1. Introduction
Foot-and-mouth disease (FMD), a clinical acute vesicular and extremely contagious infection, is transmissible among clove hoofed animals [
1]. It is caused by the foot-and-mouth disease virus (FMDV), which belong to the
Aphthovirus genus and
Picornaviridae family. It is an 8.5 kb positive-sense single-stranded RNA virus [
2]. Currently, FMDV O and A serotypes are widespread in China. The FMDV serotype A (A/WH/CHA/09 FMDV) was isolated from the 2009 outbreak in Wuhan, China. Subsequently, from 2009 to 2010, many outbreaks that are closely associated with the above strain have been reported in other regions of China. In 2013, A/GDMM/CHA/2013 FMDV, a new strain, occurred in Maoming, Guangdong, China. FMD is associated with high economic losses [
3]. Inactivated vaccines play a fundamental role in controlling FMD outbreaks, especially in developing countries [
4]. Although chemically inactivated vaccines provide good protection, they have various limitations, such as their potential for escape of a live virus during production and application and their high production costs [
5]. The technology used in multi-epitope proteins as well as their application as vaccines has been widely evaluated [
6,
7].
Antigenic region structures on capsid surfaces of FMDV have been characterized [
8]. An extremely preserved Arg-Gly-Asp (RGD) triplet motif, which is localized on highly mobile exposed G-H loop (140–160 aa) of the capsid protein, VP1, is a key site for viral entry into host cells and is capable of inducing neutralizing antibodies against this virus [
9]. On the VP1 of FMDV, the 41–60 aa as well as 200–213 aa are the major neutralizing epitopes that can elicit a strong protective immune response [
10,
11,
12]. Besides, T cell epitopes of FMDV are critical in enhancement of the immunogenicity of peptide vaccines. Collen et al. linearly coupled the T cell epitope (20–40 aa) on VP1 to the G-H loop of VP1 to immunize cattle is a virus challenge experiment [
8]. Moreover, T cell epitopes of the 3A protein (21–35 aa) can enhance immune responses to FMDV epitope vaccines [
13].
About 90% of pathogenic infections, including FMDV, are transmitted through the mucosal areas. Therefore, mucosal vaccination can help establish a protective immunity against these infections, overcoming the challenges of present injection-based vaccines [
14]. Consequently, inoculation of multi-epitope antigens through the mucosal route of entry may improve protective responses against FMDV. Oral mucosal vaccines are advantageous ince their mode of administration is convenient. They are also cost-effective and require simplified logistics with a lower dependence on cold chains during shipments and distribution [
15]. However, currently, the commercially accessible oral mucosal vaccines are very few in number. This is attributed to challenges in antigenic delivery into the mucosa and tolerance of the oral mucosal immunity [
16]. Thus, to develop efficient oral mucosal vaccines, Microfold cells (M cells) are decent targets for delivering antigens and immune response stimulation [
17]. M cells, which are epithelial cells, are localized in Peyer’s patches (PPs) in the intestines, nasopharynx-related lymphoid tissue (NALT), isolated lymphoid follicles, and the appendix. These cells are responsible for the monitoring and transcytosis of antigens, microorganisms, and pathogens [
18,
19]. M cells play vital roles as gatekeepers of mucosal immunity, even though, in the intestinal tract, only 1 in 10 million epithelial cells is an M cell [
20]. Therefore, it is interesting to target antigens to M cells to enhance immune responses. M cell-targeting ligands Co1, obtained through screening of phage display library against in vitro human M-like cells, can promote M cell uptake of oral vaccines and improve antigen-specific immune reactions in systemic as well as mucosal surfaces [
21,
22].
Currently,
L. lactis is a good strategy as a delivery vehicle for orally administered mucosal vaccines.
L. lactis, a model lactic acid bacterium, is generally recognized as safe (GRAS) owing to its longstanding use in human food fermentations and products [
23]. This bacterium is used in the development as well as delivery of cytokines and antigens to mucosal surfaces [
23]. The nisin-controlled expression (NICE) system has been developed for use in
L. lactis [
24]. This system is the most studied and widely used in expression of exogenous proteins.
We created two recombinant L. lactis-TB1 and L. lactis-TB1-Co1 as multi-epitope mucosal vaccines, which can express the FMDV multi-epitope antigens, TB1 and TB1-Co1, via the NICE system, respectively. We evaluated their immune effects after oral immunization in mice and guinea pigs.
2. Materials and Methods
2.1. Animals, Commercial Vaccine, Virus, Bacterial Strain, and Cells
Animals were procured from the experimental animal center at the Lanzhou Veterinary Research Institute (LVRI). The Institutional Animal Care and Use Committee (IACUC) of LVRI approved this study (No. LVRIAEC2018−008). Type A commercial FMDV inactivated vaccine, positive serum from type A FMDV infected swine, and purified inactivated type A FMDV were acquired from OIE/National FMD Reference Laboratory of China. Preserved strains of FMDV, including A/GDMM/CHA/2013, A/WH/CHA/09 and AF72 were acquired from the OIE/National FMD Reference Laboratory of China. L. lactis NZ9000 (glycerol bacteria) and L. lactis expression plasmid pNZ8148 (dry powder) were bought from MoBiTec, Germany. The L. lactis were cultured on an M17 medium (Oxoid, Basingstoke, UK) with 0.5% glucose (GM17) at 30 °C without shaking. Baby Hamster Kidney (BHK) cells were bought from China Center for Type Culture Collection and seeded in the Dulbecco’s modified Eagle’s medium (DMEM, Gibco, Waltham, MA, USA) with 10% fetal bovine serum (FBS, Gibco, Waltham, MA, USA), 100 IU/mL penicillin as well as 100 μg/mL streptomycin in a 5% CO2 atmosphere at 37 °C.
