
Effect of Serial Systemic and Intratumoral Injections of Oncolytic ZIKVBR in Mice Bearing Embryonal CNS Tumors

 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Human and Animal Samples
2.2. Cell Lines and Culture
2.3. 3D Brain Model and Co-Culture Assay
2.4. In Vivo Injection Assay
2.5. Orthotopic Metastatic Xenograft Animal Model
2.6. ZIKV Strain and Viral Detection
2.7. Histopathology and Immunofluorescence
2.8. Real-Time Quantitative PCR Array
2.9. Image and Statistical Analysis
3. Results
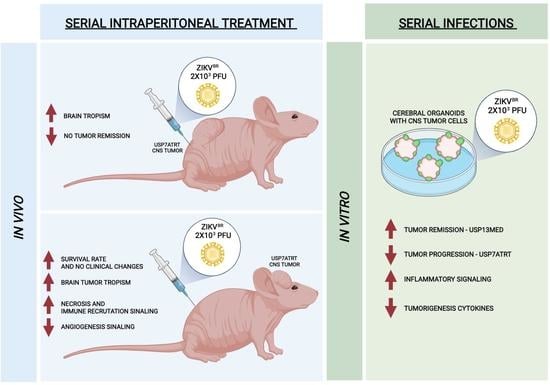
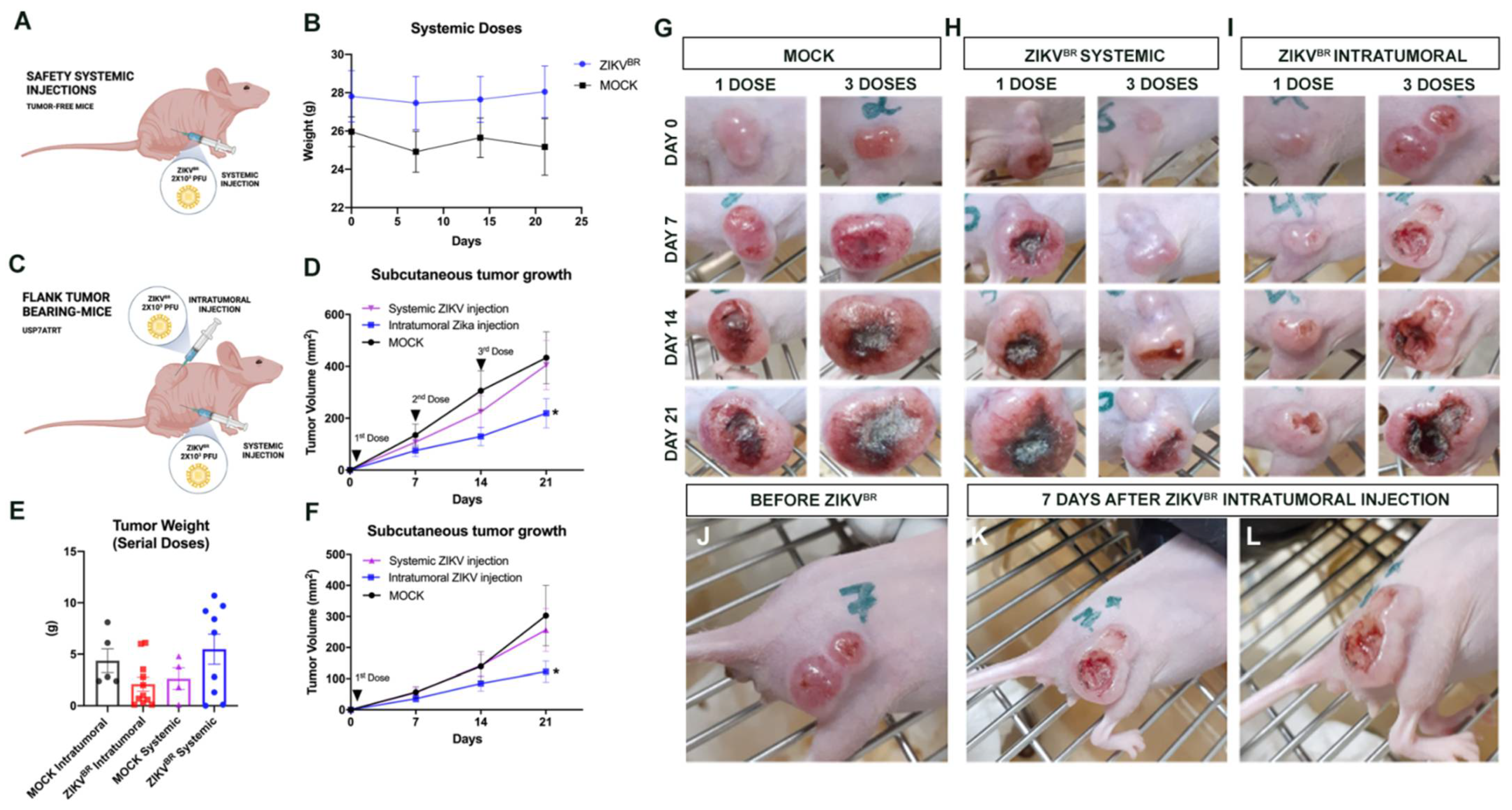
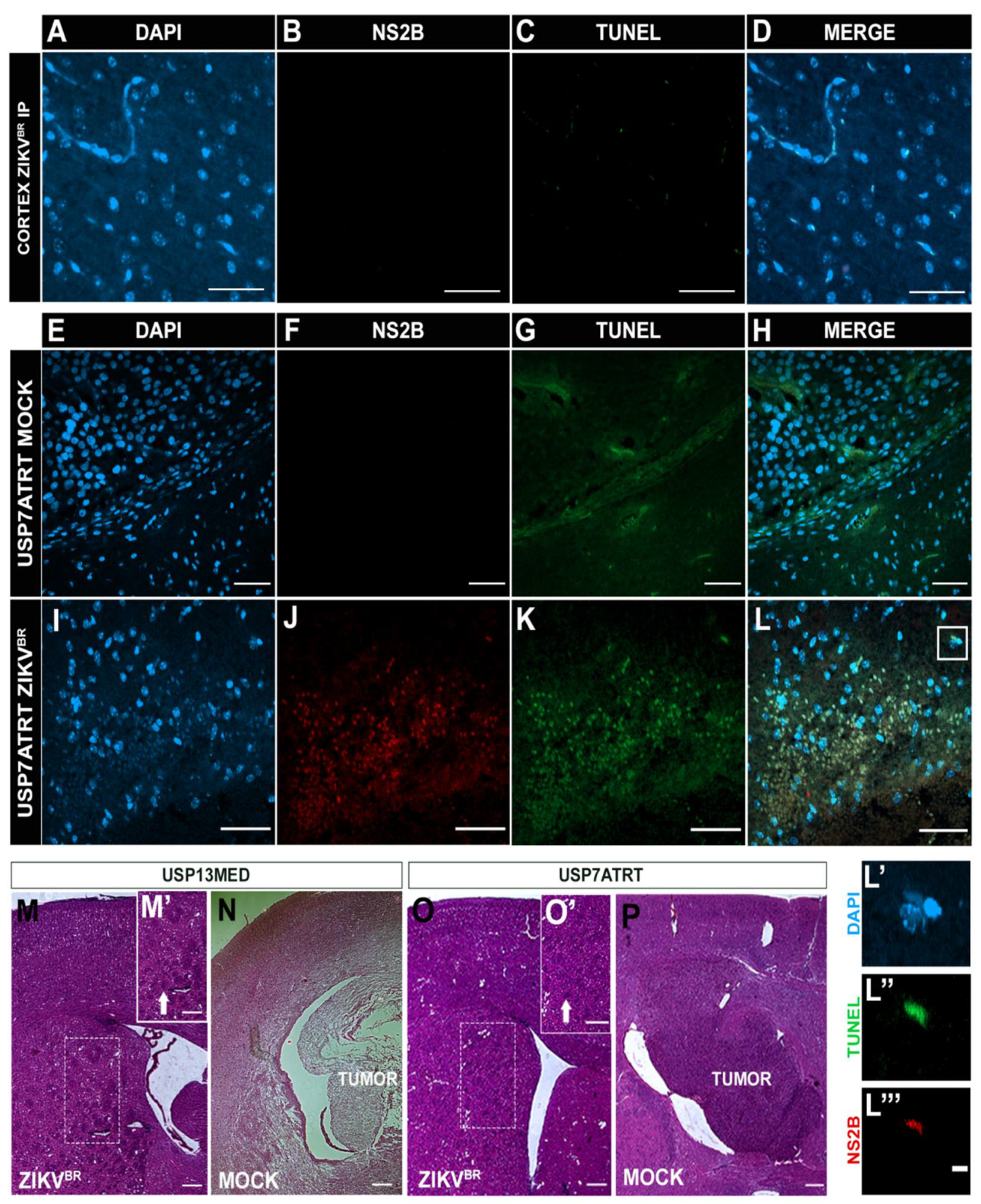
3.1. Evaluation of Viral Safety in Balb/c Nudemice after Serial Systemic and Intracerebroventricular ZIKVBR Infections
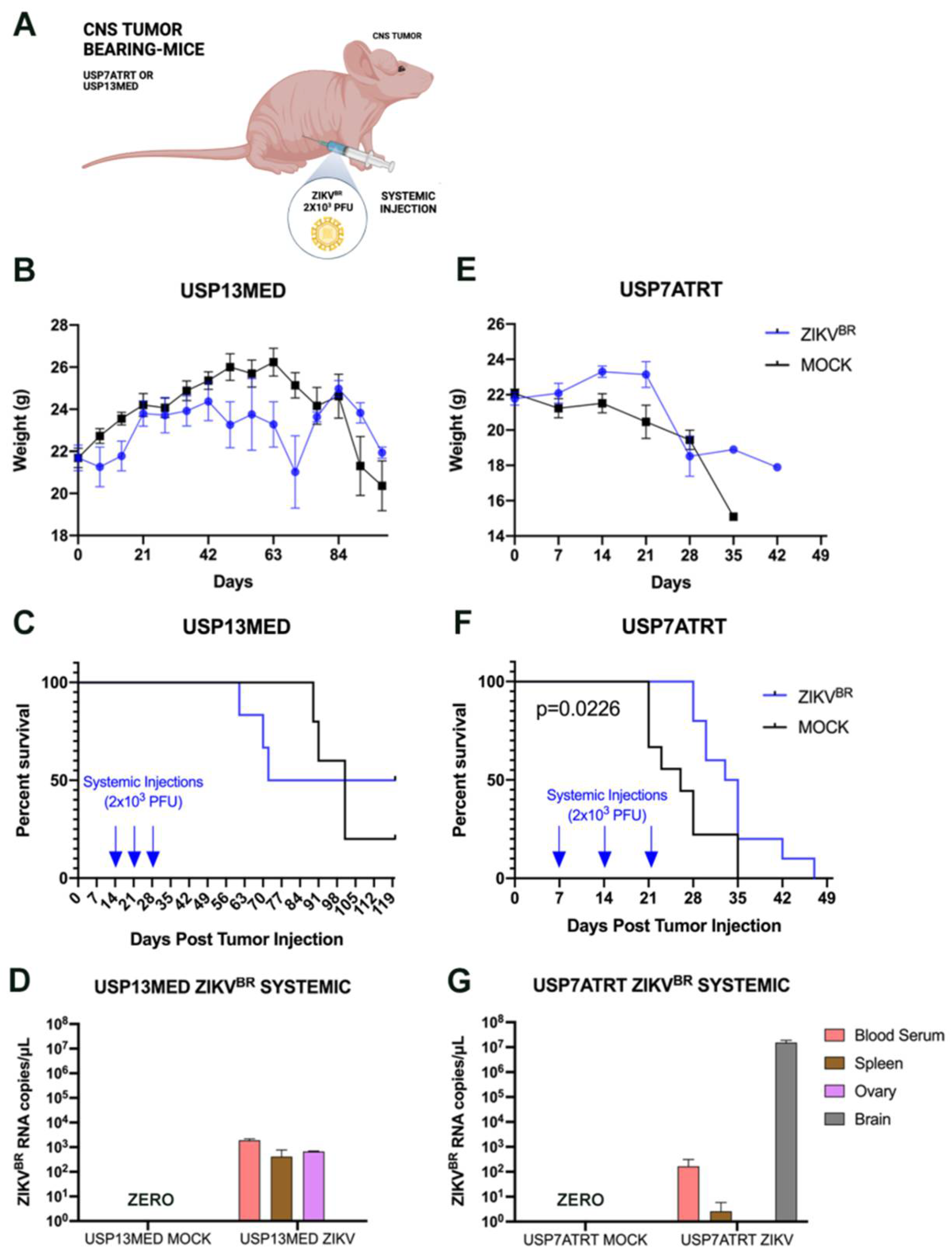
3.2. Evaluation of Viral Tropism in Mice Bearing Subcutaneous Embryonal CNS Tumor after Systemic ZIKVBR Infections
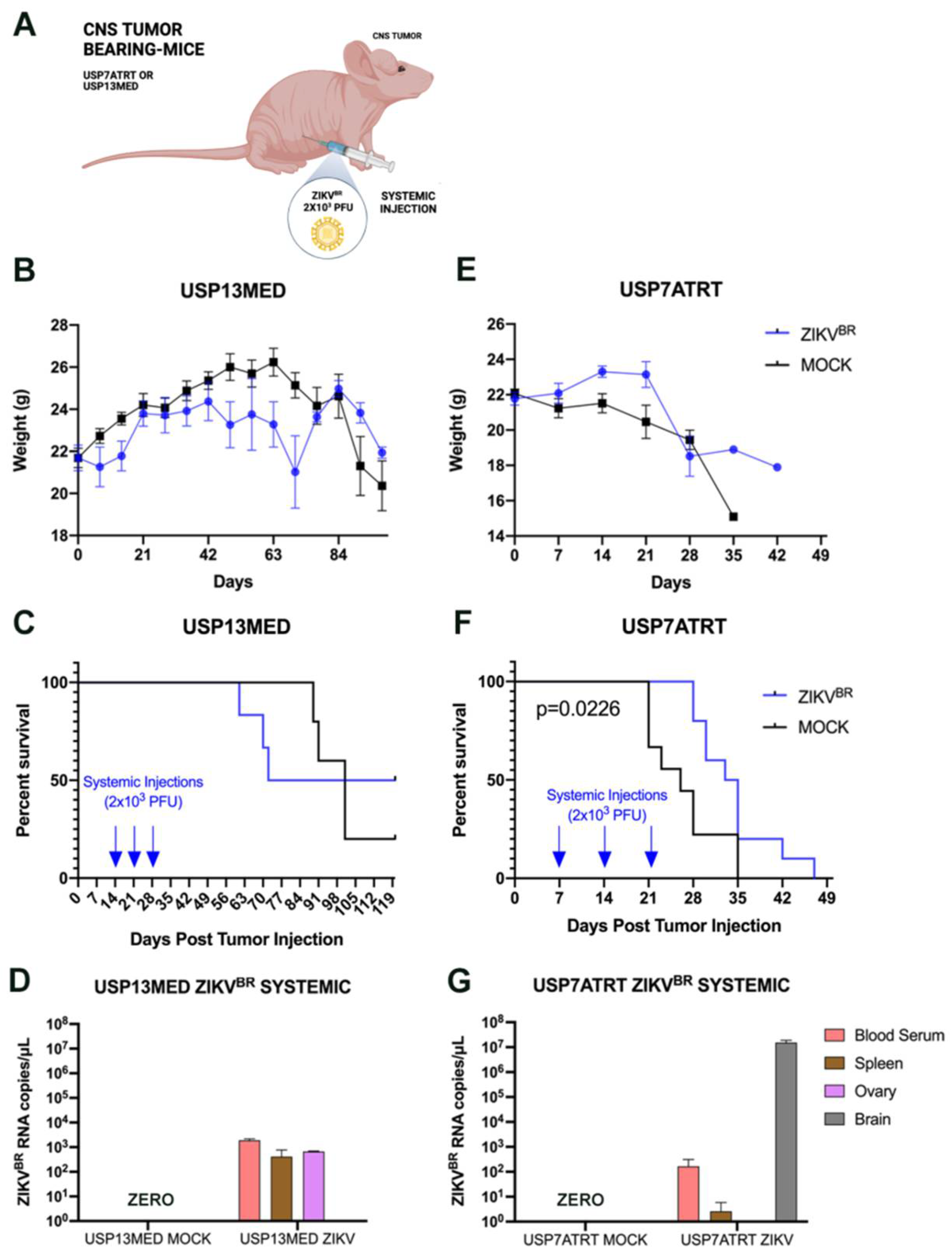
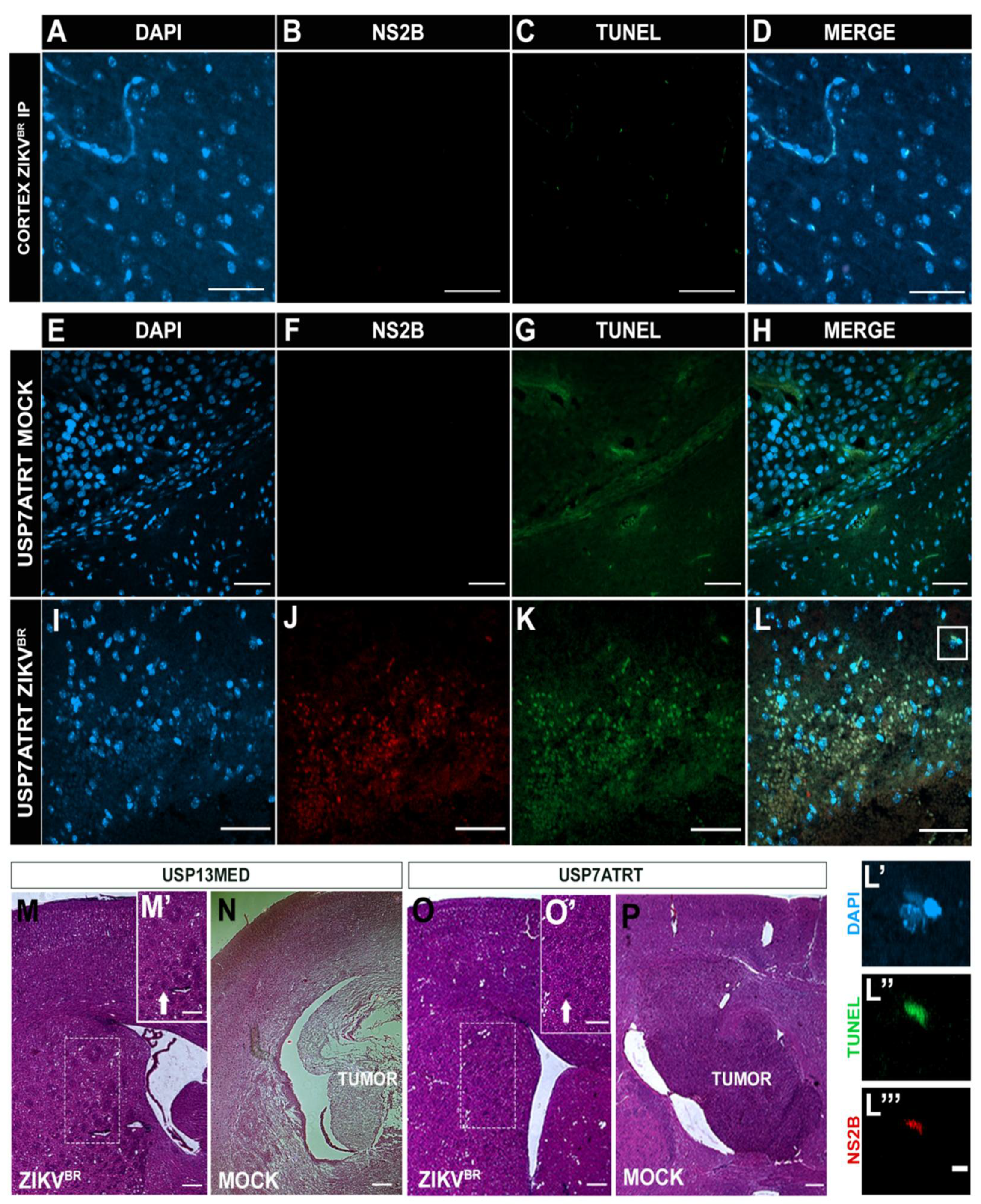
3.3. ZIKVBR Oncolytic Effect in Intracranial Tumor-Bearing Mice Varying ZIKVBR Internalization
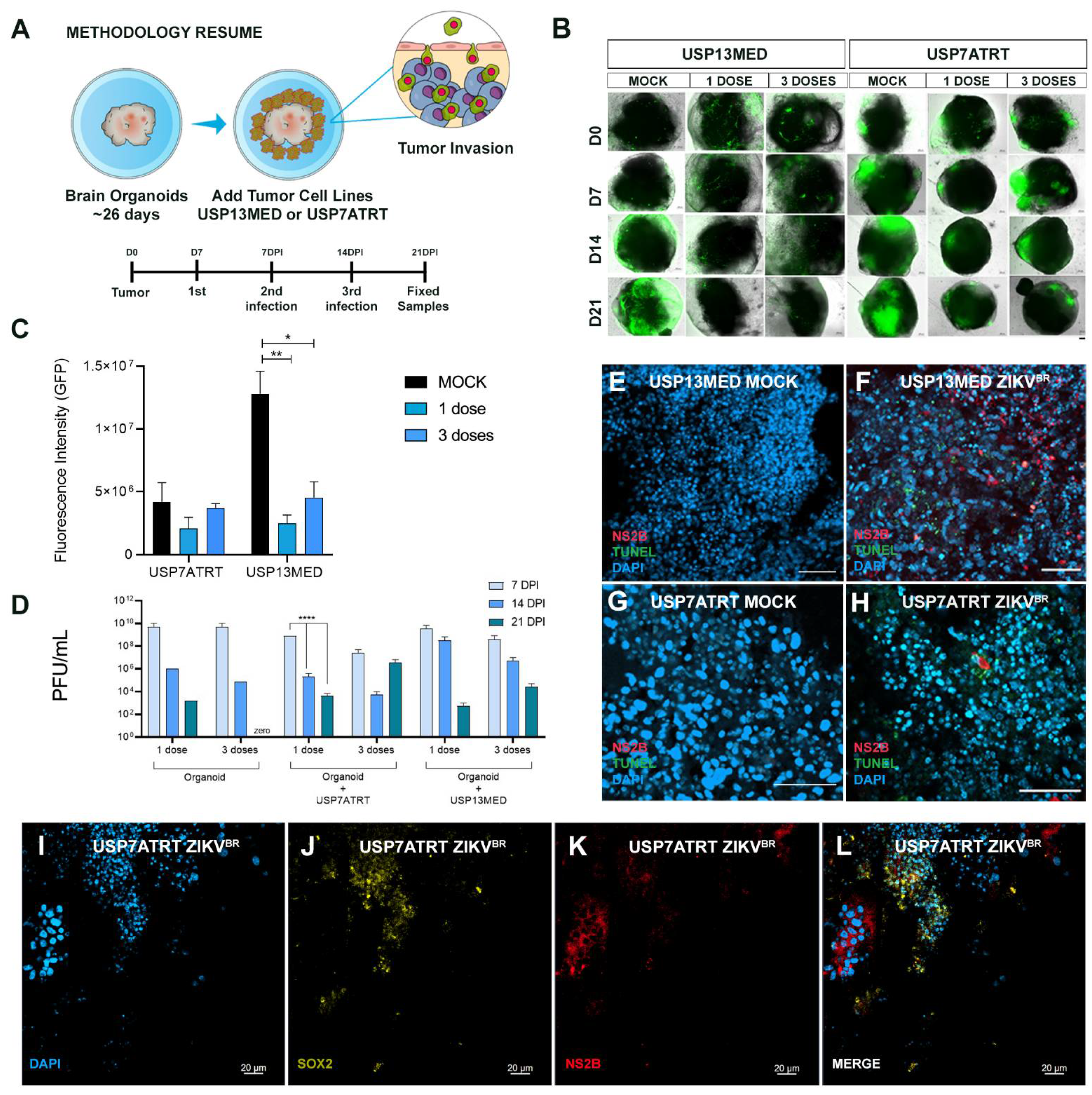
3.4. ZIKVBR Infection Impairs Tumor Cells Spread in Brain Organoids In Vitro
