Identification of the Immediate-Early Genes of Cyprinid Herpesvirus 2


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Virus Preparation
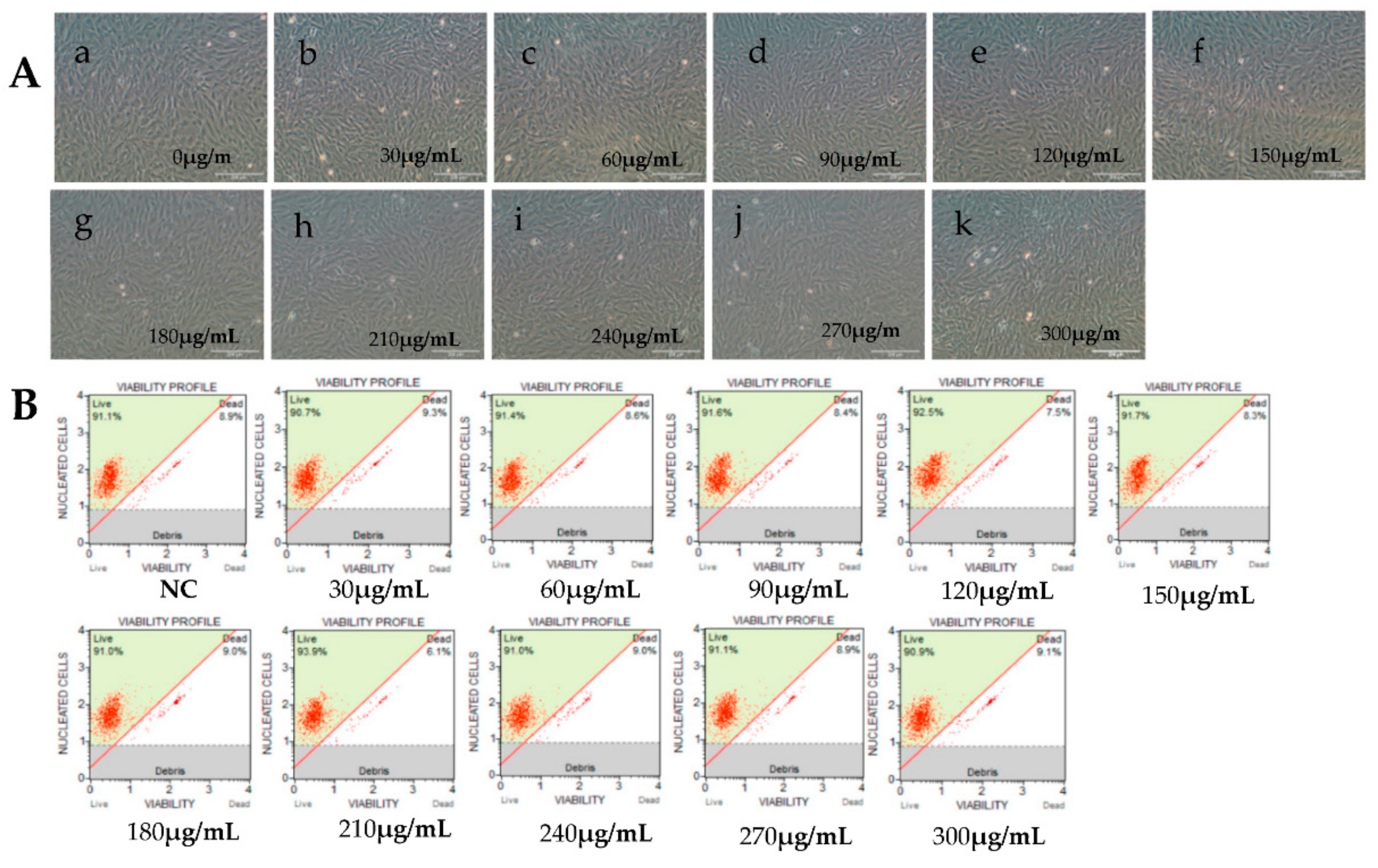
2.2. Cell Viability Test
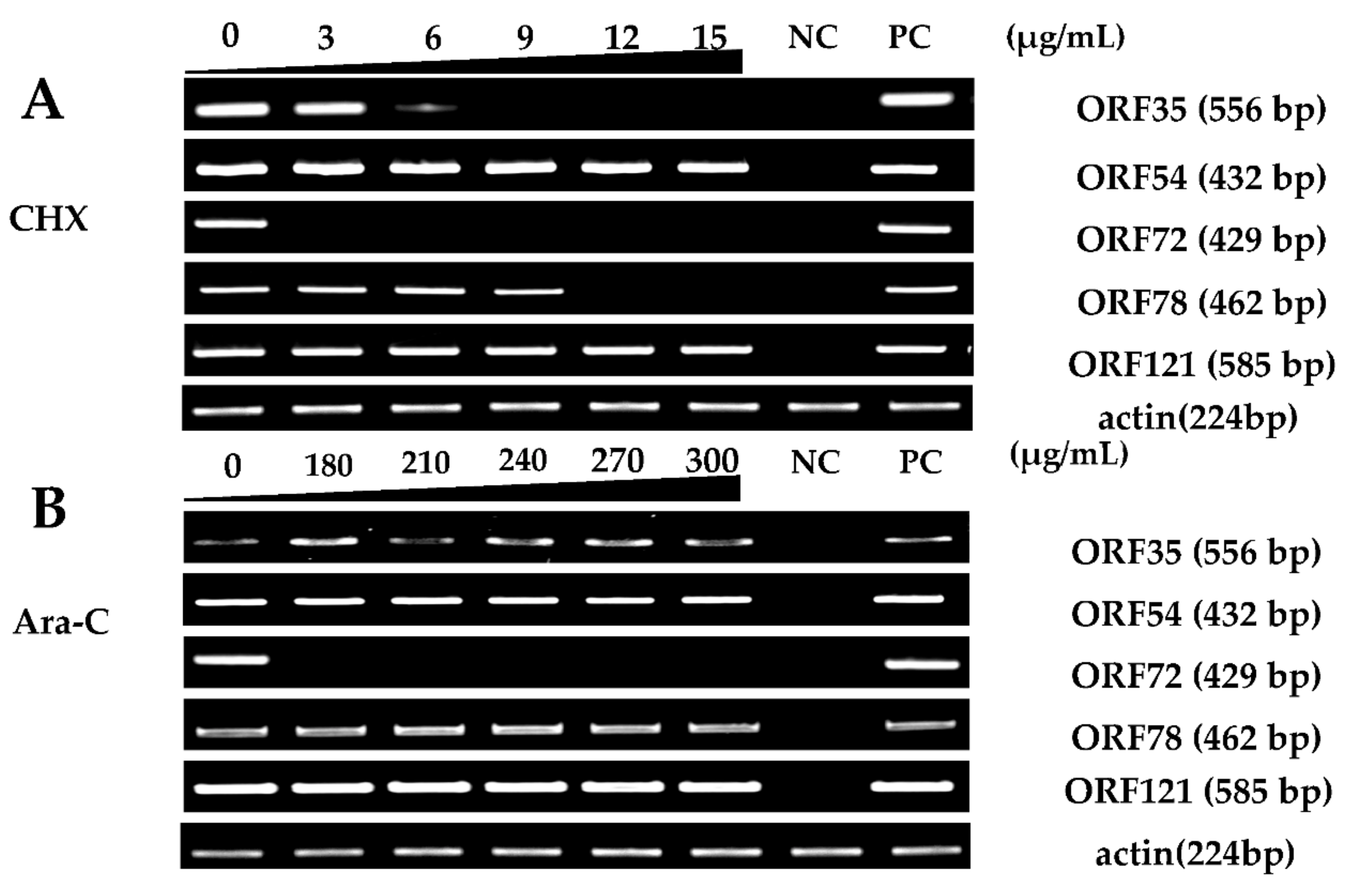
2.3. Protein and DNA Synthesis Inhibitor Treatment of Cultured Cells
2.4. Nucleic Acid Extraction and Quantitative Real-Time Reverse Transcription PCR (qRT-PCR) Analysis
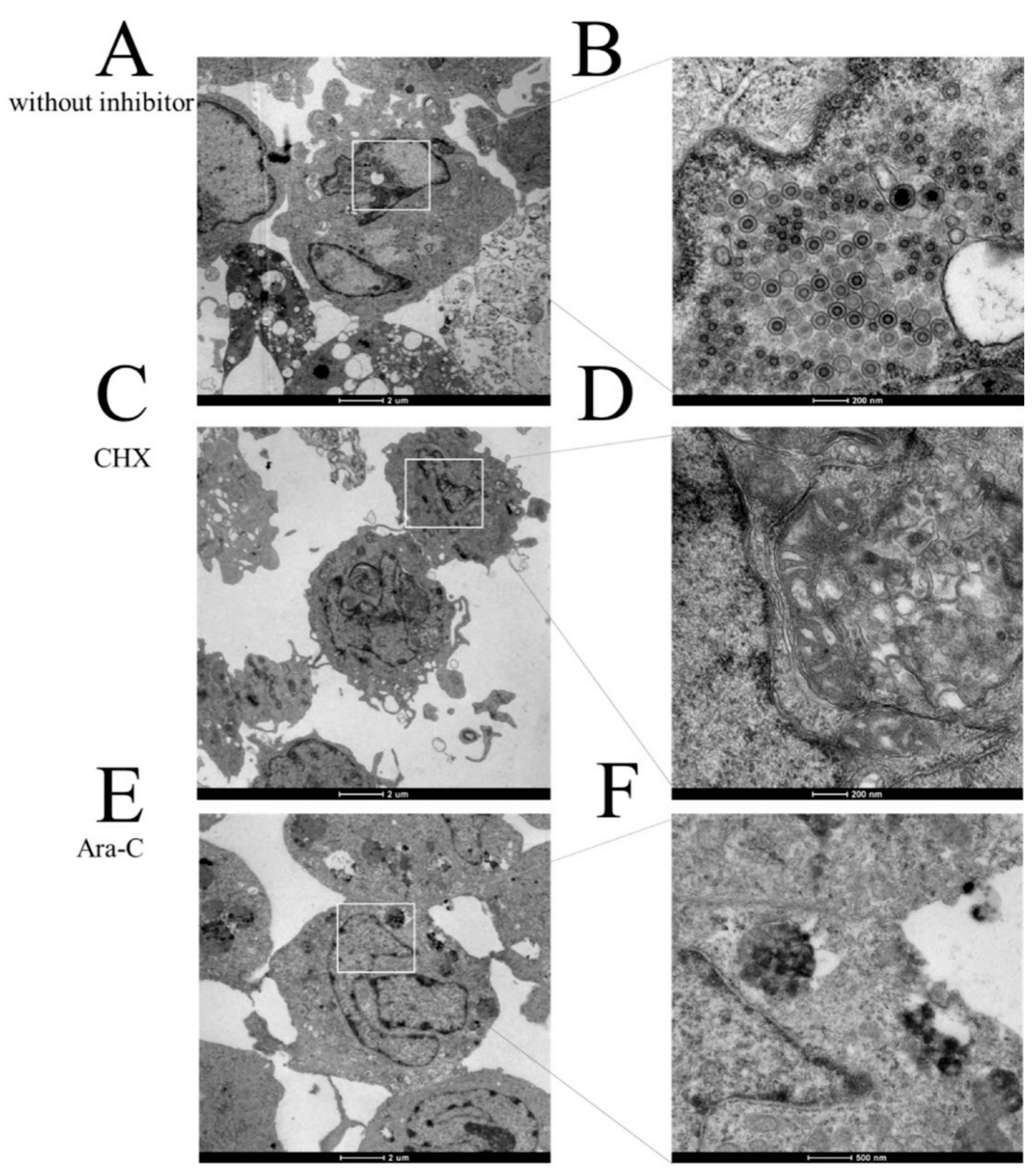
2.5. Electron Microscopy
2.6. Sequencing of the CyHV-2 Transcriptome and Bioinformatic Analyses
2.7. Analysis of CyHV-2 Replication Using PCR and qPCR
3. Results
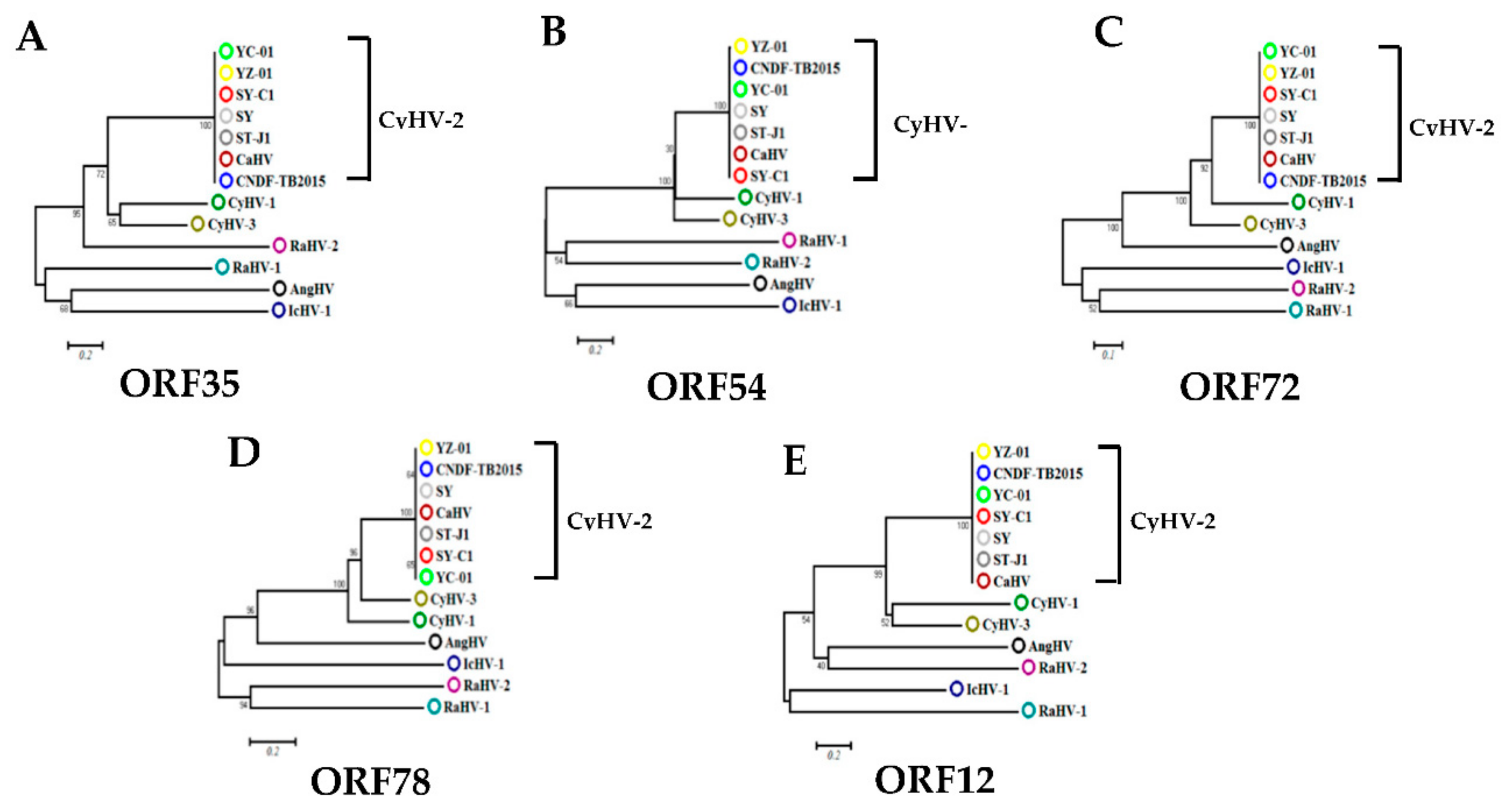
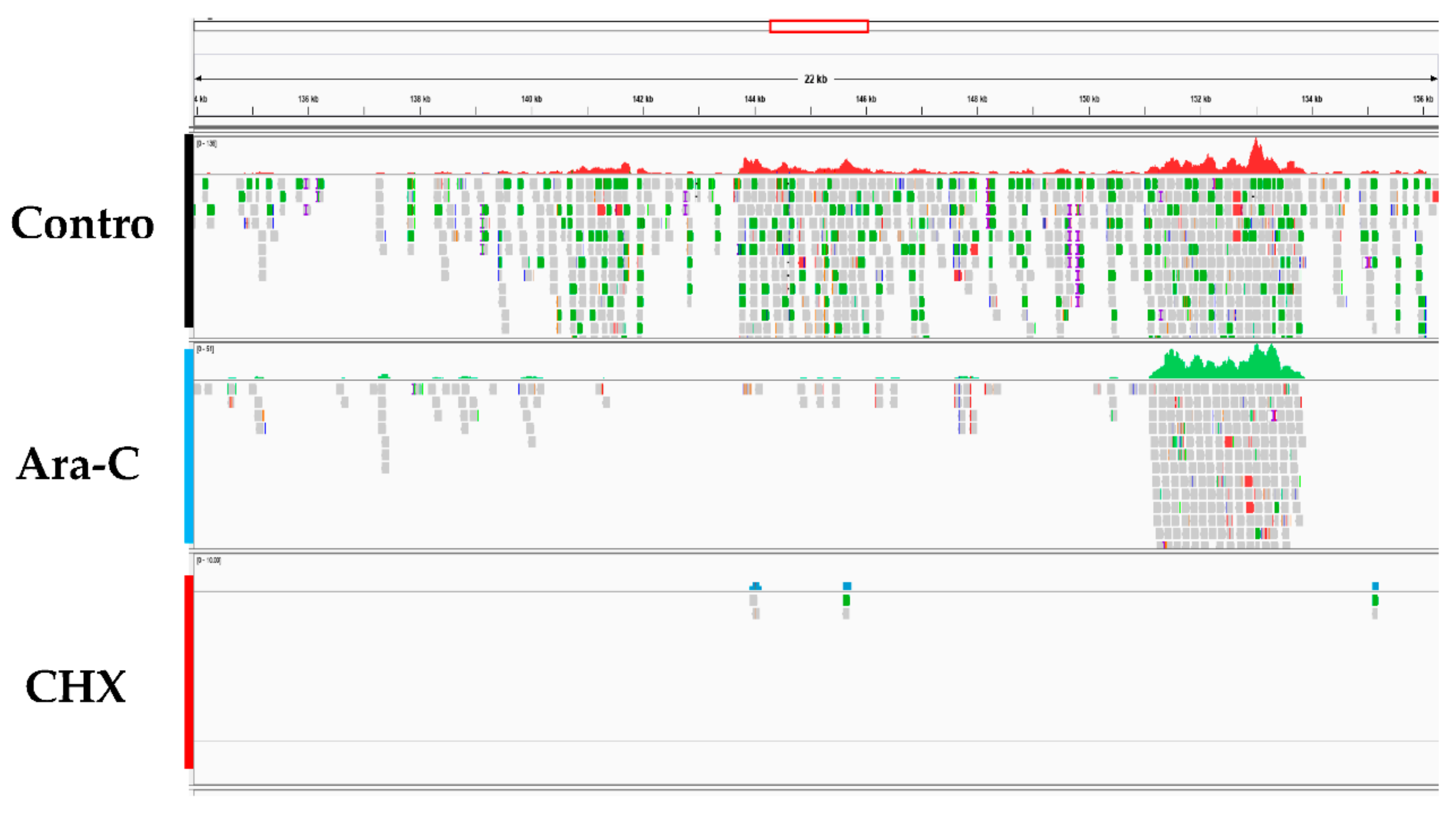
3.1. Screening of CyHV-2 IE Genes
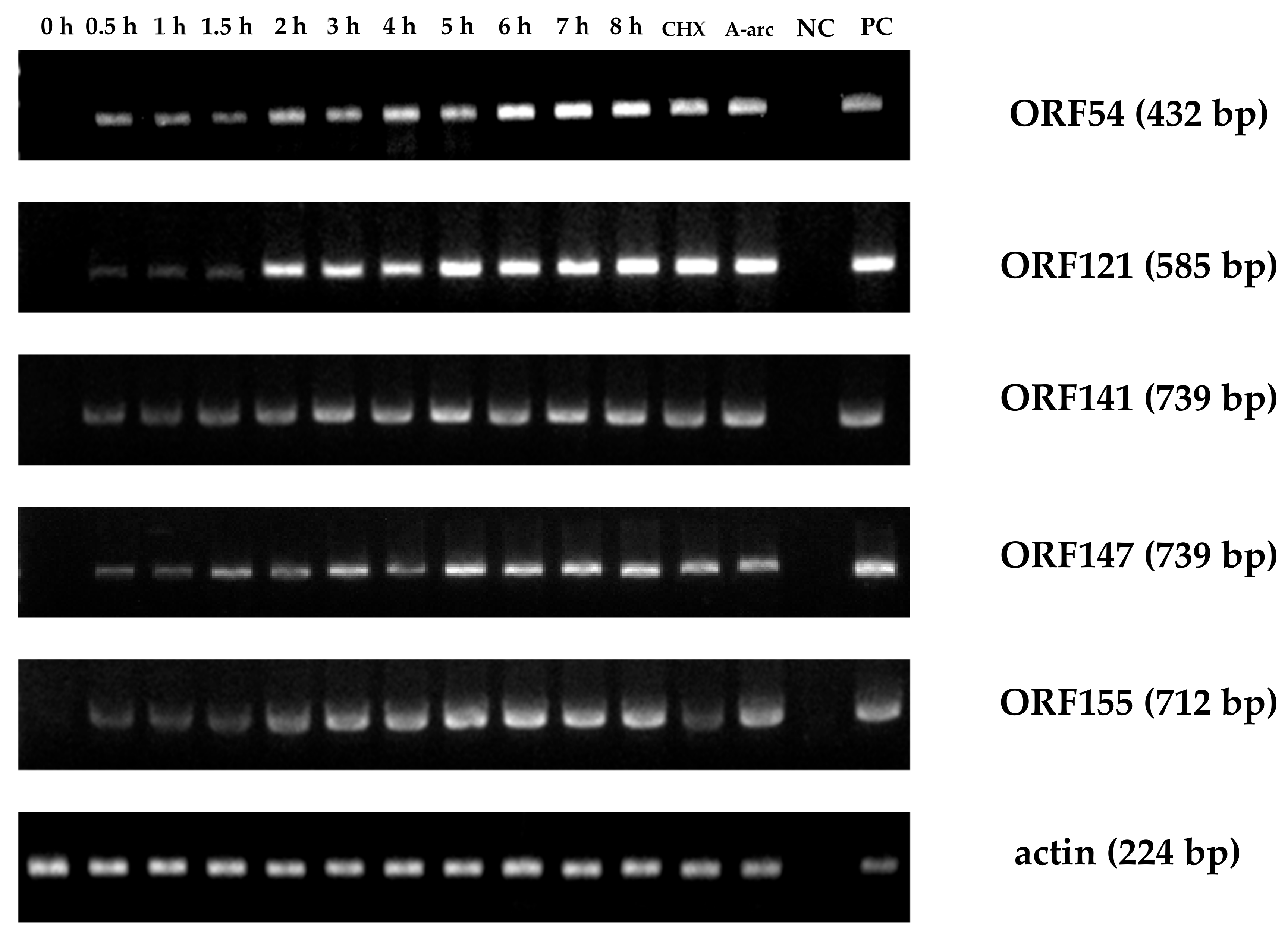
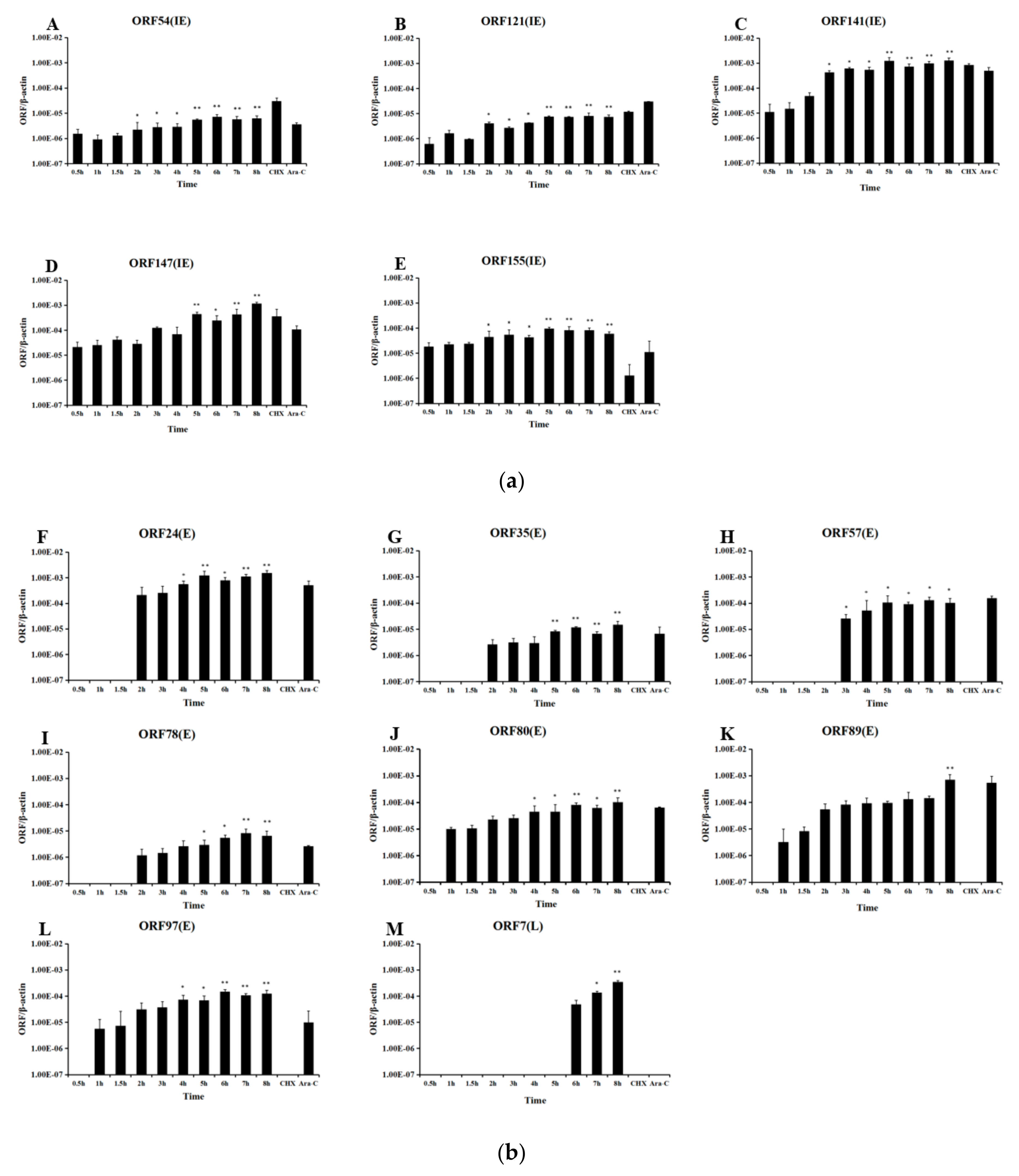
3.2. CyHV-2 IE Genes Confirmation Using qRT-PCR
3.3. IE Gene Temporal Expression during CyHV-2 Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jung, S.J.; Miyazaki, T. Herpesviral Hematopoietic Necrosis of Goldfish, Carassius-Auratus (L). J. Fish Dis. 1995, 18, 211–220. [Google Scholar] [CrossRef]
- Goodwin, A.E.; Merry, G.; Sadler, J. Detection of the herpesviral hematopoietic necrosis disease agent (Cyprinid herpesvirus 2) in moribund and healthy goldfish: Validation of a quantitative PCR diagnostic method. Dis. Aquat. Org. 2006, 69, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, K.R.; Bateman, K.S.; Bayley, A.; Feist, S.W.; Hulland, J.; Longshaw, C.; Stone, D.; Woolford, G.; Way, K. Isolation of a cyprinid herpesvirus 2 from goldfish, Carassius auratus (L.), in the UK. J. Fish Dis. 2007, 30, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Ding, Z.; Ren, M.; An, L.; Xiao, Z.; Liu, P.; Gu, W.; Meng, Q.; Wang, W. The histo- and ultra-pathological studies on a fatal disease of Prussian carp (Carassius gibelio) in mainland China associated with cyprinid herpesvirus 2 (CyHV-2). Aquaculture 2013, 412, 8–13. [Google Scholar] [CrossRef]