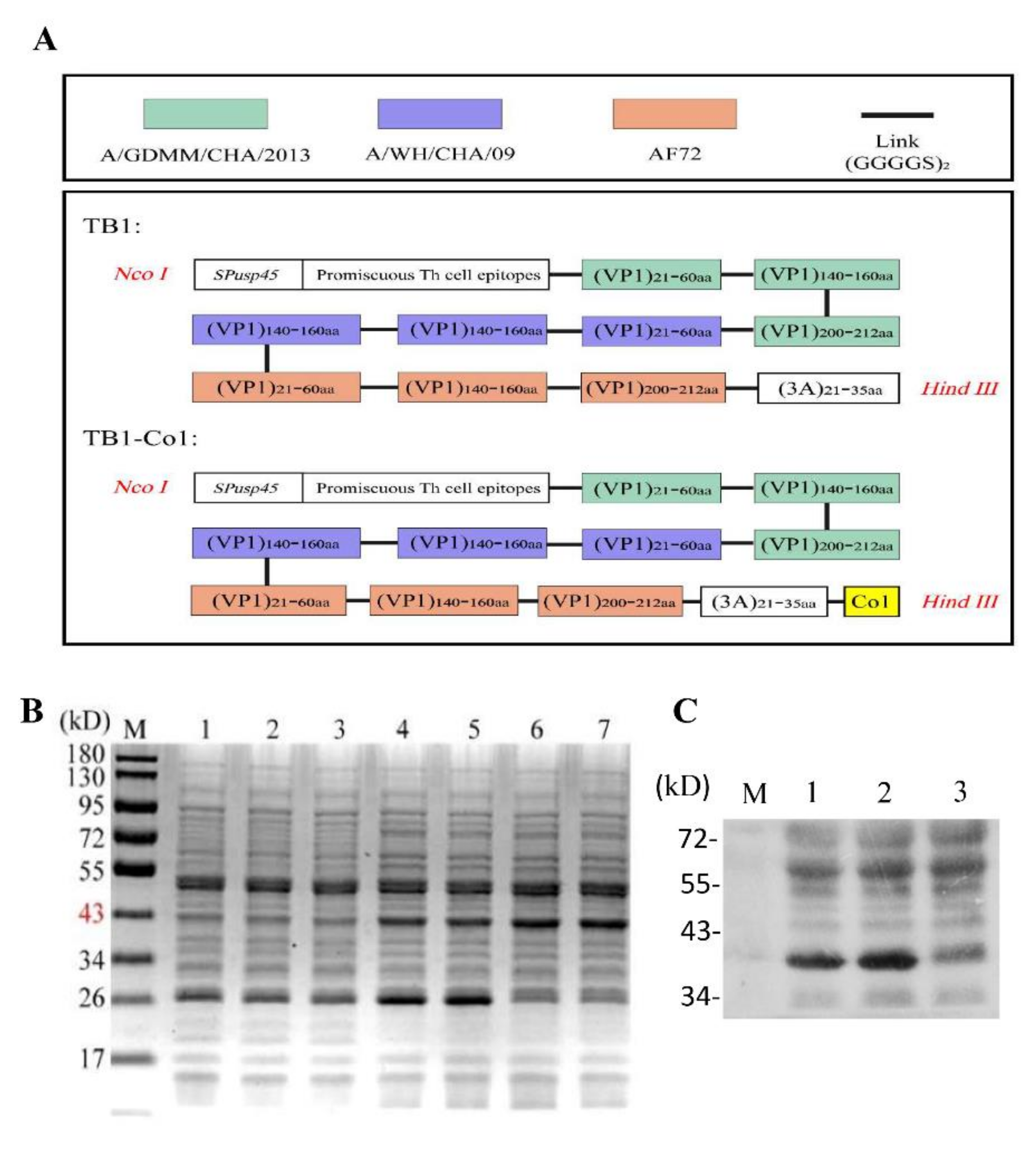
2.2. Multi-Epitope Gene Design and Synthesis
Signal peptide
SPusp45 was introduced into the recombinant protein to promote the secretion of the proteins into the culture medium [
25]. Promiscuous T helper (Th) cell epitopes were employed in development of immunogenic recombinant multi-epitope proteins [
26]. The G-H loop domain as well as VP1 epitopes from the FMDV strains, including A/GDMM/CHA/2013, A/WH/CHA/09 and AF72 (
Table 1) were linked by spacer sequences (GGGGS)
2, as shown in
Table 2. Synthesis of the two DNA sequences that encode the multi-epitope proteins (
Table 2) was performed by Nanjing GenScript Biotech Co., Ltd. (Nanjing, China) These sequences were codon optimized with
L. lactis, followed by their insertion into a pUC57 vector with
NcoI as well as
HindIII restriction sites on 5′ and 3′ ends, respectively, of multi-epitope genes. Finally, recombinant plasmids pUC57-TB1 and pUC57-TB1-Co1 were obtained. Recombinant plasmids were sequenced for verification. In addition, nucleotide sequences of recombinant proteins and oral adjuvant as shown in
Table 3.
2.3. Plasmids Were Introduced into L. lactis NZ9000 through Electrotransformation
The TB1 gene was obtained from the pUC57-TB1 plasmid and inserted in corresponding sites of the expression plasmid, pNZ8148, to yield the recombinant expression plasmid, pNZ8148-TB1. Through the same method, we obtained the recombinant expression plasmid, pNZ8148-TB1-Co1. In our previous study, were described the preparation process of NZ9000 competent cells [
28]. After melting NZ9000 competent cells on ice, 50 μL of recombinant expression plasmids were added followed by mixing with ice-cold competent cells (100 μL), and placed for 5 min on mice. Transformation of plasmids into competent cells was carried out by 0.2 cm cuvettes in a Gene Pulser electroporator (Bio-Rad, USA) at 200 Ω, 25 µF, and 2.5 kV. The duration of a single electrical pulse was 1.5–5 ms. Cuvettes were immediately mixed with 1 mL of GM17 broth (minus antibiotics but with 20 μL MgCl
2 and 2 μL CaCl
2) and placed on ice for 10 min. Incubation was carried out at 30 °C, minus shaking. Then, recombinant
L. lactis strains were grown and selected on GM17 agar medium with chloramphenicol (10 μg/mL). Finally, we obtained two recombinant strains
L. lactis-TB1 and
L. lactis-TB1-Co1.
2.4. Expression Levels of Recombinant Proteins
To obtain the TB1 and TB1-Co1 recombinant proteins, strains were grown in GM17 broth with chloramphenicol (10 μg/mL), induced at OD600 0.4 using nisin (5 ng/mL) at 30 °C followed by collection at OD600 1.0. Resuspension of harvested cells was performed in PBS after which preparation of the recombinant proteins was carried out after ultrasonic decompositions for analysis by 12% sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE). Confirmation of protein specificity was carried out by western blot using the swine anti-FMDV antibodies (1:1000 dilution) and a horseradish-peroxidase (HRP)-conjugated goat anti-swine IgG antibody (1:10,000 dilution; Abbkine Scientific Co., Ltd., Wuhan, China). Visualization used ECL-chemiluminescentkit (ECL-plus, Thermo Scientific, Pittsburgh, PA, USA).
2.5. Mice Immunization
Bacterial cells (NZ9000/pNZ8148, L. lactis-TB1 and L. lactis-TB1-Co1) in GM17 broth with chloramphenicol (10 μg/mL), were induced at OD600 0.4 using nisin (5 ng/mL) at 30 °C. Cells were collected at OD600 1.0, washed using PBS and attuned to 1 × 109 colony forming units per milliliter (CFUs/mL) prior to inoculation.
