
Astrovirus Infection in Cattle with Nonsuppurative Meningoencephalitis in South Korea

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. RNA Extraction, PCR, Sequencing, and RT-qPCR
2.3. Pathological Examination and In Situ Hybridization (ISH)
2.4. Genetic Evolutionary Analysis
3. Results
3.1. Detection of Bovine Astrovirus
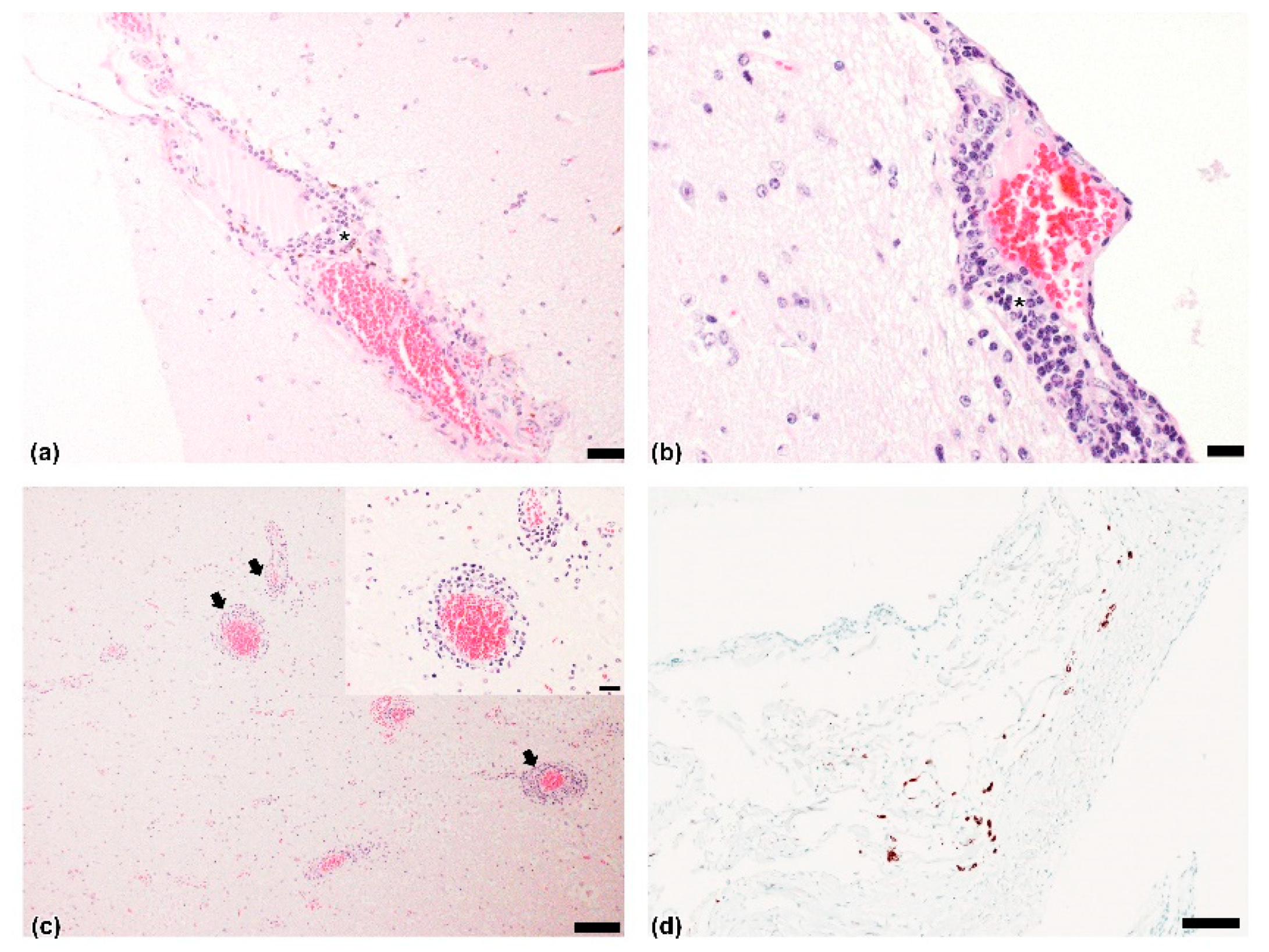
3.2. Pathological and ISH Anlaysis
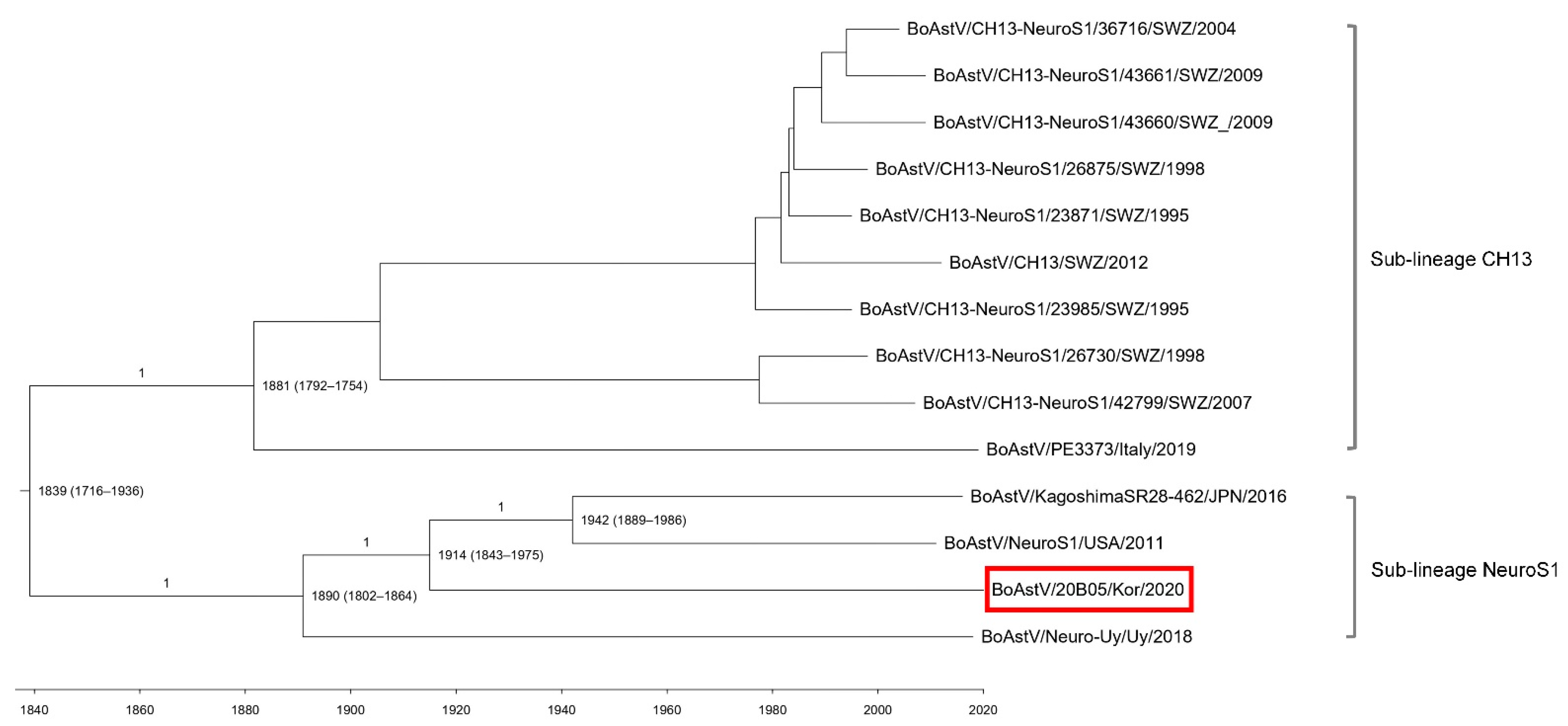
3.3. Phylogenetic and Evolutionary Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cortez, V.; Meliopoulos, V.A.; Karlsson, E.A.; Hargest, V.; Johnson, C.; Schultz-Cherry, S. Astrovirus Biology and Pathogenesis. Annu. Rev. Virol. 2017, 4, 327–348. [Google Scholar] [CrossRef]
- Méndez, E.; Arias, C. Fields Virology, 6th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 623–624. [Google Scholar]
- Wohlgemuth, N.; Honce, R.; Schultz-Cherry, S. Astrovirus evolution and emergence. Infect. Genet. Evol. 2019, 69, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Mendenhall, I.H.; Smith, G.J.; Vijaykrishna, D. Ecological drivers of virus evolution: Astrovirus as a case study. J. Virol. 2015, 89, 6978–6981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humana and animals-molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Pankovics, P.; Boros, Á. Nonsuppurative (Aseptic) meningoencephalomyelitis associated with neurovirulent astrovirus infections in humans and animals. Clin. Microbial. Rev. 2018, 31, e00040-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, P.L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D.; et al. Astrovirus encephalitis in boy with X-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Blomström, A.L.; Frederik, W.; Hammer, A.S.; Belák, S.; Berg, M. Detection of a novel astrovirus in brain tissue of mink suffering from shaking mink syndrome by use of viral metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Diab, S.; McGraw, S.; Barr, B.; Traslavina, R.; Higgins, R.; Talbot, T.; Blanchard, P.; Rimoldi, G.; Fahsbender, E.; et al. Divergent astrovirus associated with neurologic disease in cattle. Emerg. Infect. Dis. 2013, 19, 1385–1395. [Google Scholar] [CrossRef]