- Stephens, F.; Raidal, S.R.; Jones, B. Haematopoietic necrosis in a goldfish (Carassius auratus) associated with an agent morphologically similar to herpesvirus. Aust. Vet. J. 2004, 82, 167–169. [Google Scholar] [CrossRef]
- Reed, A.N.; Izume, S.; Dolan, B.P.; LaPatra, S.; Kent, M.; Dong, J.; Jin, L. Identification of B Cells as a Major Site for Cyprinid Herpesvirus 3 Latency. J. Virol. 2014, 88, 9297–9309. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y. Identification and characterization of herpesviral immediate-early genes. Adv. Struct. Saf. Stud. 2005, 292, 231–244. [Google Scholar] [CrossRef]
- Ilouze, M.; Dishon, A.; Kotler, M. Coordinated and sequential transcription of the cyprinid herpesvirus-3 annotated genes. Virus Res. 2012, 169, 98–106. [Google Scholar] [CrossRef]
- Zeng, X.-T.; Chen, Z.-Y.; Deng, Y.-S.; Gui, J.; Zhang, Q.-Y. Complete genome sequence and architecture of crucian carp Carassius auratus herpesvirus (CaHV). Arch. Virol. 2016, 161, 3577–3581. [Google Scholar] [CrossRef]
- Ito, T.; Kurita, J.; Ozaki, A.; Sano, M.; Fukuda, H.; Ototake, M. Growth of cyprinid herpesvirus 2 (CyHV-2) in cell culture and experimental infection of goldfish Carassius auratus. Dis. Aquat. Org. 2013, 105, 193–202. [Google Scholar] [CrossRef]
- Davison, A.J.; Kurobe, T.; Gatherer, D.; Cunningham, C.; Korf, I.; Fukuda, H.; Hedrick, R.P.; Waltzek, T.B. Comparative Genomics of Carp Herpesviruses. J. Virol. 2012, 87, 2908–2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Q.; Seng, E.K.; Ding, Q.Q.; Zhang, L.L. Characterization of infectious particles of grass carp reovirus by treatment with proteases. Arch. Virol. 2008, 153, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Li, D.F.; Zhang, M.C.; Yang, H.J.; Zhu, Y.B.; Xu, X. Beta-integrin mediates WSSV infection. Virology 2007, 368, 122–132. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Hennig, T.; Whisnant, A.W.; Erhard, F.; Prusty, B.K.; Friedel, C.C.; Forouzmand, E.; Hu, W.; Erber, L.; Chen, Y.; et al. Herpes simplex virus blocks host transcription termination via the bimodal activities of ICP27. Nat. Commun. 2020, 11, 293–313. [Google Scholar] [CrossRef] [Green Version]