Specific pathogen-free (SPF) female BALB/c mice (125 in number, aged 7 weeks) were randomized in 5 groups of 25 each (
Table 4). Each animal in the first four groups was orally immunized by gavage. Immunizations were conducted at three time points (days 1, 11, and 21) for three successive days at every time point. Mice in group E were immunized by intramuscular injection on day one, followed by one booster immunization at a week interval.
Blood samples were obtained by randomly selecting three experimental mice from each group at various time points (that is, on days 10, 20, 30, 37, 44 and 51 after immunization). Then, 300 μL of blood for serum testing was obtained from the angular vein. After mice had been sacrificed by an overdose of ether anesthesia, intestinal mucus samples in 200 μL of sterilized PBS as well as lung lavage liquid specimens in 300 μL of sterilized PBS (three mice per group every time) were obtained using cotton swabs into 1.5 mL centrifuge tubes with 0.01 M EDTA-Na2, followed by repeated rinsing.
2.6. Immunization of Guinea Pigs and Challenge
Cells (NZ9000/pNZ8148,
L. lactis-TB1 and
L. lactis-TB1-Co1) grown at 30 °C in GM17 broth with 10 μg/mL of chloramphenicol, were induced at OD
600 0.4 using nisin (5 ng/mL) and harvested at OD
600 1.0. They were washed and resuspended in PBS. Before inoculation, final cell concentrations were adjusted to 1 × 10
10 CFU/mL. Thirty-five FMDV-specific antibody negative female
guinea pigs (230 to 250 g in weight) were randomized into seven groups of five guinea pigs each (
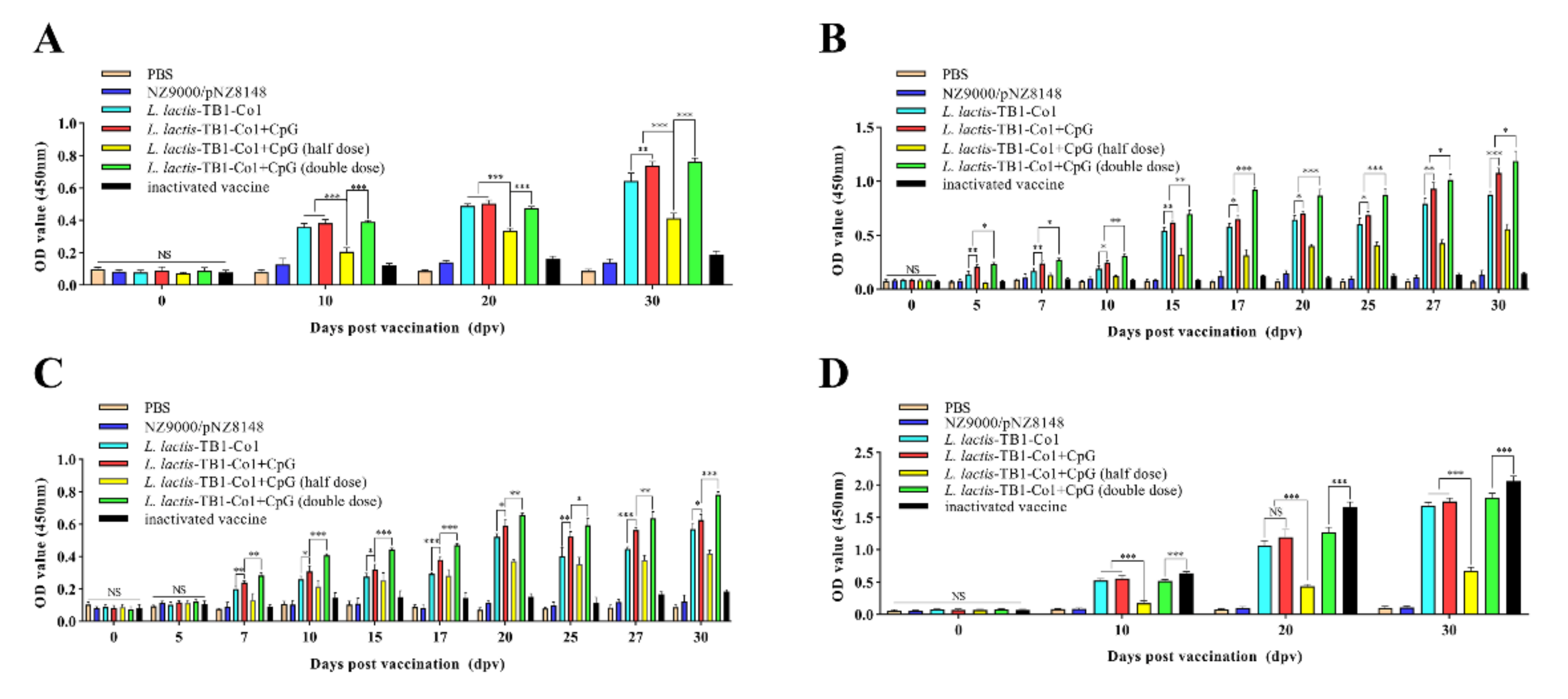
Table 5). Guinea pigs in groups D, E, and F were treated with CpG ODN (5 μg/guinea pig). Immunization procedures were the same as those of mice. Then, for serum tests, 800 μL of blood samples were obtained by cardiac puncture on days 0 (pre-immune day), 10, 20, and finally 30. Saliva and anal swab samples were collected at different time points (0 d, 5 d, 7 d, 10 d, 15 d, 17 d, 20 d, 25 d, 27 d and 30 d) after immunization and kept at −80 °C for successive analyses.
All guinea pigs were challenged with 0.2 mL of 100 GPID50 (50% guinea pig infective doses) of FMDV strain AF72 with a subcutaneous injection in the left rear foot at 30 dpv. Guinea pigs in every group were observed for a total of 10 days and clinical symptoms recorded. Assessment of protection of challenged guinea pigs was performed by the absence of FMD-associated lesions, except at the injection site, and vice versa.
