West Nile Virus in Slovenia

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Trapping
2.2. Virus Detection
2.3. Case Laboratory Investigations
2.4. Retrospective Surveillance of Patients with Meningitis/Meningoencephalitis
2.5. Screening of Dog Sera
2.6. Virus Neutralization Assay
2.7. Virus Isolation
2.8. Complete Genome Sequencing
2.9. Phylogenetic Analysis
2.10. Ethics Statement
3. Results
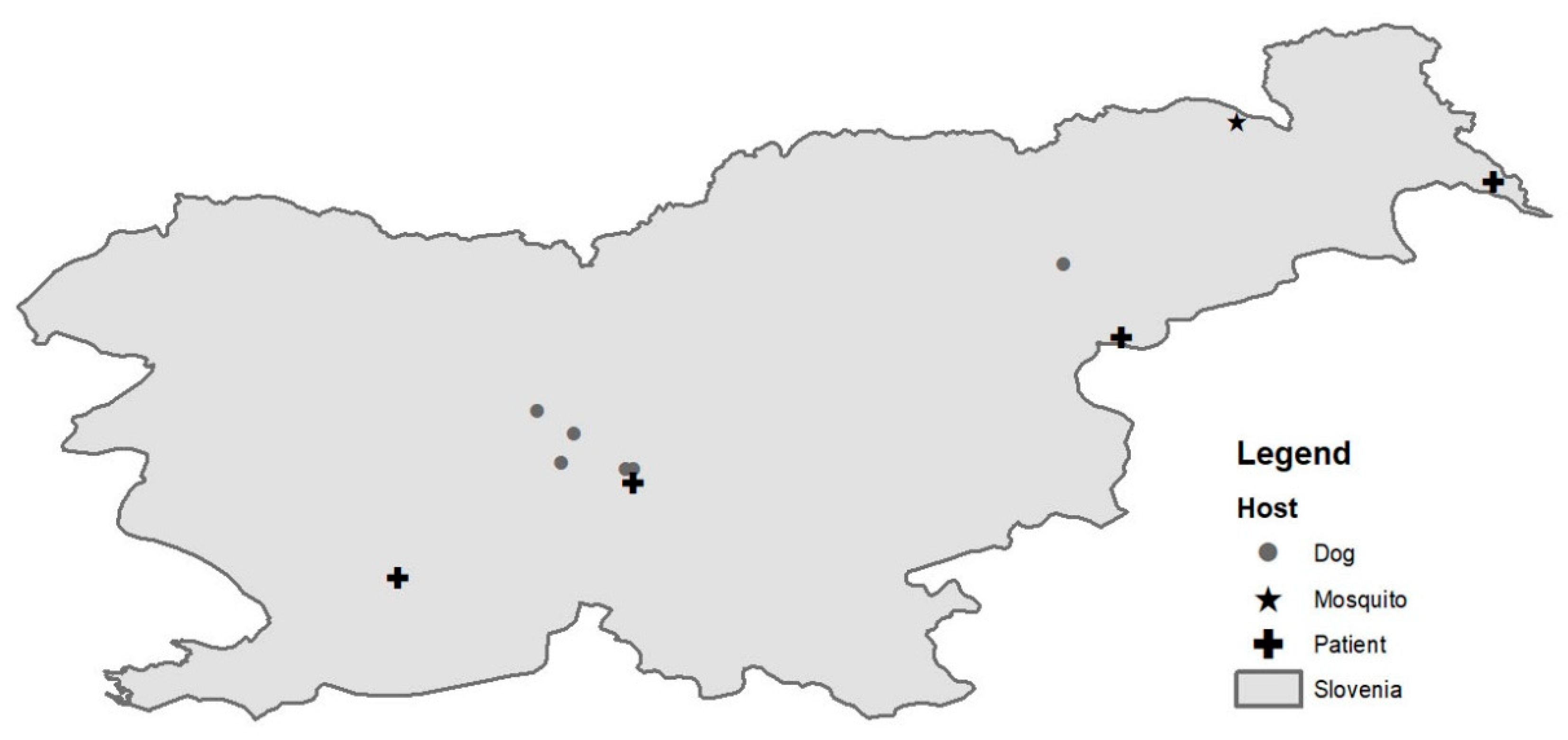
3.1. Mosquito Trapping and Virus Detection
3.2. Case Laboratory Investigations
3.3. Retrospective Surveillance of Patients with Meningitis/Meningoencephalitis
3.4. Screening of Dog Sera
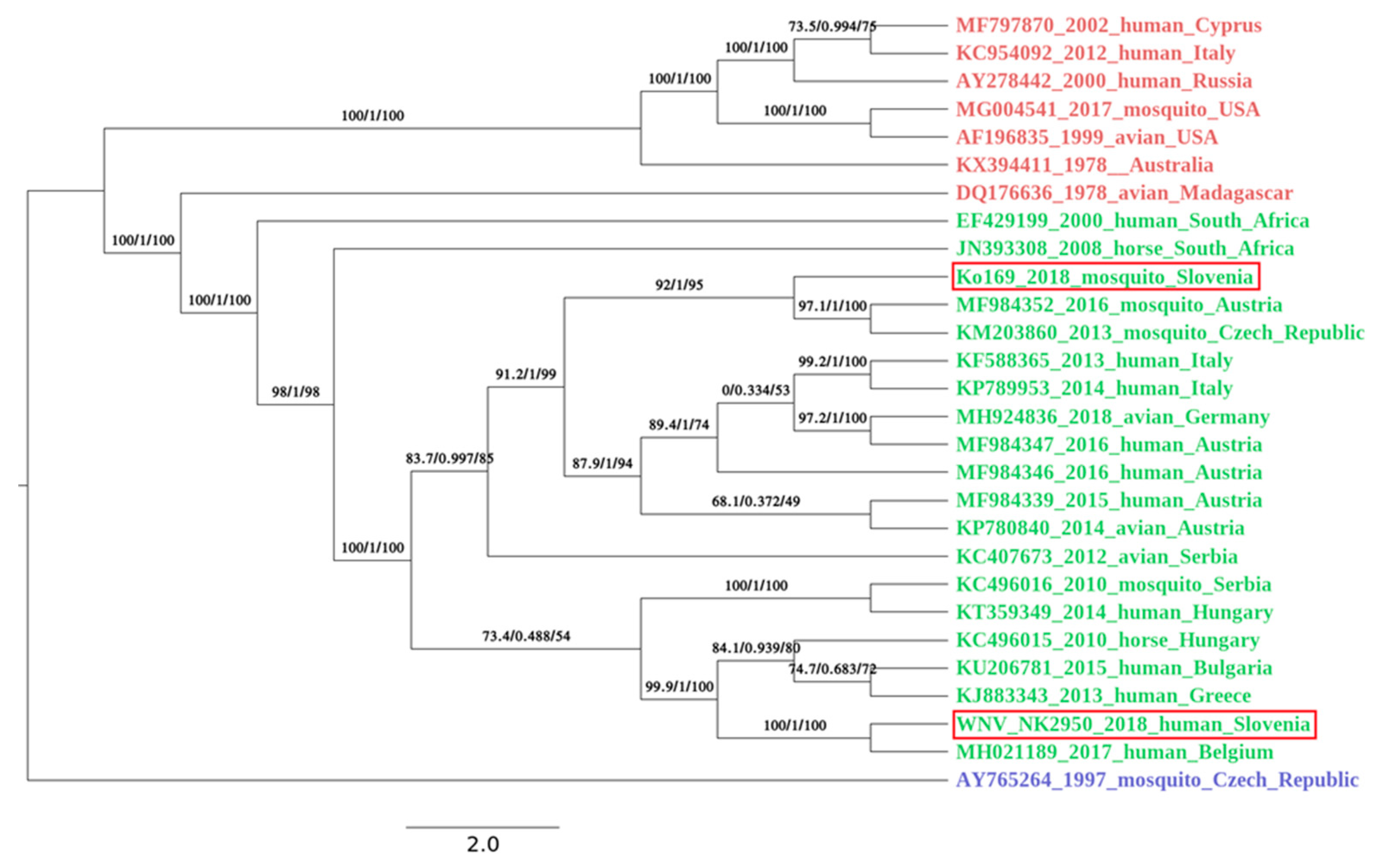
3.5. Genome Sequencing and Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hubalek, Z.; Halouzka, J. West Nile fever—A reemerging mosquito-borne viral disease in Europe. Emerg. Infect. Dis. 1999, 5, 643–650. [Google Scholar] [CrossRef]
- Hubalek, Z.; Rudolf, I.; Nowotny, N. Arboviruses pathogenic for domestic and wild animals. Adv. Virus Res. 2014, 89, 201–275. [Google Scholar] [CrossRef] [PubMed]
- Conte, A.; Candeloro, L.; Ippoliti, C.; Monaco, F.; De Massis, F.; Bruno, R.; Di Sabatino, D.; Danzetta, M.L.; Benjelloun, A.; Belkadi, B.; et al. Spatio-Temporal Identification of Areas Suitable for West Nile Disease in the Mediterranean Basin and Central Europe. PLoS ONE 2015, 10, e0146024. [Google Scholar] [CrossRef] [PubMed]
- ECDC. Weekly Updates: 2020 West Nile Virus Transmission Season. Available online: https://www.ecdc.europa.eu/en/west-nile-fever/surveillance-and-disease-data/disease-data-ecdc (accessed on 23 June 2020).