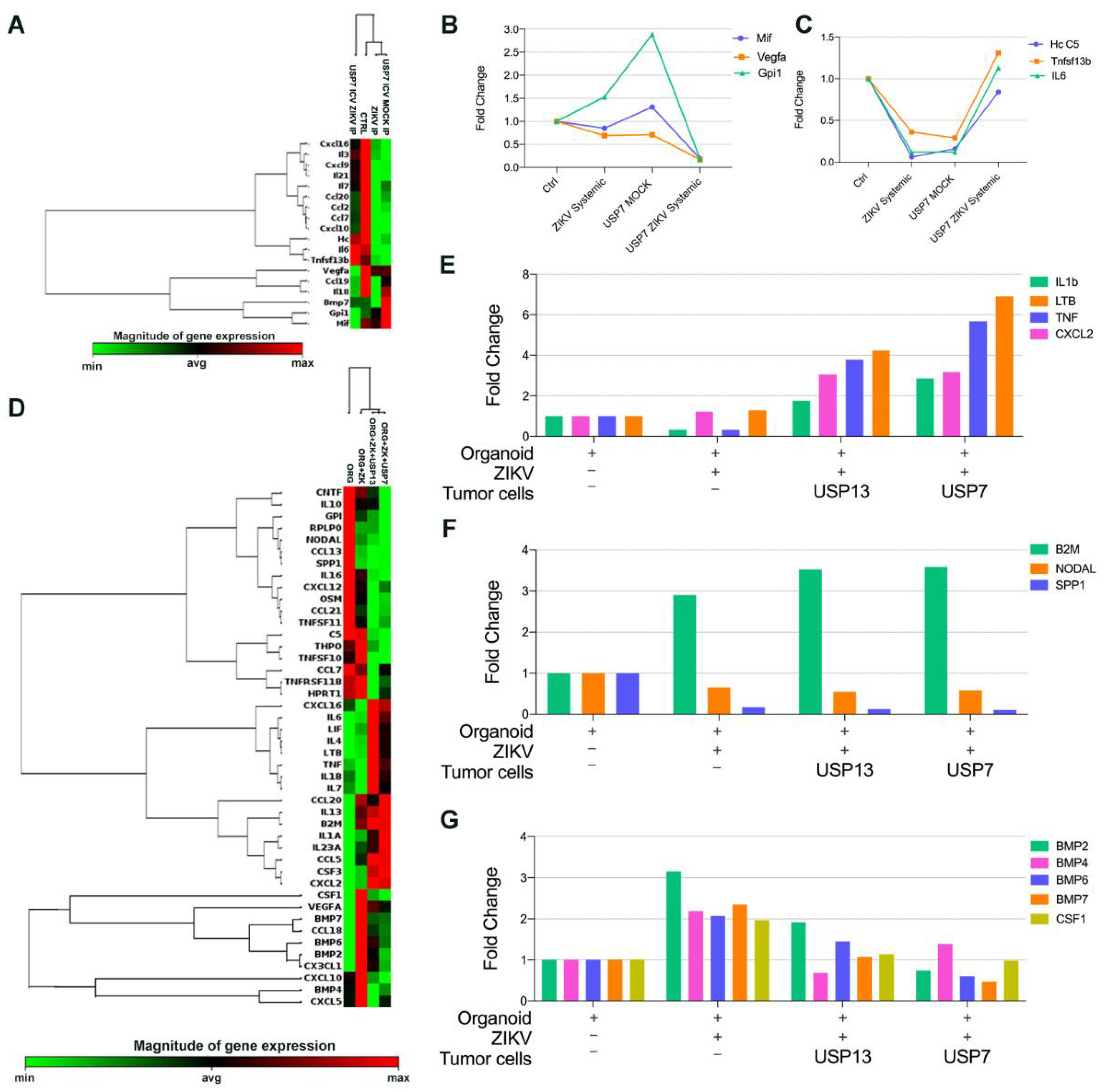
3.5. ZIKVBR Infection in Tumor Cells Changes the Cytokines and Chemokines Expression In Vitro and In Vivo
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Su, K.Y.; Balasubramaniam, V.R.M.T. Zika Virus as Oncolytic Therapy for Brain Cancer: Myth or Reality? Front. Microbiol. 2019, 10, 2715. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Mesci, P.; Bernatchez, J.A.; Gimple, R.C.; Wang, X.; Schafer, S.T.; Wettersten, H.I.; Beck, S.; Clark, A.E.; Wu, Q. Zika Virus Targets Glioblastoma Stem Cells through a SOX2-Integrin αvβ5 Axis. Cell Stem Cell 2020, 26, 187–204. [Google Scholar] [CrossRef] [PubMed]
- Kaid, C.; Goulart, E.; Caires-Júnior, L.C.; Araujo, B.H.S.; Soares-Schanoski, A.; Bueno, H.M.S.; Telles-Silva, K.A.; Astray, R.M.; Assoni, A.F.; Júnior, A.F.R. Zika Virus Selectively Kills Aggressive Human Embryonal CNS Tumor Cells In Vitro and In Vivo. Cancer Res. 2018, 78, 3363–3374. [Google Scholar] [CrossRef] [Green Version]