- Hagglund, R.; Roizman, B. Role of ICP0 in the Strategy of Conquest of the Host Cell by Herpes Simplex Virus 1. J. Virol. 2004, 78, 2169–2178. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Li, M.; Ke, W.; Ji, Y.; Bian, X.; Yan, X. Identification of the immediate-early genes of white spot syndrome virus. Virology 2009, 385, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.; Huang, H.; Xu, L.; Li, F.; Yang, F. Identification of three immediate-early genes of white spot syndrome virus. Arch. Virol. 2011, 156, 1611–1614. [Google Scholar] [CrossRef]
- Liu, W.-J.; Chang, Y.-S.; Wang, C.-H.; Kou, G.-H.; Lo, C.-F. Microarray and RT-PCR screening for white spot syndrome virus immediate-early genes in cycloheximide-treated shrimp. Virology 2005, 334, 327–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, T.; Nanjo, A.; Saito, M.; Yoshii, K.; Ito, T.; Nakanishi, T.; Sakamoto, T.; Sano, M. In vitro characteristics of cyprinid herpesvirus 2: Effect of kidney extract supplementation on growth. Dis. Aquat. Org. 2015, 115, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xu, L.; Lu, L. Detection of cyprinid herpesvirus 2 in peripheral blood cells of silver crucian carp, Carassius auratus gibelio(Bloch), suggests its potential in viral diagnosis. J. Fish Dis. 2015, 39, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Wen, H.; Wang, H.; Lu, J.; Lu, L.; Jiang, Y. Identification of structure proteins of cyprinid herpesvirus 2. Aquaculture 2020, 523, 735184. [Google Scholar] [CrossRef]
- Davidovich, M.; Dishon, A.; Ilouze, M.; Kotler, M. Susceptibility of cyprinid cultured cells to cyprinid herpesvirus 3. Arch. Virol. 2007, 152, 1541–1546. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Podok, P.; Xie, J.; Lu, L. Comparative analysis of differential gene expression in kidney tissues of moribund and surviving crucian carp (Carassius auratus gibelio) in response to cyprinid herpesvirus 2 infection. Arch. Virol. 2014, 159, 1961–1974. [Google Scholar] [CrossRef] [PubMed]