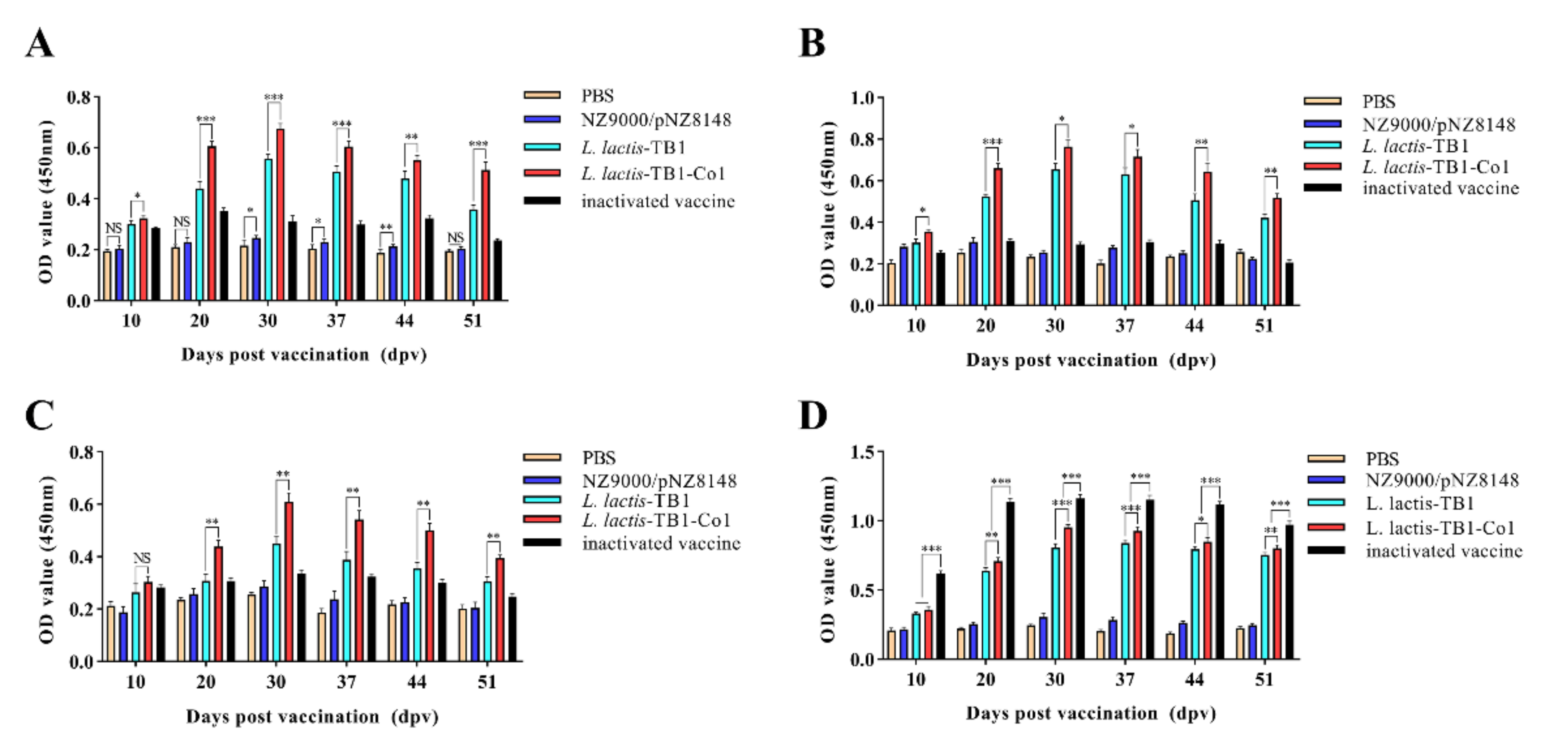
2.7. Detection of IgG and IgA
Levels of antibodies against FMDV in sera were assessed by ELISA. In brief, coating of 96-well microplates for 12 h was carried out using inactivated whole-virus antigen of FMDV at 4 °C and washed 5 times using PBST. Then, well blocking was performed for at 37 °C for 2 h using 100 μL/well 5% skim milk in PBST. Wells were washed five times using PBST. Serum samples (1:50 dilution) from guinea pigs or mice were added (100 μL/well) and incubated at 37 °C for 1 h. Plates were washed five times using PBST. For mouse serum samples, HRP-conjugated rabbit anti-mouse IgG antibody (1:1000 dilution) or HRP-conjugated sheep anti-mouse IgA antibody (1:10,000 dilution) were added (100 μL/well). For serum samples from guinea pigs, HRP-conjugated rabbit anti-guinea pig IgG (1:1000 dilution) or goat anti-guinea pig IgA (1:10,000 dilution) were added (100 μL/well). Incubation of the microplates was performed for 1 h at 37 °C, washed five times using PBST after which the TMB single-component substrate solution was added (50 μL/well). Incubation of the plates for 15 min was then performed at 37 °C. At last, stop solution (50 μL/well; 2 M H2SO4) was used to terminate the reactions. Optical absorbance for antibody reactivity was determined at 450 nm within 15 min via a microplate reader.
2.8. Detection of SIgA
Detection of SIgA antibody concentrations against FMDV in mice and guinea pig samples was carried out by ELISA. Mice samples included intestinal and lung lavage fluid, while guinea pig samples included saliva and anal swabs. ELISA assays were conducted as previously described.
2.9. Viral Neutralizing Antibody Tests (VNT)
Serum specimens from guinea pigs were obtained on day 30 after first immunization and evaluated via viral-neutralizing antibody tests (VNTs) using BHK-21 cells. After 30 min of inactivation of the serum samples at 56 °C, the double ratio dilution method (from 1:2 to 1:256) was used to add 50 μL serum samples to each well of 96-well plates with DMEM (Gibco, Loughborough, UK) with 2% FBS. The serum samples from guinea pigs that had been orally immunized with PBS or L. lactis NZ9000/pNZ8148 were used as controls. In a 96--well plate, serum samples were two-fold serial diluted (50 μL/well), after which an equal volume of FMDV AF72 solution (200 TCID50/100 μL) was added to the plates at 37 °C for 1 h. Then, addition of BHK cells (100 μL; 106 cells/mL) to the antibody-virus mixture was followed by incubation in a 5% CO2 environment at 37 °C for 72 h. Based on the Karber method, neutralizing antibody titers were assessed as reciprocal log10 of highest dilutions that could exert protective effects to 50% of the cells against cytopathic effects (CPE).