- Kaid, C.; dos Santos Madi, R.A.; Astray, R.; Goulart, E.; Caires-Junior, L.C.; Mitsugi, T.G.; Moreno, A.C.R.; Castro-Amarante, M.F.; Pereira, L.R.; Porchia, B.F.M.M. Safety, Tumor Reduction, and Clinical Impact of Zika Virus Injection in Dogs with Advanced-Stage Brain Tumors. Mol. Ther. 2020, 28, 1276–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, S.; Mazzoccoli, L.; Jash, A.; Govero, J.; Bais, S.S.; Hu, T.; Fontes-Garfias, C.R.; Shan, C.; Okada, H.; Shresta, S. Zika virus oncolytic activity requires CD8+ T cells and is boosted by immune checkpoint blockade. JCI Insight 2021, 6, e144619. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.E.; Kobayashi, G.S.; Musso, C.M.; Allen, M.; Ishiy, F.A.A.; De Caires, L.C.; Goulart, E.; Griesi-Oliveira, K.; Zechi-Ceide, R.M.; Richieri-Costa, A. EIF4A3 Deficient Human IPSCs and Mouse Models Demonstrate Neural Crest Defects That Underlie Richieri-Costa-Pereira Syndrome. Hum. Mol. Genet. 2017, 26, 2177–2191. [Google Scholar] [CrossRef] [Green Version]
- da Silva, P.B.G.; Rodini, C.O.; Kaid, C.; Nakahata, A.M.; Pereira, M.C.L.; Matushita, H.; da Costa, S.S.; Okamoto, O.K. Establishment of a novel human medulloblastoma cell line characterized by highly aggressive stem-like cells. Cytotechnology 2015, 68, 1545–1560. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, M.A.; Knoblich, J.A. Generation of Cerebral Organoids from Human Pluripotent Stem Cells. Nat. Protoc. 2014, 9, 2329. [Google Scholar] [CrossRef] [Green Version]