- Hernandez-Triana, L.M.; Jeffries, C.L.; Mansfield, K.L.; Carnell, G.; Fooks, A.R.; Johnson, N. Emergence of West Nile virus lineage 2 in europe: A review on the introduction and spread of a mosquito-borne disease. Front. Public Health 2014, 2, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakonyi, T.; Ivanics, E.; Erdelyi, K.; Ursu, K.; Ferenczi, E.; Weissenbock, H.; Nowotny, N. Lineage 1 and 2 strains of encephalitic West Nile virus, central Europe. Emerg. Infect. Dis. 2006, 12, 618–623. [Google Scholar] [CrossRef]
- Wodak, E.; Richter, S.; Bago, Z.; Revilla-Fernandez, S.; Weissenbock, H.; Nowotny, N.; Winter, P. Detection and molecular analysis of West Nile virus infections in birds of prey in the eastern part of Austria in 2008 and 2009. Vet. Microbiol. 2011, 149, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Kolodziejek, J.; Jungbauer, C.; Aberle, S.W.; Allerberger, F.; Bago, Z.; Camp, J.V.; Dimmel, K.; de Heus, P.; Kolodziejek, M.; Schiefer, P.; et al. Integrated analysis of human-animal-vector surveillance: West Nile virus infections in Austria, 2015-2016. Emerg. Microbes.Infect. 2018, 7, 25. [Google Scholar] [CrossRef] [Green Version]
- Nagy, A.; Mezei, E.; Nagy, O.; Bakonyi, T.; Csonka, N.; Kaposi, M.; Koroknai, A.; Szomor, K.; Rigó, Z.; Molnár, Z.; et al. Extraordinary increase in West Nile virus cases and first confirmed human Usutu virus infection in Hungary, 2018. Euro. Surveill. 2019, 24. [Google Scholar] [CrossRef] [Green Version]
- Vilibic-Cavlek, T.; Savic, V.; Sabadi, D.; Peric, L.; Barbic, L.; Klobucar, A.; Miklausic, B.; Tabain, I.; Santini, M.; Vucelja, M.; et al. Prevalence and molecular epidemiology of West Nile and Usutu virus infections in Croatia in the ’One health’ context, 2018. Transbound. Emerg. Dis. 2019, 66, 1946–1957. [Google Scholar] [CrossRef]
- Rizzo, C.; Napoli, C.; Venturi, G.; Pupella, S.; Lombardini, L.; Calistri, P.; Monaco, F.; Cagarelli, R.; Angelini, P.; Bellini, R.; et al. West Nile virus transmission: Results from the integrated surveillance system in Italy, 2008 to 2015. Euro. Surveill. 2016, 21. [Google Scholar] [CrossRef] [Green Version]
- Becker, N.; Zgomba, M.; Petric, D.; Dahl, C.; Boase, C.; Lane, J.; Kaiser, A. Mosquitoes and Their Control; Springer: New York, NY, USA, 2003; Volume 2. [Google Scholar]
- Gutsevich, A.V.; Monchadskii, A.S.; Shtakel’berg, A.A. Fauna of the U.S.S.R. Diptera. Volume 3, No. 4. Mosquitoes Family Culicidae. In The Quarterly Review of Biology; Dykhuizen, D.E., Dávalos, L.M., Thomson, J.D., Wiens, J.J., Wray, G.A., Eds.; Keter Publishing House Jerusalem Ltd.: Jerusalem, Israel, 1976; Volume 51, p. 149. [Google Scholar]
- Schaffner, F.; Angel, G.; Geoffroy, B.; Hervy, J.P.; Rhaiem, A.; Brunhes, J. The Mosquitoes of Europe—An Identification and Training Programme; [CD-ROM]; Institut de Recherche pour le Developpement: Montpellier, France, 2001. [Google Scholar]