2.10. Analysis of Cytokine Levels
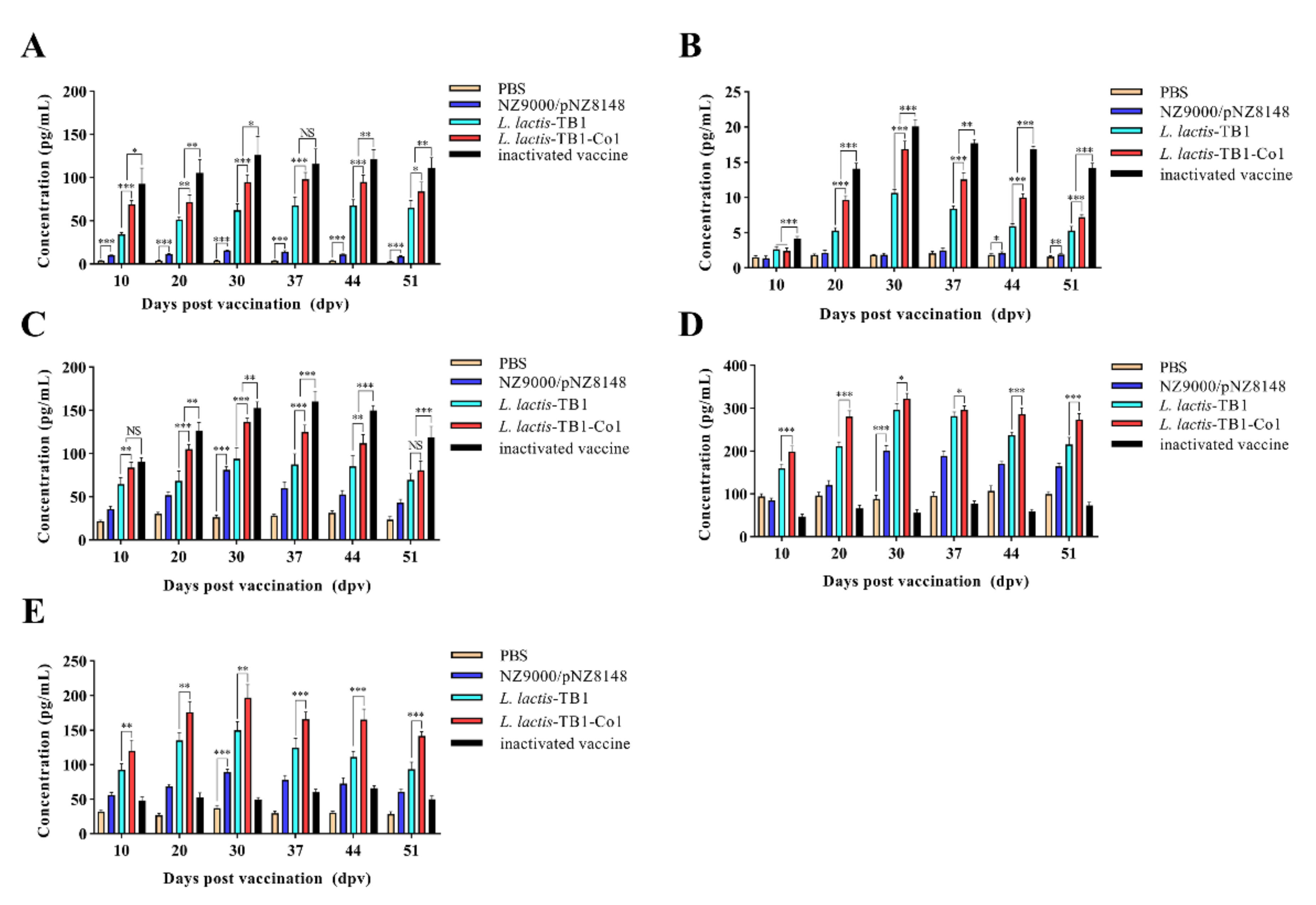
Given that Th cell epitopes contained in recombinant multi-epitope proteins have been shown to upregulate the expression of some cytokines to enhance cellular immune responses, levels of IL-2, IFN-γ, IL-4, IL-10 and IL-5 in mice serum samples were assessed by cytokine ELISA kits (Mlbio, Shanghai, China) as described by the manufacturer.
2.11. Determination of T Lymphocyte Subsets
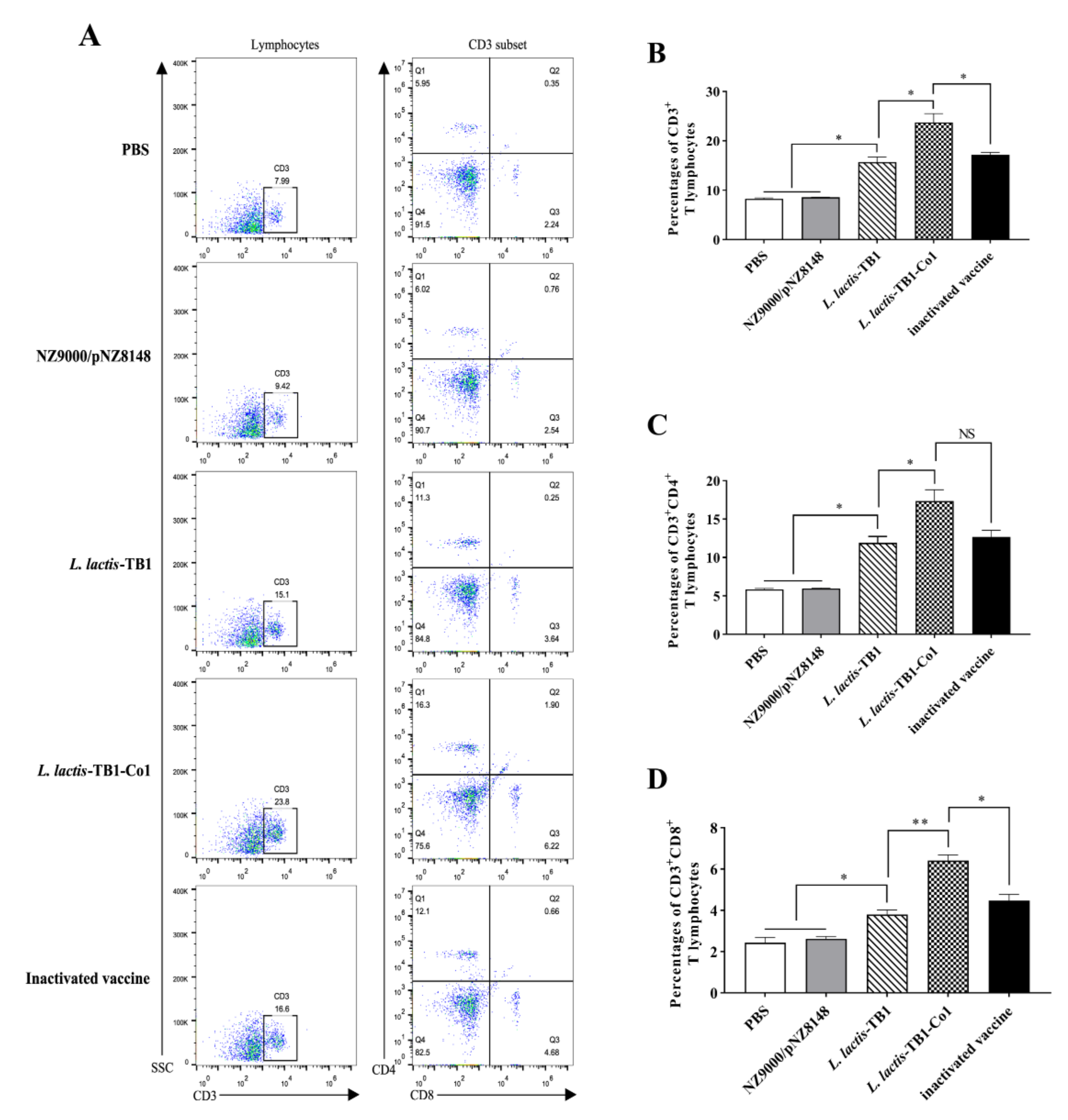
Guinea pig anticoagulated blood samples were obtained on day 30 after initial immunization and stained for 30 min using APC-conjugated rat anti-mouse CD3 antibody, RPE-conjugated rat anti-mouse CD4 antibody and FITC-conjugated rat anti-mouse CD8 antibody (AbD Serotec, Oxford, UK) at room temperature (RT). Evaluation of gated CD3 positive events was carried out for CD3+CD8+ as well as CD3+CD4+ T cells. Flow cytometry (BD Biosciences, San Jose, CA, USA) was conducted, followed by analysis by the FlowJo software (FlowJo_V10).