- Studebaker, A.W.; Hutzen, B.; Pierson, C.R.; Russell, S.J.; Galanis, E.; Raffel, C. Oncolytic measles virus prolongs survival in a murine model of cerebral spinal fluid–disseminated medulloblastoma. Neuro-Oncology 2012, 14, 459–470. [Google Scholar] [CrossRef]
- Mlakar, J.; Korva, M.; Tul, N.; Popović, M.; Poljšak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Resman, R.K.; Vesnaver, V.T.; Fabjan, V.V. Zika Virus Associated with Microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Jevtić, P.; Levy, D.L. Mechanisms of nuclear size regulation in model systems and cancer. Cancer Biol. Nucl. Envel. 2014, 773, 537–569. [Google Scholar] [CrossRef]
- Cugola, F.R.; Fernandes, I.R.; Russo, F.B.; Freitas, B.C.; Dias, J.L.M.; Guimarães, K.P.; Benazzato, C.; Almeida, N.; Pignatari, G.C.; Romero, S. The Brazilian Zika virus strain causes birth defects in experimental models. Nature 2016, 534, 267–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caires-Júnior, L.C.; Goulart, E.; Melo, U.S.; Araujo, B.H.S.; Alvizi, L.; Soares-Schanoski, A.; Oliveira, D.F.; de