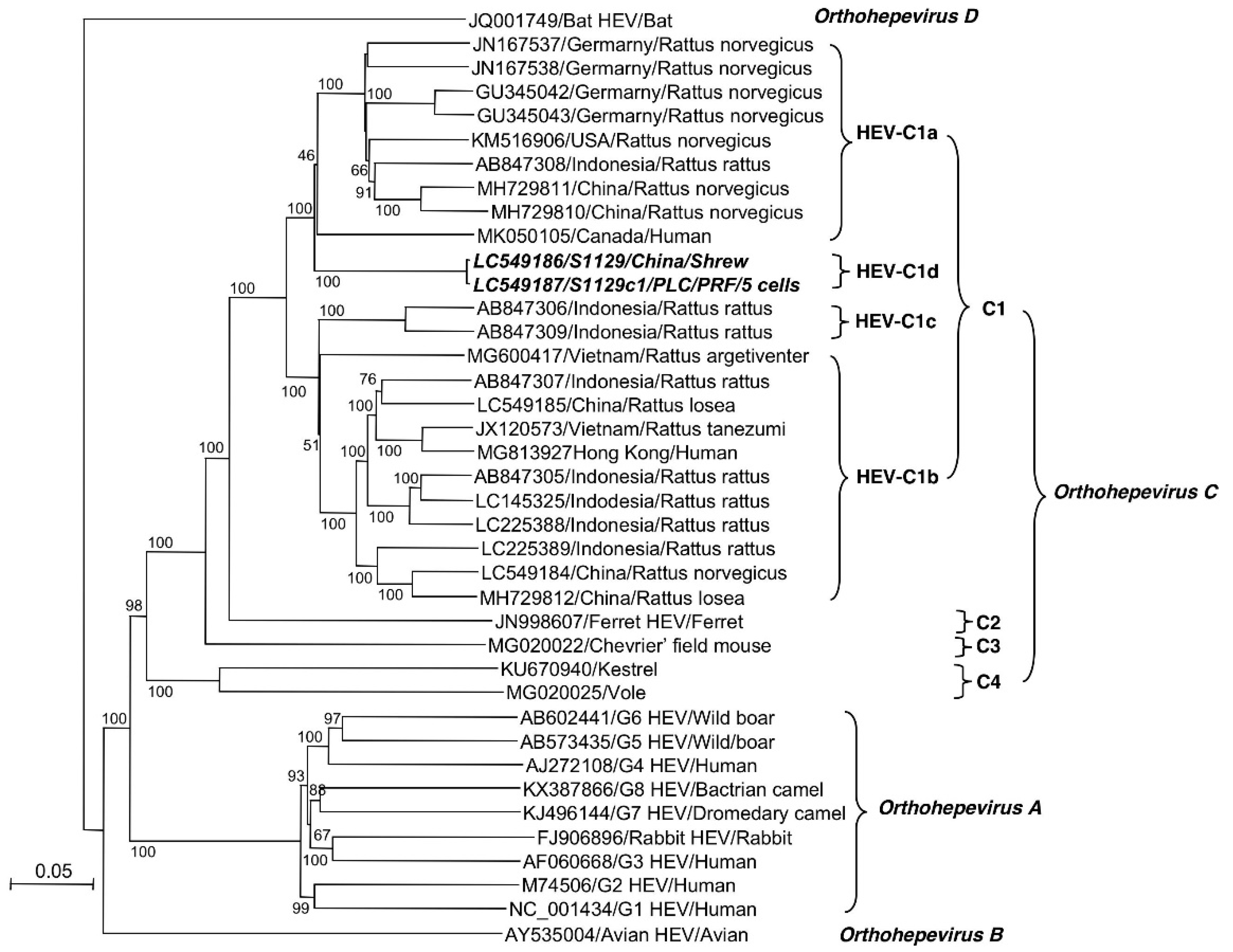
3.5. S1129 Belongs to a New Subtype of HEV-C1
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, D.B.; Simmonds, P.; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.-J.; Okamoto, H.; Van Der Poel, W.H.M.; Purdy, M.A.; members of the International Committee on the Taxonomy of Viruses Hepeviridae Study Group; et al. Consensus proposals for classification of the family Hepeviridae. J. Gen. Virol. 2014, 95, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Balayart, M.; Andjaparidze, A.; Savinskaya, S.; Ketiladze, E.; Braginsky, D.; Savinov, A.; Poleschük, V.; Balayan, M. Evidence for a Virus in Non-A, Non-B Hepatitis Transmitted via the Fecal-Oral Route. Intervirology 1983, 20, 23–31. [Google Scholar] [CrossRef]
- Meng, X.-J.; Purcell, R.H.; Halbur, P.G.; Lehman, J.R.; Webb, D.M.; Tsareva, T.S.; Haynes, J.S.; Thacker, B.J.; Emerson, S.U. A novel virus in swine is closely related to the human hepatitis E virus. Proc. Natl. Acad. Sci. USA 1997, 94, 9860–9865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Ling, R.; Erker, J.C.; Zhang, H.; Li, H.; Desai, S.; Mushahwar, I.K.; Harrison, T.J. A divergent genotype of hepatitis E virus in Chinese patients with acute hepatitis. J. Gen. Virol. 1999, 80, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Ma, Z.; Harrison, T.J.; Feng, R.; Zhang, C.; Qiao, Z.; Fan, J.; Ma, H.; Li, M.; Song, A.; et al. A novel genotype of hepatitis E virus prevalent among farmed rabbits in China. J. Med. Virol. 2009, 81, 1371–1379. [Google Scholar] [CrossRef]
- Takahashi, K.; Terada, S.; Kokuryu, H.; Arai, M.; Mishiro, S. A wild boar-derived hepatitis E virus isolate presumably representing ao far unidentified “genotype 5”. Kanzo 2010, 51, 536–538. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, M.; Nishizawa, T.; Sato, H.; Sato, Y.; Jirintai; Nagashima, S.; Okamoto, H. Analysis of the full-length genome of a hepatitis E virus isolate obtained from a wild boar in Japan that is classifiable into a novel genotype. J. Gen. Virol. 2011, 92, 902–908. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Teng, J.L.; Tsang, A.K.L.; Joseph, M.; Wong, E.Y.; Tang, Y.; Sivakumar, S.; Xie, J.; Bai, R.; et al. New Hepatitis E Virus Genotype in Camels, the Middle East. Emerg. Infect. Dis. 2014, 20, 1044–1048. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Teng, J.L.; Cao, K.-Y.; Wernery, U.; Schountz, T.; Chiu, T.H.; Tsang, A.K.; Wong, P.-C.; Wong, E.Y.; et al. New Hepatitis E Virus Genotype in Bactrian Camels, Xinjiang, China, 2013. Emerg. Infect. Dis. 2016, 22, 2219–2221. [Google Scholar] [CrossRef]