2.12. Splenic Lymphocyte Proliferation Assays
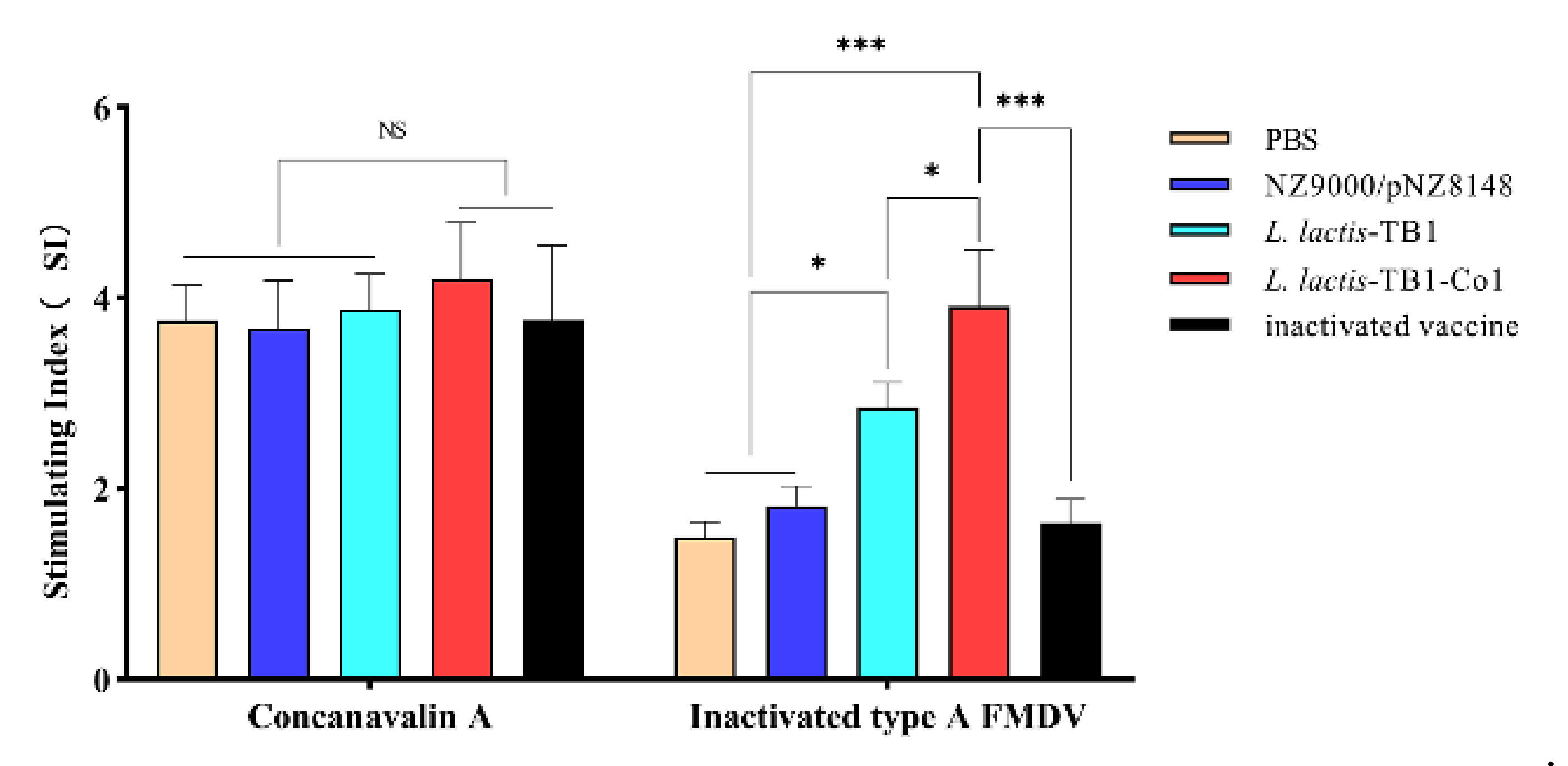
On the 30th day after first immunization of mice, isolation of splenic lymphocytes was performed using a Spleen Lymphocyte Separation Medium (Solarbio, Beijing, China). Assessment of cell proliferation levels was carried out by the MTT Lymphocyte Proliferation Assay Kit (Solarbio, Beijing, China). In brief, suspension of splenic lymphocytes was carried out in RPMI-1640 medium supplemented with FBS (10%; Gibco, Waltham, MA, USA) and 1% penicillin/streptomycin (Gibco, Waltham, MA, USA). Then, they were culivated in 96-well plates (5 × 106 cells per mL and 50 μL/well). Sample stimulation was carried out using 50 μL inactivated FMDV antigens (10 μg/mL; specific antigen stimulation), concanavalin A (ConA, 10 μg/mL) (positive control) and culture medium (negative control), respectively, incubated for 72 h in 5% CO2 at 37 °C. Proliferation responses were detected by an MTT Lymphocyte Proliferation Assay Kit (Solarbio, Beijing, China). Findings are expressed as stimulation indices (SI, ratio of stimulated/unstimulated samples at OD490 nm).