Kobayashi, G.S.; Griesi-Oliveira, K.; Musso, C.M. Discordant congenital Zika syndrome twins show differential in vitro viral susceptibility of neural progenitor cells. Nat. Commun. 2018, 9, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicenti, I.; Boccuto, A.; Giannini, A.; Dragoni, F.; Saladini, F.; Zazzi, M. Comparative analysis of different cell systems for Zika virus (ZIKV) propagation and evaluation of anti-ZIKV compounds in vitro. Virus Res. 2018, 244, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Funasaka, T.; Hogan, V.; Raz, A. Phosphoglucose Isomerase/Autocrine Motility Factor Mediates Epithelial and Mesenchymal Phenotype Conversions in Breast Cancer. Cancer Res. 2009, 69, 5349–5356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.-C.; Kung, M.-L.; Hu, T.-H.; Chen, H.-Y.; Wu, J.-C.; Kuo, H.-M.; Tsai, H.-E.; Lin, Y.-W.; Wen, Z.-H.; Liu, J.-K. Hepatoma-derived growth factor regulates breast cancer cell invasion by modulating epithelial-mesenchymal transition. J. Pathol. 2012, 228, 158–169. [Google Scholar] [CrossRef]
- Le Tourneau, C.; Stathis, A.; Vidal, L.; Moore, M.J.; Siu, L.L. Choice of Starting Dose for Molecularly Targeted Agents Evaluated in First-in-Human Phase I Cancer Clinical Trials. J. Clin. Oncol. 2010, 28, 1401–1407. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.; Hollidge, B.; Daye, S.; Zeng, X.; Blancett, C.; Kuszpit, K.; Bocan, T.; Koehler, J.W.; Coyne, S.; Minogue, T. Neuropathogenesis of Zika Virus in a Highly Susceptible Immunocompetent Mouse Model after Antibody Blockade of Type I Interferon. PLOS Negl. Trop. Dis. 2017, 11, e0005296. [Google Scholar] [CrossRef] [Green Version]
- Musso, D.; Gubler, D.J. Zika virus. Clin. Microbiol. Rev. 2016, 29, 487–524. [Google Scholar] [CrossRef] [Green Version]
- Dix, A.R.; Brooks, W.H.; Roszman, T.L.; Morford, L.A. Immune defects observed in patients with primary malignant brain tumors. J. Neuroimmunol. 1999, 100, 216–232. [Google Scholar] [CrossRef]
- Yockey, L.J.; Jurado, K.A.; Arora, N.; Millet, A.; Rakib, T.; Milano, K.M.; Hastings, A.K.; Fikrig, E.; Kong, Y.; Horvath, T.L. Type I interferons instigate fetal demise after Zika virus infection. Sci. Immunol. 2018, 3, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Wang, X.; Shen, J.; Yao, J. Macrophage migration inhibitory factor in the pathogenesis of leukemia (Review). Int. J. Oncol. 2021, 59, 62. [Google Scholar] [CrossRef] [PubMed]
