
Severe Stunting Symptoms upon Nepovirus Infection Are Reminiscent of a Chronic Hypersensitive-like Response in a Perennial Woody Fruit Crop

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material and Virus Inoculation
2.2. Symptom Scoring
2.3. Detection of GFLV
2.4. RNA Extraction and RNA-Sequencing (RNA-Seq)
2.5. Sample Infection Status and GFLV Diversity
2.6. Differential Gene Expression (DEG) Analysis
2.7. Reverse Transcription Quantitative PCR (RT-qPCR)
2.8. Metabolomic Analyses
3. Results
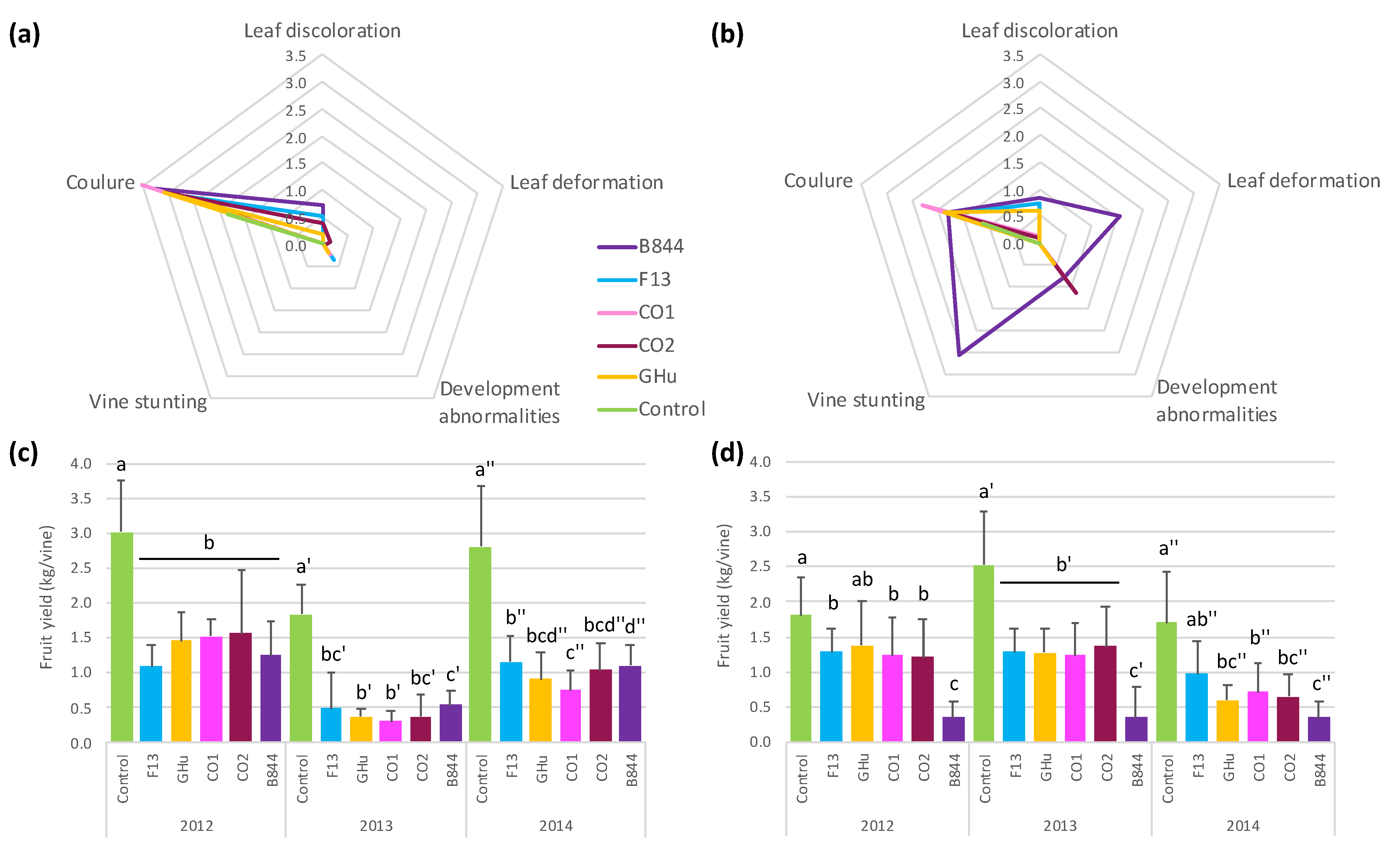
3.1. Differential Symptoms in Vitis vinifera cv. Gewurztraminer Infected by Five Different GFLV Strains
3.2. RNA-Sequencing and Read Mapping on Virus and Grapevine Genomes
3.2.1. Infection Status of the Plant Samples
3.2.2. Grapevine Transcriptome Analysis
- Mapping to the grapevine reference genome.