2.13. Statistical Analysis
Data are shown as mean ± SD. GraphPad Prism 7 (LaJolla, CA, USA) was used for statistical analyses. Two-way ANOVA, t tests and multiple t tests were performed to determine significant differences among and between means, respectively. p ≤ 0.05) was significant and p ≤ 0.01 was very significant.
4. Discussion
Traditional inactivated vaccines induce strong humoral immune responses and weak cell-mediated immune responses [
29]. Moreover, it could not prevent viral entry through the mucosa [
30]. Generally, FMDV infections occur at mucosal surfaces, therefore, mucosal immunization using specific multi-epitope antigens can protect against infections. In addition, mucosal IgA was associated with protective effects among FMDV-infected pigs, indicating the necessity for establishment of oral mucosa vaccines against FMDV [
31]. Therefore, oral mucosa vaccines against FMDV using multi-epitope antigens may overcome the limitations associated with injection vaccines, and can significantly activate the first line immune defenses in the gut. Thus, intestinal mucosa immune systems, which forms the first barrier against bacterial, viral and parasitic intestinal infections is being evaluated [
17]. SIgA is very important on mucosal surfaces, therefore, SIgA secretion is vital for efficient antigen-specific mucosal immune responses [
32]. As a first line immune component, SIgA protects via intracellular neutralization, immune exclusion, and antigen excretion. Immune exclusion is mediated via antigenic or bacterial entrapment and suppression of their interactions with epithelial cells [
33].
In mucosal lymphoid tissues, vaccine antigens are essential for generation of antigen-specific SIgA [
34]. In the mucosal epithelium, macromolecule transcytosis occur in M cells via specific or non-specific receptor-associated processes [
35]. This, studies have aimed at evaluating immune strategies that target antigens to M cells to develop efficient oral mucosa vaccines. The interaction between M cell-targeting ligand Co1 and C5aR on M cells is associated with antigen delivery as well as initiation of antigen-specific immune responses [
21]. In oral mucosal immunization, since the antigen is degraded in the gastric juice, the antigen can be presented with a capsule or a protective carrier.
L. lactis was used as a live carrier to present antigenic proteins. Moreover,
L. lactis exerts pro-inflammatory effects and responses in the intestinal mucosa, making it a good oral vaccine [
36]. Induction of TB1 and TB1-Co1 protein expressions in
L. lactis was carried out using the NICE system. Multi-epitope antigen proteins were identified in bacterial lysates and not culture supernatants, implying intracellular expressions in
L. lactis. We found that the Co1-conjugated multi-epitope antigen, TB1 of FMDV, had effective interactions with M cells of ileal PPs relative to TB1 alone minus Co1 ligand conjugation. In addition, in this study, the oral mucosal vaccine,
L. lactis-TB1-Co1, against FMDV was developed and experimented in mice and guinea pigs. We found that
L. lactis-TB1-Co1 triggered systemic, mucosal, as well as cell-mediated immune reactions.
Mucosal immune reactions, especially IgA, are efficient against mucosal surface pathogens [
37].
L. lactis-TB1-Co1 administration in mice led to elevations in SIgA titers in intestinal and lung lavage fluids. However, intestinal SIgA levels were elevated, compared to lungs. In addition, IgG antibodies revealed that
L. lactis-TB1-Co1 triggered high-level systemic immune responses. Therefore, mucosal immunization can initiate mucosal as well as circulating antibody responses. This outcome can be attributed to various reasons. One, there was an abundance of M cells and dendritic cells in the follicle epitheliums of gut-related lymphoid tissues [
38]. These cells transported antigenic proteins, TB1-Co1, from gut lumens to systemic immune cells, which stimulated systemic immune responses. Two, common mucosal immune system (CMIS) has an integrated cross-communication pathway of lymphoid tissues that are made up of inductive as well as effector sites for host protection against pathogens. Major CMIS effector molecules are IgA, IgG antibodies as well as cytokines. These effector molecules can home to system-related lymphoreticular tissues via blood circulation [
39]. Comparable findings were obtained in guinea pigs. Moreover, we found that IgG antibody secretions occurred later than SIgA, which may play an important role against pathogen invasion from mucosal surfaces. However, the involved mechanisms are uncertain. As previously reported [
40], intramuscular inoculation with inactivated vaccines, regardless of the assay, does not induce IgA responses, consistent with our findings. This highlights the need to understand immune responses to this virus, so as to develop new, rapid-action vaccines for FMDV.
Various pathogens are capable of inducing cytokine secretion by CD4
+ T cells. Cytokines, which are small-molecule proteins with many biological activities, are produced by both immune as well as some non-immune cells [
41]. As cell signaling molecules, they regulate immune responses, participate in immune cell differentiation and development, mediate inflammatory responses, stimulate hematopoiesis, and participate in tissue repairs. Auxiliary signals for B cell proliferation as well as differentiation are obtained from CD4
+ T cells, while CD8
+ T cells are cytotoxic killer cells and innate immune regulators [
42]. Effector CD4
+ T cells include Th1 as well as Th2-specific responses. IL-2 and IFN-γ, which are produced by Th1 cells, enhance the activation as well as proliferation of CTL, macrophages, and NK cells, promote pathogen cytotoxicity and phagocytosis, and enhance CD8
+ T cell and innate immune responses [
43]. IL-10 and IL-4, which are produced by Th2 cells enhance B cell proliferation as well as differentiation to plasma cells, thereby initiating antibody secretion and promoting humoral immune responses. Th1 responses, such as IFN-γ and IL-2 secretion are very effective against intracellular pathogens. Other Th2 responses, including IL-5, IL-4, and IL-10 secretions can efficiently eliminate pathogens from body fluids. In this study,
L. lactis-TB1-Co1 enhanced Th1- as well as Th2-type responses, consistent with other studies [
44,
45]. This is due to the fact that Th cell epitopes are vital in the promotion of both humoral as well as cell immune responses by stimulating T cells [
26,
46]. T cell-secreted cytokines can also enhance the proliferation as well as differentiation of B cells into plasma cells [
47]. Cellular immunity effectively protects against FMD [
48]. Cellular immune responses are assessed by lymphocyte proliferation
in vitro. Con A stimulates T cell proliferation. Our results demonstrated that
L. lactis-TB1-Co1 regulated specific T lymphocyte proliferative responses, and secreted cytokines to enhance the abundance of CD4
+ and CD8
+ T lymphocytes in mice.