- Huang, F.F.; Sun, Z.F.; Emerson, S.U.; Purcell, R.H.; Shivaprasad, H.L.; Pierson, F.W.; Toth, T.E.; Meng, X.-J. Determination and analysis of the complete genomic sequence of avian hepatitis E virus (avian HEV) and attempts to infect rhesus monkeys with avian HEV. J. Gen. Virol. 2004, 85, 1609–1618. [Google Scholar] [CrossRef]
- Drexler, J.F.; Seelen, A.; Corman, V.M.; Tateno, A.F.; Cottontail, V.; Zerbinati, R.M.; Gloza-Rausch, F.; Klose, S.M.; Adu-Sarkodie, Y.; Oppong, S.K.; et al. Bats Worldwide Carry Hepatitis E Virus-Related Viruses That Form a Putative Novel Genus within the Family Hepeviridae. J. Virol. 2012, 86, 9134–9147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johne, R.; Heckel, G.; Plenge-Bönig, A.; Kindler, E.; Maresch, C.; Reetz, J.; Schielke, A.; Ulrich, R.G. Novel Hepatitis E Virus Genotype in Norway Rats, Germany. Emerg. Infect. Dis. 2010, 16, 1452–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raj, V.S.; Smits, S.L.; Pas, S.D.; Provacia, L.B.; Moorman-Roest, H.; Osterhaus, A.; Haagmans, B.L. Novel Hepatitis E Virus in Ferrets, the Netherlands. Emerg. Infect. Dis. 2012, 18, 1369–1370. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Boros, Á.; Mátics, R.; Kapusinszky, B.; Delwart, E.; Pankovics, P. Divergent hepatitis E virus in birds of prey, common kestrel (Falco tinnunculus) and red-footed falcon (F. vespertinus), Hungary. Infect. Genet. Evol. 2016, 43, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Li, W.; Zhou, J.-H.; Li, B.; Zhang, W.; Yang, W.-H.; Pan, H.; Wang, L.-X.; Bock, C.-T.; Shi, Z.; et al. Chevrier’s Field Mouse (Apodemus chevrieri) and Père David’s Vole (Eothenomys melanogaster) in China Carry Orthohepeviruses that form Two Putative Novel Genotypes Within the Species Orthohepevirus C. Virol. Sin. 2018, 33, 44–58. [Google Scholar] [CrossRef]
- Sridhar, S.; Yip, C.C.; Wu, S.; Cai, J.; Zhang, A.J.-X.; Leung, K.-H.; Chung, T.W.; Chan, J.F.-W.; Chan, W.-M.; Teng, J.L.; et al. Rat Hepatitis E Virus as Cause of Persistent Hepatitis after Liver Transplant. Emerg. Infect. Dis. 2018, 24, 2241–2250. [Google Scholar] [CrossRef] [Green Version]
- Andonov, A.; Robbins, M.; Borlang, J.; Cao, J.; Hatchette, T.; Stueck, A.; Deschambault, Y.; Murnaghan, K.; Varga, J.; Johnston, L. Rat Hepatitis E Virus Linked to Severe Acute Hepatitis in an Immunocompetent Patient. J. Infect. Dis. 2019, 220, 951–955. [Google Scholar] [CrossRef]