- Pfaff, F.; Schlottau, K.; Scholes, S.; Courtenay, A.; Hoffmann, B.; Höper, D.; Beer, M. A novel astrovirus associated with encephalitis and ganglionitis in domestic sheep. Transbound. Emerg. Dis. 2017, 64, 677–682. [Google Scholar] [CrossRef]
- Boros, Á.; Albert, M.; Pankovics, P.; Bíró, H.; Pesavento, P.A.; Phan, T.G.; Delwart, E.; Reuter, G. Outbreaks of neuroinvasive astrovirus associated with encephalomyelitis, weakness, and paralysis among weaned pigs, Hungary. Emerg. Infect. Dis. 2017, 23, 1982–1993. [Google Scholar] [CrossRef]
- Bouzalas, I.G.; Wüthrich, D.; Walland, J.; Drögemüller, C.; Zurbriggen, A.; Vandevelde, M.; Oevermann, A.; Bruggmann, R.; Seuberlich, T. Neurotropic astrovirus in cattle with nonsuppurative encephalitis in Europe. J. Clin. Microbiol. 2014, 52, 3318–3324. [Google Scholar] [CrossRef] [Green Version]
- Schlottau, K.; Schulze, C.; Bilk, S.; Hanke, D.; Höper, D.; Beer, M.; Hoffmann, B. Detection of a novel bovine astrovirus in a cow with encephalitis. Transbound. Emerg. Dis. 2016, 63, 253–259. [Google Scholar] [CrossRef]
- Hirashima, Y.; Okada, D.; Shoichi, S.; Yoshida, S.; Fujisono, S.; Omatsu, T.; Mizutani, T.; Nagai, M. Whole genome analysis of a novel neurotropic bovine astrovirus detected in a Japanese black steer with non-suppurative encephalomyelitis in Japan. Arch. Virol. 2018, 163, 2805–2810. [Google Scholar] [CrossRef] [PubMed]
- Giannitti, F.; Caffarena, R.D.; Pesavento, P.; Uzal, F.A.; Maya, L.; Fraga, M.; Colina, R.; Castells, M. The first case of bovine astrovirus-associated encephalitis in the southern hemisphere (Uruguay), uncovers evidence of viral introduction to the Americas from Europe. Front. Microbiol. 2019, 10, 1240. [Google Scholar] [CrossRef] [Green Version]