Challenge test results showed that guinea pigs immunized with
L. lactis-TB1-Co1 exhibited 60% protection rates. These findings demonstrate that, to some extent, oral immunization with
L. lactis-TB1-Co1 protected against FMDV. To promote the effects of immunization of recombinant
L. lactis, we used an efficient oral mucosal adjuvant, CpG-ODN [
49,
50]. However, it was found that guinea pigs orally vaccinated with
L. lactis-TB1-Co1 plus CpG-ODN did not exhibit significant protection rates. In this regard, we speculate that there are two reasons for the situation. On the one hand, it is possible that the dose of CPG ODN used is low in this experiment, which makes it unable to effectively activate B cells, monocytes, macrophages and dendritic cells to secrete antibodies, various cytokines and chemokines [
50]. On the other hand, it may not be effective in guinea pigs due to differences in species. For higher protection effects, double doses of
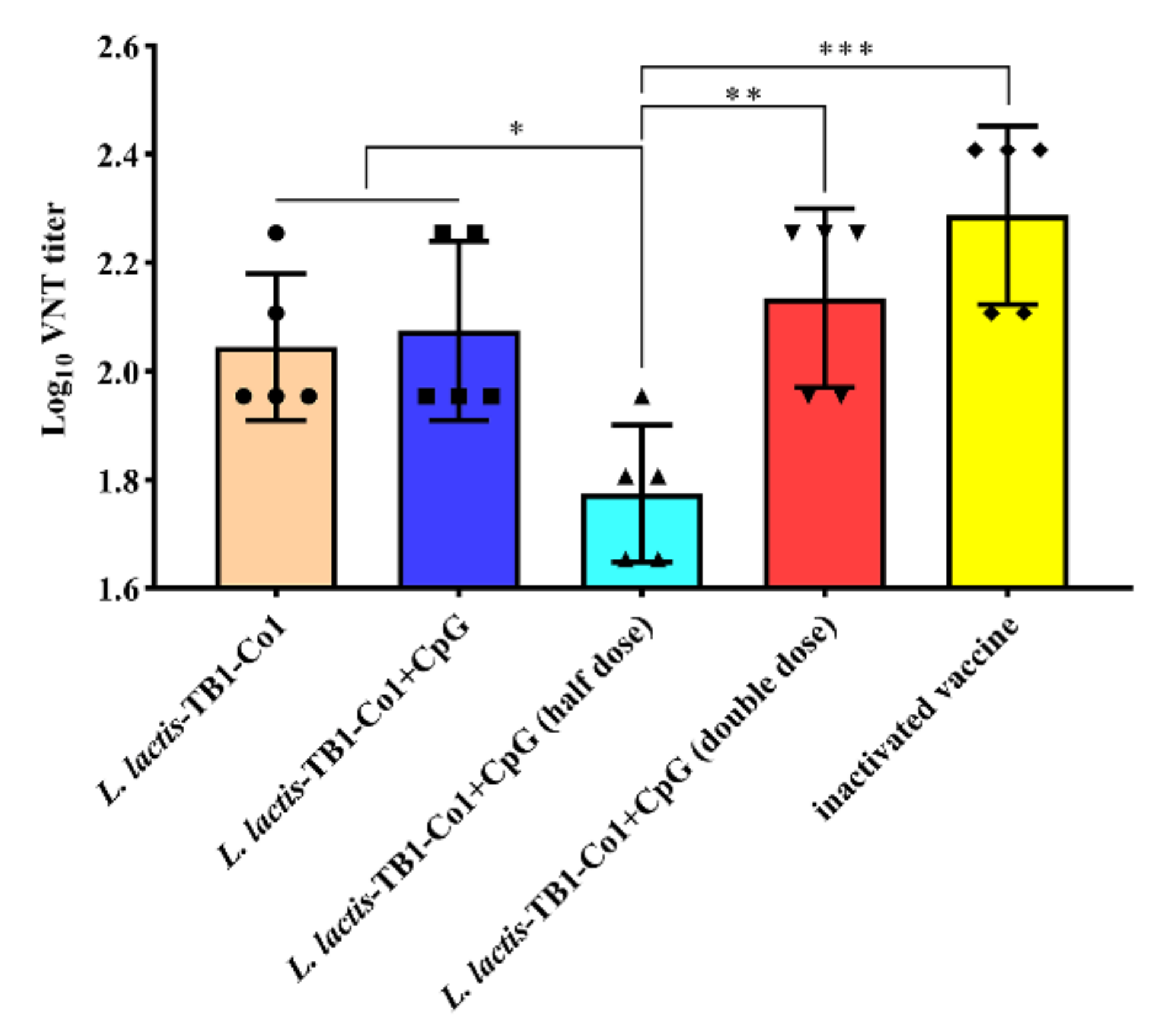
L. lactis-TB1-Co1 were administered, we found a consistent result. Interestingly, half of the dose used did not protect the guinea pigs. We found that protection rates did not increase when the vaccination volume was doubled. This outcome may be due to the ability of M cells to present antigens had reached its limit. However, it inhibited disease severity and delayed the onset of clinical symptoms, relative to negative-control animals. Thus,
L. lactis-TB1-Co1 may have led to the production of specific neutralizing antibody titers. As previously reported [
26,
51,
52], there was not a strong association between neutralizing antibodies and protection. It has been proven that interferon-mediated cellular immunity can protect pigs from FMDV infection [
53,
54]. Thus, to evaluate the efficacy of vaccines for FMD, in addition to in vitro neutralizing activities, cellular immune responses, antibody affinities and other antibody responses, should be considered [
26].
As an exploratory research, this study has several limitations. First, since no purified multi-epitope antigen proteins were obtained, in vitro and in vivo experiments on antigen uptake were not performed. Second, there is no currently unified gold standard for evaluating mucosal immunity. Therefore, we adopted traditional challenge methods, which may weaken protection efficiencies of this oral mucosal vaccine. We postulate that better experimental results may be obtained if the challenge is carried out by oral or aerosol methods. In future studies, we will adopt safe and cheap mucosal adjuvants to achieve better experimental results. More studies are needed to assess the cross-protection that is mediated by oral mucosal vaccines against other FMDV serotype A lineages in guinea pigs and swine, which will better help us understand the effectiveness of the vaccine.
In conclusion, we have demonstrated that M cell-targeting ligand Co1 is an important adjuvant for FMDV oral mucosal vaccines. It is important in efficient delivery of ligand-conjugated multi-epitope antigens to mucosal immune components and effective initiation of mucosal and systemic immune responses in mice. Moreover, we showed that CpG-ODN, as the oral mucosal vaccine adjuvant, could enhance mucosal immune responses and systemic immune responses of guinea pigs. Therefore, although oral vaccination with L. lactis-TB1-Co1 could, to some extent, protect against FMDV challenge, it suppressed disease severity and delayed clinical symptoms, implying that L. lactis-TB1-Co1 has potential as a novel oral mucosal vaccine for FMD.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}