- Gw genes differentially expressed during GFLV-F13 and GFLV-B844 infections.
- Pathways differently affected by the two GFLV strains.
- Validation of the RNA-Seq differential genes expression by RT-qPCR.
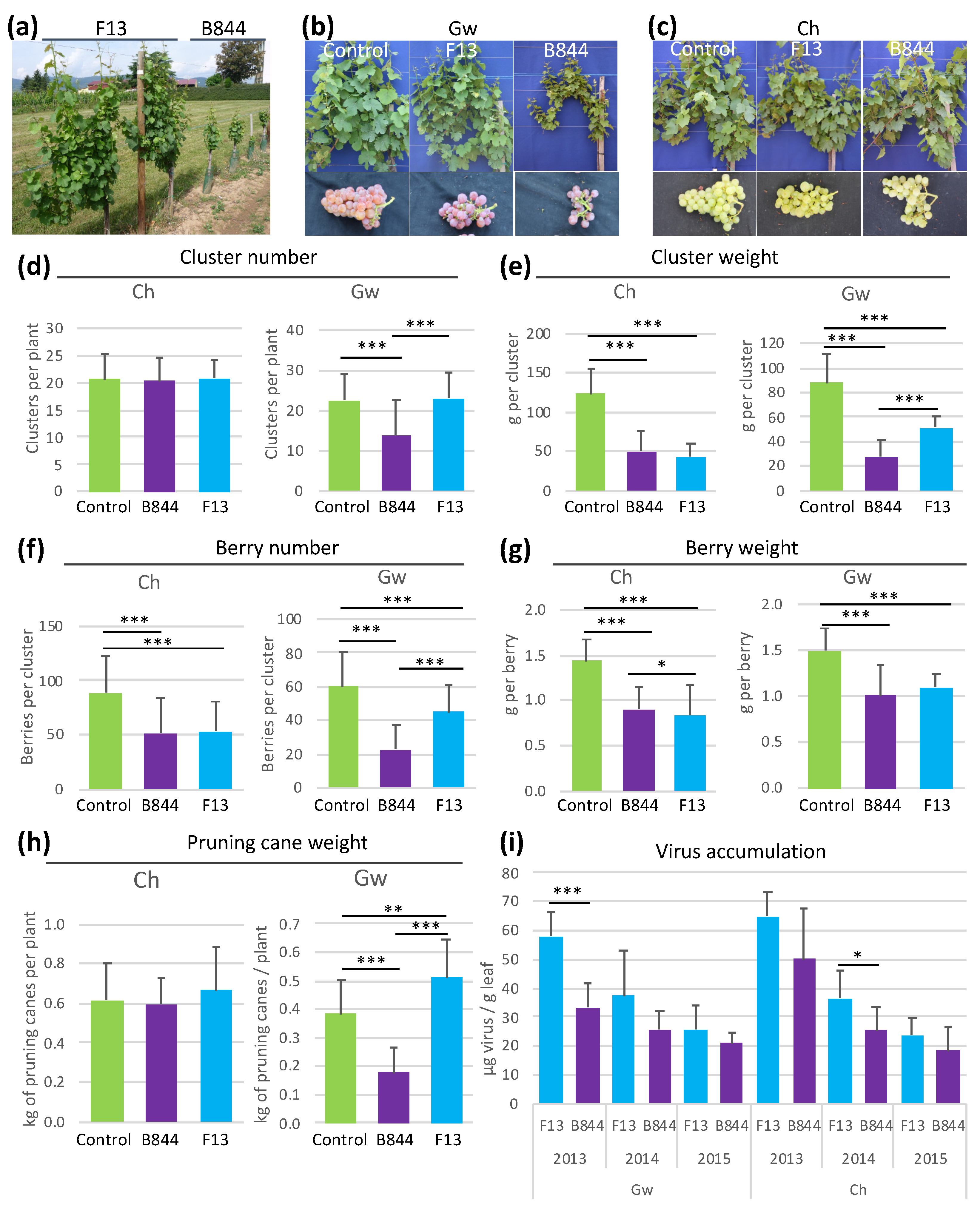
3.3. Biological Validation of an HR-Like Pathway Activation by GFLV-B844 in Gw Vines
Expression Analysis of Candidate Genes in Relation to PLANT Defense Pathways
- Metabolomic analyses.