- Sridhar, S.; Yip, C.C.; Wu, S.; Chew, N.F.; Leung, K.; Chan, J.F.; Zhao, P.S.; Chan, W.; Poon, R.W.; Tsoi, H.; et al. Transmission of rat hepatitis E virus infection to humans in Hong Kong: A clinical and epidemiological analysis. Hepatology 2020. [Google Scholar] [CrossRef]
- Ryll, R.; Bernstein, S.; Heuser, E.; Schlegel, M.; Dremsek, P.; Zumpe, M.; Wolf, S.; Pépin, M.; Bajomi, D.; Müller, G.; et al. Detection of rat hepatitis E virus in wild Norway rats (Rattus norvegicus) and Black rats (Rattus rattus) from 11 European countries. Vet. Microbiol. 2017, 208, 58–68. [Google Scholar] [CrossRef]
- Li, W.; Guan, D.; Su, J.; Takeda, N.; Wakita, T.; Li, T.; Ke, C. High prevalence of rat hepatitis E virus in wild rats in China. Vet. Microbiol. 2013, 165, 275–280. [Google Scholar] [CrossRef]
- Purcell, R.H.; Engle, R.E.; Rood, M.P.; Kabrane-Lazizi, Y.; Nguyen, H.T.; Govindarajan, S.; Claire, M.S.; Emerson, S.U. Hepatitis E Virus in Rats, Los Angeles, California, USA. Emerg. Infect. Dis. 2011, 17, 2216–2222. [Google Scholar] [CrossRef] [PubMed]
- Mulyanto; Suparyatmo, J.B.; Andayani, I.G.A.S.; Khalid; Takahashi, M.; Ohnishi, H.; Jirintai, S.; Nagashima, S.; Nishizawa, T.; Okamoto, H. Marked genomic heterogeneity of rat hepatitis E virus strains in Indonesia demonstrated on a full-length genome analysis. Virus Res. 2014, 179, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Primadharsini, P.P.; Mulyanto; Wibawa, I.D.N.; Anggoro, J.; Nishizawa, T.; Takahashi, M.; Jirintai, S.; Okamoto, H. The identification and characterization of novel rat hepatitis E virus strains in Bali and Sumbawa, Indonesia. Arch. Virol. 2018, 163, 1345–1349. [Google Scholar] [CrossRef] [PubMed]
- Li, T.-C.; Ami, Y.; Suzaki, Y.; Yasuda, S.P.; Yoshimatsu, K.; Arikawa, J.; Takeda, N.; Takaji, W. Characterization of Full Genome of Rat Hepatitis E Virus Strain from Vietnam. Emerg. Infect. Dis. 2013, 19, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Johne, R.; Dremsek, P.; Reetz, J.; Heckel, G.; Hess, M.; Ulrich, R.G. Hepeviridae: An expanding family of vertebrate viruses. Infect. Genet. Evol. 2014, 27, 212–229. [Google Scholar] [CrossRef]
- Guan, D.; Li, W.; Su, J.; Fang, L.; Takeda, N.; Wakita, T.; Li, T.-C.; Ke, C. Asian Musk Shrew as a Reservoir of Rat Hepatitis E Virus, China. Emerg. Infect. Dis. 2013, 19, 1341–1343. [Google Scholar] [CrossRef]