- Linke, S.; Ellerbrok, H.; Niedrig, M.; Nitsche, A.; Pauli, G. Detection of West Nile virus lineages 1 and 2 by real-time PCR. J. Virol. Methods 2007, 146, 355–358. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narasimhan, V.; Danecek, P.; Scally, A.; Xue, Y.; Tyler-Smith, C.; Durbin, R. BCFtools/RoH: A hidden Markov model approach for detecting autozygosity from next-generation sequencing data. Bioinformatics 2016, 32, 1749–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- ECDC. Surveillance and Disease Data for West Nile Fever. Available online: https://www.ecdc.europa.eu/en/west-nile-fever/surveillance-and-disease-data (accessed on 23 June 2020).
- Minh, B.Q.; Nguyen, M.A.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Anisimova, M.; Gascuel, O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Hlad, B.; Skoberne, P.; Orozen Adamic, M.; Anko, B.; Veselic, Z.; Kraigher, H.; Novak, T.; Devetak, D.; Jurc, D.; Urbancic, M.; et al. Pregled Stanja Biotske Raznovrstnosti in Krajinske Pestrosti v Sloveniji; Ministrstvo za Okolje in Prostor Republike Slovenije, Agencija RS za okolje: Ljubljana, Slovenia, 2002.
- Račnik, J.; Trilar, T.; Slavec, B.; Marhold, C.; Jelovšek, M.; Verglas Rataj, A.; Avšič-Županc, T.; Zorman Rojs, O. Study of some pathogens in living wild passerine birds captured during autumn migration in Slovenia. Slov. Vet. Res. 2011, 48, 146–149. [Google Scholar]
- Zorman-Rojs, O.; Slavec, B.; Račnik, J.; Krapež, U.; Zadravec, M.; Dovč, A.; Hari, A.; Trilar, T.; Avšič-Županc, T. Incidence of avian influenza and West Nile in birds in Slovenia. Med. Razgl. 2012, 51, 157–164. [Google Scholar]
- Kalan, K.; Avšič-Županc, T.; Rus, K.R.; Bužan, E.; Duh, D. Mosquito-only flaviviruses, isolated from Aedes albopictus in Slovenia: Results of a pilot mosquito monitoring program. Biologia 2018, 73, 1277–1282. [Google Scholar] [CrossRef]
- Kalan, K.; Ivovic, V.; Glasnovic, P.; Buzan, E. Presence and Potential Distribution of Aedes albopictus and Aedes japonicus japonicus (Diptera: Culicidae) in Slovenia. J. Med. Entomol. 2017, 54, 1510–1518. [Google Scholar] [CrossRef]
- Kalan, K.; Susnjar, J.; Ivovic, V.; Buzan, E. First record of Aedes koreicus (Diptera, Culicidae) in Slovenia. Parasitol. Res. 2017, 116, 2355–2358. [Google Scholar] [CrossRef]
- Avsic-Zupanc, T.; Petrovec, M.; Jelovsek, M.; Strle, F. Medicinsko pomembni arbovirusi v Sloveniji. Zdr. Vestn. 1995, 64, 15–19. [Google Scholar]
- ECDC. Table. Transmission of West Nile fever, May to November 2013—Table of cases, 2013. Available online: https://www.ecdc.europa.eu/en/publications-data/table-transmission-west-nile-fever-may-november-2013-table-cases-2013 (accessed on 23 June 2020).