- Zaccaria, G.; Lorusso, A.; Hierweger, M.M.; Malatesta, D.; Defourny, S.V.; Ruggeri, F.; Cammà, C.; Ricci, P.; Domenico, M.D.; Rinaldi, A.; et al. Detection of astrovirus in a cow with neurological signs by nanopore technology, Italy. Viruses 2020, 12, 530. [Google Scholar] [CrossRef]
- Selimovic-Hamza, S.; Sanchez, S.; Philibert, H.; Clark, E.G.; Seuberlich, T. Bovine astrovirus infection in feedlot cattle with neurological disease in western Canada. Can. Vet. J. 2017, 58, 601–603. [Google Scholar] [PubMed]
- Spinato, M.T.; Vince, A.; Cai, H.; Ojkic, D. Identification of bovine astrovirus in cases of bovine non-suppurative encephalitis in eastern Canada. Can. Vet. J. 2017, 58, 607–609. [Google Scholar] [PubMed]
- Oem, J.K.; An, D.J. Phylogenetic anlaysis of bovine astrovirus in Korean cattle. Virus Genes 2014, 48, 372–375. [Google Scholar] [CrossRef]
- Lee, S.Y.; Son, K.D.; Kim, Y.S.; Wang, S.J.; Kim, Y.K.; Jheong, W.H.; Oem, J.K. Genetic diversity and phylogenetic analysis of newly discovered bat astroviruses in Korea. Arch. Virol. 2018, 163, 3065–3072. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Lam, T.T.; Carvalho, L.M.; Pybus, O.G. Exploring the temporal structure of heterochronous sequences using TempEst (formerly Path-O-Gen). Virus Evol. 2016, 2, vew007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selimovic-Hamza, S.; Boujon, C.L.; Hilbe, M.; Oevermann, A.; Seuberlich, T. Frequency and pathological phenotype of bovine astrovirus CH13/NeuroS1 infection in neurologically-diseased cattle: Towards assessment of causality. Viruses 2017, 9, 12. [Google Scholar] [CrossRef]