- Wang, B.; Cai, C.-L.; Li, B.; Zhang, W.; Zhu, Y.; Chen, W.-H.; Zhuo, F.; Shi, Z.-L.; Yang, X. Detection and characterization of three zoonotic viruses in wild rodents and shrews from Shenzhen city, China. Virol. Sin. 2017, 32, 290–297. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Wen, Y.; Xiong, Y.-Q.; Zhang, M.; Cheng, M.-J.; Chen, Q. The prevalence and genomic characteristics of hepatitis E virus in murine rodents and house shrews from several regions in China. BMC Vet. Res. 2018, 14, 414. [Google Scholar] [CrossRef]
- Johne, R.; Plenge-Bönig, A.; Hess, M.; Ulrich, R.G.; Reetz, J.; Schielke, A. Detection of a novel hepatitis E-like virus in faeces of wild rats using a nested broad-spectrum RT-PCR. J. Gen. Virol. 2009, 91, 750–758. [Google Scholar] [CrossRef]
- Li, T.-C.; Yang, T.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Ishii, K.; Haga, K.; Nakamura, T.; Ochiai, S.; Takaji, W.; et al. Construction and characterization of an infectious cDNA clone of rat hepatitis E virus. J. Gen. Virol. 2015, 96, 1320–1327. [Google Scholar] [CrossRef]
- Li, T.-C.; Yoshimatsu, K.; Yasuda, S.P.; Arikawa, J.; Koma, T.; Kataoka, M.; Ami, Y.; Suzaki, Y.; Mai, L.T.Q.; Hoa, N.T.; et al. Characterization of self-assembled virus-like particles of rat hepatitis E virus generated by recombinant baculoviruses. J. Gen. Virol. 2011, 92, 2830–2837. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Li, T.-C.; Zhou, X.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Nakamura, T.; Takeda, N.; Wakita, T. Production of infectious dromedary camel hepatitis E virus by a reverse genetic system: Potential for zoonotic infection. J. Hepatol. 2016, 65, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Li, T.-C.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Yasuda, S.P.; Yoshimatsu, K.; Arikawa, J.; Takeda, N.; Wakita, T. Susceptibility of laboratory rats against genotypes 1, 3, 4, and rat hepatitis E viruses. Vet. Microbiol. 2013, 163, 54–61. [Google Scholar] [CrossRef]
- Cossaboom, C.M.; Córdoba, L.; Sanford, B.J.; Piñeyro, P.; Kenney, S.P.; Dryman, B.A.; Wang, Y.; Meng, X.-J. Cross-species infection of pigs with a novel rabbit, but not rat, strain of hepatitis E virus isolated in the United States. J. Gen. Virol. 2012, 93, 1687–1695. [Google Scholar] [CrossRef] [PubMed]