- Neave, M.J.; Sunarto, A.; McColl, K.A. Transcriptomic analysis of common carp anterior kidney during Cyprinid herpesvirus 3 infection: Immunoglobulin repertoire and homologue functional divergence. Sci. Rep. 2017, 7, 41531. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCR | Gene Name | Nucleotide Sequence (5′–3′) | Amplicon Sizen (bp) | Tm (°C) | |
|---|---|---|---|---|---|
| 1 | ORF1 | F:CTACAGAAGCCTGCCACC | 1341 | 55 | |
| R:TCCGTGGAGTCTGGTCTG | |||||
| 2 | ORF2-A | F:ACCACCACCTCTGCGAAAC | 311 | 55 | |
| R:ATTGTGCCCTGTGCGTTT | |||||
| 3 | ORF3 | F:CGTGTATGGAATCCCTCG | 1035 | 55 | |
| R:TGCCTCTGTGGCTTGTAT | |||||
| 4 | ORF5 | F:CGCTGAACCTGAAACCCTCC | 404 | 60 | |
| R:GCGTTCCCTGTGGTTCCTG | |||||
| 5 | ORF6 | F:TGGCTTTCTGCTGGAGGTG | 824 | 60 | |
| R:CACCAGCACCAACCACTCA | |||||
| 6 | ORF7 | F:CGGTTCTCAGCCAGTTCG | 355 | 60 | |
| R:TGCTTGACCCTCCATCCC | |||||
| 7 | ORF8 | F:CAGGGACCAGACCGAAGACA | 509 | 60 | |
| R:CGTCCTCGTCTCCAGGGTCT | |||||
| 8 | ORF10 | F:AGCAGCAGGTGGCTTCGGATAG | 1643 | 60 | |
| R:GCGGCGTGGATTGTTGGAGT | |||||
| 9 | ORF16 | F:CAAGGGTTCAAGAAAGTAA | 537 | 50 | |
| R:TTGGAGCGTCTATGGTAT | |||||
| 10 | ORF17 | F:CTCGGCGTAGATGTAAGTG | 1314 | 50 | |
| R:GCCTCTGTAACGGATGAA | |||||
| 11 | ORF19 | F:CGAAGGAAACCTGGGAGC | 471 | 60 | |
| R:CGGGCTCGCTTGTTGACT | |||||
| 12 | ORF20 | F:GATGCTGTTGCCGACCTT | 543 | 55 | |
| R:TGCGACGTTACCGACCTA | |||||
| 13 | ORF24 | F:CGAGGGACTCCACAACCG | 470 | 58 | |
| R:TCAAATCACCCAGACAATACCA | |||||
| 14 | ORF25-C | F:CACCCTACCCTCCGAAAT | 655 | 55 | |
| R:TGAACCGCAATACACCTT | |||||
| 15 | ORF25-D | F:AACCAGACCATCACTCGCATAG | 376 | 60 | |
| R:CCACCACTTGGGCTCCTC | |||||
| 16 | ORF33 | F:AGAGTCCGTGAGTAACCTGTGG | 507 | 60 | |
| R:GCATCCGACACCACCTTG | |||||
| 17 | ORF35 | F:GTGATGCCGACAGAGGACA | 554 | 60 | |
| R:TGATACTCGGCTCCCTTCT | |||||
| 18 | ORF41 | F:TTCTTCTGACGCCAACCA | 657 | 58 | |
| R:CACGGCTTGGGTAGGTTG | |||||
| 19 | ORF43 | F:CCAAGCCGTGCGACATAG | 485 | 58 | |
| R:GTTGCCGCCAAACATTAC | |||||