- Theil, D.; Fatzer, R.; Schiller, I.; Caplazi, P.; Zurbriggen, A.; Vandevelde, M. Neuropathological and aetiological studies of sporadic non-suppurative meningoencephalomyelitis of cattle. Vet. Rec. 1998, 143, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Kauer, R.V.; Koch, M.C.; Schönecker, L.; Becker, J.; Holwerda, M.; Glaus, A.N.; Hierweger, M.M.; Werder, S.; Dijkman, R.; Meylan, M.; et al. Fecal shedding of bovine astrovirus CH13/NeuroS1 in veal calves. J. Clin. Microbiol. 2020, 58, e01964-19. [Google Scholar] [CrossRef]
- Arias, C.F.; DuBois, R.M. The astrovirus capsid: A review. Viruses 2017, 9, 15. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Brain Tissue | RT-PCR 1 | Position (Nucleotide) | Cycle Threshold Value | Copy Number |
|---|---|---|---|---|
| Cerebellum | + (+) | 2677–3791 | 22.53 | 5.37E6 |
| Forebrain | + (+) | 486–1386, 1932–5992 | 21.34 | 1.17E7 |
| Midbrain | + (+) | 2708–3797 | 21.70 | 9.33E6 |
| Hindbrain | − (+) | 3400–3788 | 25.04 | 1.07E6 |
| Lymph node | − (+) | 3403–3788 | 30.53 | 3.09E4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-Y.; Kim, J.-H.; Kim, Y.-J.; Kim, Y.-S.; Roh, S.-G.; Lee, K.-H.; Kim, H.-J.; Shin, J.-H.; Oem, J.-K. Astrovirus Infection in Cattle with Nonsuppurative Meningoencephalitis in South Korea. Viruses 2021, 13, 1941. https://doi.org/10.3390/v13101941
Lee S-Y, Kim J-H, Kim Y-J, Kim Y-S, Roh S-G, Lee K-H, Kim H-J, Shin J-H, Oem J-K. Astrovirus Infection in Cattle with Nonsuppurative Meningoencephalitis in South Korea. Viruses. 2021; 13(10):1941. https://doi.org/10.3390/v13101941
Chicago/Turabian StyleLee, Sook-Young, Jong-Ho Kim, Yoon-Ji Kim, Young-Sik Kim, Su-Gwon Roh, Kyung-Hyun Lee, Heui-Jin Kim, Jae-Ho Shin, and Jae-Ku Oem. 2021. "Astrovirus Infection in Cattle with Nonsuppurative Meningoencephalitis in South Korea" Viruses 13, no. 10: 1941. https://doi.org/10.3390/v13101941
APA StyleLee, S.-Y., Kim, J.-H., Kim, Y.-J., Kim, Y.-S., Roh, S.-G., Lee, K.-H., Kim, H.-J., Shin, J.-H., & Oem, J.-K. (2021). Astrovirus Infection in Cattle with Nonsuppurative Meningoencephalitis in South Korea. Viruses, 13(10), 1941. https://doi.org/10.3390/v13101941