- Haussig, J.M.; Young, J.J.; Gossner, C.M.; Mezei, E.; Bella, A.; Sirbu, A.; Pervanidou, D.; Drakulovic, M.B.; Sudre, B. Early start of the West Nile fever transmission season 2018 in Europe. Euro. Surveill. 2018, 23, 1800428. [Google Scholar] [CrossRef] [Green Version]
- Veo, C.; Della Ventura, C.; Moreno, A. Evolutionary Dynamics of the Lineage 2 West Nile Virus That Caused the Largest European Epidemic: Italy 2011-2018. Viruses 2019, 11, 814. [Google Scholar] [CrossRef] [Green Version]
- Semenza, J.C.; Suk, J.E. Vector-borne diseases and climate change: A European perspective. Fems Microbiol. Lett. 2018, 365, fnx244. [Google Scholar] [CrossRef] [PubMed]
- Hubalek, Z.; Tomesek, M.; Kosina, M.; Sikutova, S.; Strakova, P.; Rudolf, I. West Nile virus outbreak in captive and wild raptors, Czech Republic, 2018. Zoonoses Public Health 2019, 66, 978–981. [Google Scholar] [CrossRef] [PubMed]
- Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; et al. Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018. Viruses 2019, 11, 674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UVHVVR. Virus Zahodnega Nila in Virus Usutu. Available online: http://www.uvhvvr.gov.si/si/delovna_podrocja/zdravje_zivali/bolezni/virus_zahodnega_nila_in_virus_usutu/ (accessed on 31 October 2018).




{kind=link}
{kind=link}
{kind=link}
| Mosquito Species | 2017 | 2018 | 2019 |
|---|---|---|---|
| Culex sp. | 54.75% | 96.04% | 25.01% |
| Aedes/Ochlerotatus sp. | 21.79% | 1.34% | 68.98% |
| Anopheles sp. | 20.11% | 1.98% | 3.05% |
| Coquillettidia sp. | 2.79% | 0.35% | 1.29% |
| Culiseta sp. | 0.56% | 0.29% | 1.67% |
| Year | Number of Patients Examined * | Period of the Year | ELISA IgG—WNV Reactive | ELISA IgM—WNV Reactive | ELISA—IgG TBE Reactive and ELISA—IgG WNV Reactive |
|---|---|---|---|---|---|
| 2011 | 44 | August–September | 4 | 0 | 4 out of 4 |
| 2012 | 35 | August–September | 0 | 0 | 0 |
| 2013 | 27 | August–September | 3 | 1 | 2 out of 3 |
| 2014 | 145 | January–December | 10 | 0 | 6 out of 10 |
| 2015 | 102 | January–December | 2 | 0 | 2 out of 2 |
| 2016 | 98 | June–September | 6 | 0 | 6 out of 6 |
| 2017 | 84 | June–September | 0 | 0 | 0 |
| 2018 | 126 | June–September | 4 | 2 | 2 out of 4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knap, N.; Korva, M.; Ivović, V.; Kalan, K.; Jelovšek, M.; Sagadin, M.; Zakotnik, S.; Strašek Smrdel, K.; Slunečko, J.; Avšič-Županc, T. West Nile Virus in Slovenia. Viruses 2020, 12, 720. https://doi.org/10.3390/v12070720
Knap N, Korva M, Ivović V, Kalan K, Jelovšek M, Sagadin M, Zakotnik S, Strašek Smrdel K, Slunečko J, Avšič-Županc T. West Nile Virus in Slovenia. Viruses. 2020; 12(7):720. https://doi.org/10.3390/v12070720
Chicago/Turabian StyleKnap, Nataša, Miša Korva, Vladimir Ivović, Katja Kalan, Mateja Jelovšek, Martin Sagadin, Samo Zakotnik, Katja Strašek Smrdel, Jan Slunečko, and Tatjana Avšič-Županc. 2020. "West Nile Virus in Slovenia" Viruses 12, no. 7: 720. https://doi.org/10.3390/v12070720
APA StyleKnap, N., Korva, M., Ivović, V., Kalan, K., Jelovšek, M., Sagadin, M., Zakotnik, S., Strašek Smrdel, K., Slunečko, J., & Avšič-Županc, T. (2020). West Nile Virus in Slovenia. Viruses, 12(7), 720. https://doi.org/10.3390/v12070720