| 20 | ORF44 | F:GCGGAGATAATGGAGGTGA | 450 | 55 | |
| R:CAGGAGTCGGGTTGTTGC | |||||
| 21 | ORF45 | F:CCACGGTCAAGCAGGGTA | 385 | 55 | |
| R:GCGAGGTCGTCGAAATCA | |||||
| 22 | ORF46 | F:ATGGGCGTCGTGGATTAC | 371 | 55 | |
| R:TTCTTGCTGCTGCTGTCG | |||||
| 23 | ORF47 | F:TGAATACGCATCCAACTG | 437 | 50 | |
| R:CGTCCATAGAATCCAACC | |||||
| 24 | ORF49 | F:GTGGCGGTAGATGCTTTC | 305 | 55 | |
| R:CAGGGTCTGATGGTTGGA | |||||
| 25 | ORF50 | F:AAGTCGGCAAGGTGTTTC | 341 | 55 | |
| R:CTTTTGGCGTCTGATGTG | |||||
| 26 | ORF51 | F:ACTCGCAGGTGTACTTCAAGGG | 533 | 60 | |
| R:GGCGACGCTGATGAACACG | |||||
| 27 | ORF52 | F:CTACTACGCTTCTACTGCTCT | 183 | 50 | |
| R:GTGTCGTTGGGTCCTCTT | |||||
| 28 | ORF54 | F:AGGGATGGCCGTCTAGTTTT | 432 | 55 | |
| R:TCGTTGTCGTTGACGAAGAG | |||||
| 29 | ORF56 | F:AGTCCATTTACGACGCTA | 499 | 50 | |
| R:TGGTGAAACGATTTAGGA | |||||
| 30 | ORF57 | F:CGAGTTTCCGCCCTACAA | 425 | 58 | |
| R:CCCAGTGCTGCCAGTGAT | |||||
| 31 | ORF61 | F:CGGCATCCAACACCCTAC | 353 | 55 | |
| R:GAACGAAAGCAGCAGCAA | |||||
| 32 | ORF62 | F:CTCCCATTTCCAACCACTC | 327 | 55 | |
| R:CCTCGTAGACCGACTCCTG | |||||
| 33 | ORF66 | F:CTCAAGGATTTCAGGACG | 487 | 50 | |
| R:TGATGTAAGGGTTGGTGAC | |||||
| 34 | ORF67 | F:CACCCAGACTATTTTACGC | 371 | 50 | |
| R:GCCACTATTGTAGACGATGT | |||||
| 35 | ORF68 | F:GGACATACGAAAGGCTACG | 545 | 58 | |
| R:TTGCGGAGGCTGTGACTT | |||||
| 36 | ORF69 | F:GCCACCTACCTAAACGAGC | 319 | 55 | |
| R:CGTGTTGAGGAAAGCGAGT | |||||
| 37 | ORF72 | F:ACGACCAGAAGGACACCAAG | 429 | 55 | |
| R:GGGGTGGGTAGAGAGGCTAC | |||||
| 38 | ORF74 | F:CCGTGATGAACTTGCTGTT | 189 | 55 | |
| R:GGTACGATTCGGAAGGTAGA | |||||
| 39 | ORF75 | F:CGTCTACTATCAGCCGTGTC | 313 | 55 | |
| R:GCAATAGGTGGTGAGTTCC | |||||
| 40 | ORF76 | F:CAGTTTTCGGTTGGTGTTGAGTTGG | 554 | 55 | |
| R:TTCTGAACGATGATATGAGCGTGAC | |||||
| 41 | ORF77 | F:ACGCTGAACATTTCCACTC | 417 | 50 | |
| R:CGTCCTCGTCCTCTTTGA | |||||
| 42 | ORF78 | F:AGGATTCCCAAACAAAGCC | 462 | 60 | |
| R:GTCATCGTCGGTGAGTTCC | |||||
| 43 | ORF84 | F:GCTCAGGGTCACCAATCA | 388 | 58 | |
| R:ATCCCCGAGTCCGTTCTT | |||||