- Dremsek, P.; Wenzel, J.J.; Johne, R.; Ziller, M.; Hofmann, J.; Groschup, M.H.; Werdermann, S.; Mohn, U.; Dorn, S.; Motz, M.; et al. Seroprevalence study in forestry workers from eastern Germany using novel genotype 3- and rat hepatitis E virus-specific immunoglobulin G ELISAs. Med. Microbiol. Immunol. 2011, 201, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Krumbholz, A.; Joel, S.; Neubert, A.; Dremsek, P.; Dürrwald, R.; Johne, R.; Hlinak, A.; Walther, M.; Lange, J.; Wutzler, P.; et al. Age-related and regional differences in the prevalence of hepatitis E virus-specific antibodies in pigs in Germany. Vet. Microbiol. 2013, 167, 394–402. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nucleotide | Amino Acid | ||||||
|---|---|---|---|---|---|---|---|
| a Position | S1129 | S1129c1 | b Position | ORF1 | ORF2 | ORF3 | ORF4 |
| 346 | T | C | 107 | Val/Ala | |||
| 766 | T | C | |||||
| 957 | T | A | 316 | Phe/Tyr | |||
| 1639 | C | T | |||||
| 1671 | C | T | 534 | Thr/Ile | |||
| 1732 | T | C | |||||
| 1756 | C | T | |||||
| 1822 | T | C | |||||
| 1907 | C | T | 633 | His/Tyr | |||
| 2560 | T | C | |||||
| 2561 | T | C | 851 | Ser/Pro | |||
| 2563 | T | C | |||||
| 2740 | C | T | |||||
| 2914 | T | G | |||||
| 2958 | C | T | |||||
| 3211 | T | C | |||||
| 3649 | T | C | |||||
| 3859 | G | A | |||||
| 4144 | T | C | |||||
| 4588 | A | T | |||||
| 5115 | G | A | c 52 (56) | Arg/Gln | Gly/Arg | ||
| 5156 | C | T | 66 | Leu/Phe | |||
| 5179 | C | T | 77 | Ala/Val | |||
| 5335 | C | T | |||||
| 5552 | C | A | 198 | Leu/Met | |||
| 5634 | T | C | 225 | Val/Ala | |||
| 5661 | T | G | 234 | Val/Gly | |||
| 5869 | C | T | |||||
| 6624 | C | T | 555 | Ser/Phe | |||
| 6695 | C | T | |||||
| 6762 | C | T | 579 | Ser/Leu | |||
| 6908 | T | C | |||||
| Rat HEV S1129 (LC549186) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Nucleotides (%) | Amino Acids (%) | |||||||
| HEV (Accession No) | Species | Entire Genome | ORF1 | ORF2 | ORF3 | ORF1 | ORF2 | ORF3 |
| Bat HEV (JQ001749) | HEV-D | 46.1 | 48.5 | 55.2 | 46.0 | 50.5 | 52.8 | 50.0 |
| Rat HEV (GU345042) | HEV-C1 | 80.9 | 79.4 | 84.3 | 89.0 a | 90.1 | 95.4 | 78.4 |
| Rat HEV (GU345043) | HEV-C1 | 80.0 | 79.1 | 83.4 | 88.7 | 89.5 | 95.5 | 78.4 |
| Rat HEV (JN167537) | HEV-C1 | 80.8 | 79.8 | 83.3 | 87.7 | 90.9 | 94.6 | 75.5 |
| Rat HEV (AB847308) | HEV-C1 | 80.5 | 80.8 | 83.0 | 86.4 | 90.8 | 94.1 | 74.5 |
| Rat HEV (KM516906) | HEV-C1 | 81.8 | 81.0 | 83.5 | 88.0 | 90.6 | 94.7 | 79.4 |
| Rat HEV (AB847306) | HEV-C1 | 79.4 | 77.7 | 80.3 | 80.9 | 87.7 | 92.9 | 67.6 |
| Rat HEV (AB847309) | HEV-C1 | 78.8 | 77.7 | 79.9 | 79.5 | 86.5 | 92.4 | 66.7 |
| Rat HEV (AB847305) | HEV-C1 | 78.5 | 78.4 | 79.3 | 81.6 | 87.5 | 93.2 | 67.6 |