- Alban, T.J.; Bayik, D.; Otvos, B.; Rabljenovic, A.; Leng, L.; Jia-Shiun, L.; Roversi, G.; Lauko, A.; Momin, A.A.; Mohammadi, A.M. Glioblastoma Myeloid-Derived Suppressor Cell Subsets Express Differential Macrophage Migration Inhibitory Factor Receptor Profiles That Can Be Targeted to Reduce Immune Suppression. Front. Immunol. 2020, 11, 1191. [Google Scholar] [CrossRef] [PubMed]
- Garancher, A.; Suzuki, H.; Haricharan, S.; Chau, L.Q.; Masihi, M.B.; Rusert, J.M.; Norris, P.S.; Carrette, F.; Romero, M.M.; Morrissy, S.A. Tumor Necrosis Factor Overcomes Immune Evasion in p53-Mutant Medulloblastoma. Nat. Neurosci. 2020, 23, 842. [Google Scholar] [CrossRef]
- Hayes, K.; Kim, Y.-K.; Pera, M.F. A case for revisiting Nodal signaling in human pluripotent stem cells. Stem Cells 2021, 39, 1137–1144. [Google Scholar] [CrossRef]
- Gámez, B.; Rodriguez-Carballo, E.; Ventura, F. BMP Signaling in Telencephalic Neural Cell Specification and Maturation. Front. Cell. Neurosci. 2013, 7, 87. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, R.O.; Granha, I.; Ferreira, R.S.; Bueno, H.d.S.; Okamoto, O.K.; Kaid, C.; Zatz, M. Effect of Serial Systemic and Intratumoral Injections of Oncolytic ZIKVBR in Mice Bearing Embryonal CNS Tumors. Viruses 2021, 13, 2103. https://doi.org/10.3390/v13102103
Ferreira RO, Granha I, Ferreira RS, Bueno HdS, Okamoto OK, Kaid C, Zatz M. Effect of Serial Systemic and Intratumoral Injections of Oncolytic ZIKVBR in Mice Bearing Embryonal CNS Tumors. Viruses. 2021; 13(10):2103. https://doi.org/10.3390/v13102103
Chicago/Turabian StyleFerreira, Raiane Oliveira, Isabela Granha, Rodolfo Sanches Ferreira, Heloisa de Siqueira Bueno, Oswaldo Keith Okamoto, Carolini Kaid, and Mayana Zatz. 2021. "Effect of Serial Systemic and Intratumoral Injections of Oncolytic ZIKVBR in Mice Bearing Embryonal CNS Tumors" Viruses 13, no. 10: 2103. https://doi.org/10.3390/v13102103
APA StyleFerreira, R. O., Granha, I., Ferreira, R. S., Bueno, H. d. S., Okamoto, O. K., Kaid, C., & Zatz, M. (2021). Effect of Serial Systemic and Intratumoral Injections of Oncolytic ZIKVBR in Mice Bearing Embryonal CNS Tumors. Viruses, 13(10), 2103. https://doi.org/10.3390/v13102103