| 44 | ORF85 | F:CACCAGCGGTAGAAGTCG | 1004 | 55 | |
| R:CCACCCATTGTAATAAACG | |||||
| 45 | ORF86 | F:ATCATCCCTTTCCCAGACA | 534 | 55 | |
| R:GCAAACCGTCCTCAAAGA | |||||
| 46 | ORF88 | F:CTGCTGCCGCTGTTTACC | 121 | 58 | |
| R:TCGCATCCTCGCACTGAC | |||||
| 47 | ORF89 | F:ATGGCGGTGGCATTCCTA | 408 | 60 | |
| R:ATTCGGCGACTGACTTGG | |||||
| 48 | ORF92 | F:GAGCCACCTTCCTGTTCC | 963 | 55 | |
| R:CGTAGAAGAGGGACGAGTG | |||||
| 49 | ORF95 | F:CACTGGGCAGAGGTTGAG | 574 | 55 | |
| R:CAAGTGGTGATCCTGGTCT | |||||
| 50 | ORF97 | F:GACGAGCGATGAATACGG | 574 | 55 | |
| R:GTCTCCGTTGATGGGTCT | |||||
| 51 | ORF101 | F:CGATGACAAGCCTACCGC | 200 | 58 | |
| R:GAGCCAGCCAGTCGTTGA | |||||
| 52 | ORF103 | F:CCGAACCCGAGTTTTATG | 567 | 55 | |
| R:ACGCCACAAACACGACCT | |||||
| 53 | ORF104 | F:GTTACTTTCAACCGAGACGC | 541 | 55 | |
| R:GCCGTCGTTACTGCTGAT | |||||
| 54 | ORF107 | F:AACAGGACTGGGAGTTTG | 230 | 50 | |
| R:GCGACTGAATATGGGATG | |||||
| 55 | ORF109 | F:GGTGCGTCCGACTGGTAGA | 323 | 58 | |
| R:ATGGAGGCGGTGTTCAGC | |||||
| 56 | ORF111 | F:AGATTTGGCGAACGAGGAG | 411 | 60 | |
| R:CGTGTTTGTCGGGCAGGT | |||||
| 57 | ORF112 | F:CAACGAATTGACGGCTAA | 351 | 55 | |
| R:ATGGACCAGAGTGGGAGT | |||||
| 58 | ORF114 | F:CCGTCAACGAGTATTCCA | 585 | 50 | |
| R:CCAGCGTATAGACCCAGA | |||||
| 59 | ORF115 | F:TCGGCGAAGAGTCAAAGG | 471 | 55 | |
| R:CCTGCCACAGCGTAAACA | |||||
| 60 | ORF120 | F:GGTGATTGTAGCCAACGAG | 448 | 55 | |
| R:TTAGAACGACGGTCTGTGAT | |||||
| 61 | ORF121 | F:GCTACTACCACCGTTGTTCCA | 585 | 55 | |
| R:ATCGCTACCATCTTTCTCCTTCT | |||||
| 62 | ORF125 | F:GAGAAAGCAGGTCCGAGTT | 461 | 55 | |
| R:ATCCTCTGGTAGCGTGGG | |||||
| 63 | ORF126 | F:TACGCTACAACGGACAAG | 101 | 50 | |
| R:ATCGCCCAGATCAAAGAC | |||||
| 64 | ORF128 | F:CCACTGCTACCGCTTCTA | 719 | 50 | |
| R:GGCGTACACTCTACCGTCT | |||||
| 65 | ORF129 | F:GTTGGGCTGGCATTGTAT | 314 | 55 | |
| R:CGTTAGAGGTCGGTTCTTTG | |||||
| 66 | ORF130 | F:CACCAAACCCGCCTCATA | 716 | 55 | |
| R:GTATTATTCTGGGTTTGCTTGG | |||||
| 67 | ORF131 | F:GAAGACACGGTCCAAGCG | 444 | 58 | |
| R:CGAGACTCCGATGATTCCTAC | |||||