| Rat HEV (AB847307) | HEV-C1 | 78.3 | 78.5 | 79.4 | 82.8 | 87.1 | 93.8 | 72.5 |
| Rat HEV (JX120573) | HEV-C1 | 78.9 | 78.1 | 80.0 | 83.7 | 86.8 | 93.5 | 72.5 |
| Rat HEV (MK050105) | HEV-C1 | 80.7 | 79.7 | 82.9 | 86.6 | 90.1 | 94.6 | 75.6 |
| Ferret HEV (AB890001) | HEV-C2 | 60.8 | 56.4 | 71.1 | 63.7 | 73.9 | 79.1 | 45.6 |
| Chevrier HEV(MG020022) | HEV-C3 | 64.9 | 63.4 | 68.3 | 65.1 | 66.8 | 74.3 | 46.1 |
| Kestrel HEV (KU670940) | HEV-C4 | 54.0 | 57.2 | 62.4 | 46.9 | 54.2 | 69.1 | 29.2 |
| Genotype 1 (NC-001434) | HEV-A | 49.3 | 51.3 | 60.4 | 51.9 | 57.1 | 55.5 | 33.3 |
| Genotype 2 (M74506) | HEV-A | 49.0 | 50.5 | 58.8 | 52.6 | 57.0 | 55.2 | 30.0 |
| Genotype 3 (AF060668) | HEV-A | 47.3 | 51.0 | 60.1 | 49.4 | 57.3 | 56.8 | 26.6 |
| Genotype 4 (AJ272108) | HEV-A | 49.2 | 51.4 | 60.5 | 56.5 | 57.0 | 56.5 | 25.7 |
| Genotype 5 (AB573435) | HEV-A | 50.0 | 51.9 | 60.9 | 55.3 | 57.5 | 56.0 | 27.1 |
| Genotype 6 (AB602441) | HEV-A | 48.4 | 51.2 | 60.5 | 53.6 | 57.5 | 56.1 | 28.9 |
| Genotype 7 (KJ496143) | HEV-A | 48.2 | 50.2 | 58.5 | 50.3 | 57.5 | 56.6 | 23.7 |
| Genotype 8 (KX387866) | HEV-A | 48.3 | 48.2 | 60.5 | 51.6 | 57.6 | 55.6 | 25.0 |
| Rabbit HEV (FJ906895) | HEV-A | 48.9 | 51.1 | 58.8 | 51.4 | 51.8 | 56.1 | 27.6 |
| Avian HEV (AY535004) | HEV-B | 48.5 | 49.7 | 54.2 | 47.0 | 44.7 | 47.4 | 33.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, H.; Li, W.; Guan, D.; Su, J.; Ke, C.; Ami, Y.; Suzaki, Y.; Takeda, N.; Muramatsu, M.; Li, T.-C. Characterization of a Novel Rat Hepatitis E Virus Isolated from an Asian Musk Shrew (Suncus murinus). Viruses 2020, 12, 715. https://doi.org/10.3390/v12070715
Bai H, Li W, Guan D, Su J, Ke C, Ami Y, Suzaki Y, Takeda N, Muramatsu M, Li T-C. Characterization of a Novel Rat Hepatitis E Virus Isolated from an Asian Musk Shrew (Suncus murinus). Viruses. 2020; 12(7):715. https://doi.org/10.3390/v12070715
Chicago/Turabian StyleBai, Huimin, Wei Li, Dawei Guan, Juan Su, Changwen Ke, Yasushi Ami, Yuriko Suzaki, Naokazu Takeda, Masamichi Muramatsu, and Tian-Cheng Li. 2020. "Characterization of a Novel Rat Hepatitis E Virus Isolated from an Asian Musk Shrew (Suncus murinus)" Viruses 12, no. 7: 715. https://doi.org/10.3390/v12070715
APA StyleBai, H., Li, W., Guan, D., Su, J., Ke, C., Ami, Y., Suzaki, Y., Takeda, N., Muramatsu, M., & Li, T.-C. (2020). Characterization of a Novel Rat Hepatitis E Virus Isolated from an Asian Musk Shrew (Suncus murinus). Viruses, 12(7), 715. https://doi.org/10.3390/v12070715