| 68 | ORF132 | F:ACCATCCGTCGTCAACCA | 407 | 55 | |
| R:AAGCGGTATTGTCGTAGCC | |||||
| 69 | ORF133 | F:TAGGCTCATCAGGCACAT | 319 | 55 | |
| R:GAAGCAGGACAAGACCAAA | |||||
| 70 | ORF135 | F:TCCCAGCCCAACAGACAG | 230 | 55 | |
| R:CCAGAACCCAGGACGAGA | |||||
| 71 | ORF135A | F:CACCCAAGGACCCAGAAC | 489 | 55 | |
| R:CGTCTACGCATCTCAAAGC | |||||
| 72 | ORF139 | F:GAGGTGCGGAGAAGTCAG | 1442 | 55 | |
| R:TCTTGTCCTTATCTCCCAGT | |||||
| 73 | ORF140 | F:TGATCCGTTCAAAGATAGA | 515 | 50 | |
| R:TGATCCGTTCAAAGATAGA | |||||
| 74 | ORF141 | F:ATCCGCTTCAAGTGGCTAA | 739 | 58 | |
| R:GCGTTGAAGATGCGAAGG | |||||
| 75 | ORF143 | F:GAGCAGGAGTAGCAGGGTC | 370 | 55 | |
| R:GCTGATGCGTCTGAGAAT | |||||
| 76 | ORF145 | F:TGGCAGTGTCTGTCTTGTC | 641 | 50 | |
| R:AGATTATACGCCCTTTGG | |||||
| 77 | ORF146 | F:GCGTATGGCGTAGGTAGG | 198 | 55 | |
| R:TGCGAGGATGATGAAGTG | |||||
| 78 | ORF147 | F:ATCCGCTTCAAGTGGCTAA | 739 | 58 | |
| R:GCGTTGAAGATGCGAAGG | |||||
| 79 | ORF147B | F:GAGTGGCTGTGGCTGATG | 461 | 55 | |
| R:GTTGCGGTTGTTTGTCTT | |||||
| 80 | ORF147C | F:GAGTGGCTGTGGCTGATG | 463 | 55 | |
| R:ACGTTGCGGTTGTTTGTC | |||||
| 81 | ORF151A | F:ACAGCGAATGGTTGGAAA | 198 | 55 | |
| R:AACGGTTGAAATGGACAGG | |||||
| 82 | ORF155 | F:AAAACTCTGCTATGCGAAAT | 712 | 55 | |
| R:TGAGGAATGGCTTGACTG | |||||
| 83 | ORF156 | F:ACACGCTACCACCGCCTAC | 713 | 58 | |
| R:CAGCAGCAGCCGTTGAGT | |||||
| 84 | actin | F:CACTGTGCCCATCTACGAG | 224 | 55 | |
| R:CCATCTCCTGCTCGAAGTC | |||||
| RT-PCR | Gene Name | Nucleotide Sequence (5′–3′) | Amplification Efficiency (%) | r2 | Amplicon Size (bp) |
| 1 | actin | F:CACTGTGCCCATCTACGAG | 97.6 | 0.9902 | 224 |
| R:CCATCTCCTGCTCGAAGTC | |||||
| 2 | ORF7-1 | F:TGATGACATCGTCCCCCTCT | 101.5 | 0.9872 | 149 |
| R:TATGGCTGTGCCTCAACGAG | |||||
| 3 | ORF24 | F:TCGACAAGGTCGAAGTGGTG | 98.7 | 0.9972 | 84 |
| R:TGCGAACCACTCCTGTAACC | |||||
| 4 | ORF35 | F:GTGATGCCGACAGAGGACA | 101.8 | 0.9880 | 553 |
| R:TGATACTCGGCTCCCTTCT | |||||
| 5 | ORF54 | F:CAAGATTGCATCGAGCGTGG | 109.6 | 0.9887 | 281 |
| R:TGTCATCGTCGTCCGCATAG | |||||
| 6 | ORF57 | F:GATTCGGGCATCAGAAACGC | 96.6 | 0.9988 | 106 |
| R:GCCCATCACGTAGTAGGCTC | |||||
| 7 | ORF78 | F:AGGATTCCCAAACAAAGCC | 99.0 | 0.9903 | 482 |
| R:GTCATCGTCGGTGAGTTCC | |||||
| 8 | ORF80 | F:TCTGGATCTCGTCGAAAGCG | 91.6 | 0.9932 | 107 |
| R:GCCCAGCCTGAGAAAGAACT | |||||
| 9 | ORF89 | F:CCTGCCCATAAAGGAACGGT | 107.8 | 0.9981 | 88 |
| R:GAGCTCGCGTCCATTATCCA | |||||
| 10 | ORF97 | F:CGAGACGCGTACGATAACCA | 113 | 0.9962 | 147 |
| R:AACGCTTTCAAAACGGGACG | |||||
| 11 | ORF121 | F:GGACATCAAATCGGCAGCTC | 101.4 | 0.9938 | 193 |
| R:CTCCTCCATGGTCACATCGG | |||||
| 12 | ORF141 | F:CAGTGGCTCCCGTACAGTTT | 100.6 | 0.9969 | 114 |
| R:TCGATTGCTTCTTGGGGCTT | |||||
| 13 | ORF147 | F:GTCCTGTCAGTGTGGTAGCC | 97.1 | 0.9991 | 172 |
| R:GTCCATACAGCTGTGGTGCT | |||||
| 14 | ORF147-C | F:TCAACCTGCTCGTGTCACTC | 97 | 0.9823 | 229 |
| R:ACCGTTGCATTACAGTCCGT | |||||
| 15 | ORF155 | F:TCAAGCTGTACTCGTGGCTG | 93.5 | 0.9936 | 73 |
| R:GAAGTGACACACCACAACGC | |||||
| Sample | NCBI BioSample | Treatment | Total Reads | Cleaned Reads | Mapped to CyHV-2 Genome |
|---|---|---|---|---|---|
| 1 | SAMN14501569 | infectin | 98539532 | 98128002 | 26583(0.2697%) |
| A | SAMN14501570 | A-arc | 100866928 | 100455032 | 5887(0.1253%) |
| C | SAMN14501571 | CHX | 46974030 | 46710488 | 2969(0.0029%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, R.; Lu, L.; Wang, B.; Yu, J.; Wang, H. Identification of the Immediate-Early Genes of Cyprinid Herpesvirus 2. Viruses 2020, 12, 994. https://doi.org/10.3390/v12090994
Tang R, Lu L, Wang B, Yu J, Wang H. Identification of the Immediate-Early Genes of Cyprinid Herpesvirus 2. Viruses. 2020; 12(9):994. https://doi.org/10.3390/v12090994
Chicago/Turabian StyleTang, Ruizhe, Liqun Lu, Beiyang Wang, Jiao Yu, and Hao Wang. 2020. "Identification of the Immediate-Early Genes of Cyprinid Herpesvirus 2" Viruses 12, no. 9: 994. https://doi.org/10.3390/v12090994
APA StyleTang, R., Lu, L., Wang, B., Yu, J., & Wang, H. (2020). Identification of the Immediate-Early Genes of Cyprinid Herpesvirus 2. Viruses, 12(9), 994. https://doi.org/10.3390/v12090994