- Virus restriction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AFC1 | Serine/threonine-protein kinase afc1 |
| AGAL2 | Alpha galactosidase |
| AIL6 | Ap2-like ethylene-responsive transcription factor ail6 |
| APL1 | ADP-glucose pyrophosphorylase large subunit |
| BCP | Blue copper protein |
| CDT1A | cdt1-like protein chloroplastic-like |
| Ch | Chardonnay |
| CYP94 | cytochrome p450 CYP94B3 |
| ERF4 | Ethylene Response Factor 4 |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GFLV | Grapevine fanleaf virus |
| GPTA6 | Glycerol-3-phosphate acyltransferase 6 |
| GRP | Glycine rich protein |
| GRSPaV | Grapevine rupestris stem pitting-associated virus |
| Gw | Gewurztraminer |
| GYSVd | Grapevine yellow speckle viroid |
| HR | Hypersensitive reaction |
| HSVd | Hop stunt viroid |
| HTS | High throughput sequencing |
| KCS5 | 3-ketoacyl- synthase |
| NA | Not annotated |
| NDF6 | NAD(P)H dehydrogenase complex |
| NEMF1 | Nuclear export mediator factor nemf-like |
| NFYC4 | Nuclear transcription factor y subunit c-4 |
| NQO1 | Quinone oxidoreductase 1 |
| PDF1 | Protodermal factor |
| PDF2 | Defensin-like protein 6-like |
| PIN5 | Auxin efflux carrier component |
| PP2A | Serine/threonine protein phosphatase 2A (PP2A) 65 KDa regulatory subunit A |
| PP7 | Serine/threonine-protein phosphatase 7 |
| PR10 | Pathogenesis related protein 10 |
| RD22 | Dehydration-responsive protein rd22 |
| RDR1 | RNA polymerase RNA dependante |
| ROXY2 | Glutaredoxines ROXY2 |
| SYP121 | Syntaxin-related protein nt-syr1 |
| TIP41 | Tip41-like protein |
| VST | Variance-stabilized transformation |
References
- García, J.A.; Pallás, V. Viral factors involved in plant pathogenesis. Curr. Opin. Virol. 2015, 11, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Osterbaan, L.J.; Fuchs, M. Dynamic interactions between plant viruses and their hosts for symptom development. J. Plant Pathol. 2019, 101, 885–895. [Google Scholar] [CrossRef]
- Culver, J.N.; Padmanabhan, M.S. Virus-induced disease: Altering host physiology one interaction at a time. Annu. Rev. Phytopathol. 2007, 45, 221–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallas, V.; García, J.A. How do plant viruses induce disease? Interactions and interference with host components. J. Gen. Virol 2011, 92, 2691–2705. [Google Scholar] [CrossRef]
- Shi, B.; Lin, L.; Wang, S.; Guo, Q.; Zhou, H.; Rong, L.; Li, J.; Peng, J.; Lu, Y.; Zheng, H.; et al. Identification and regulation of host genes related to rice stripe virus symptom production. New Phytol. 2016, 209, 1106–1119. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, T.; Shen, D.; Wang, J.; Ling, X.; Hu, Z.; Chen, T.; Hu, J.; Huang, J.; Yu, W.; et al. Tomato yellow leaf curl virus intergenic siRNAs target a host long noncoding RNA to modulate disease symptoms. PLoS Pathog. 2019, 15, e1007534. [Google Scholar] [CrossRef]
- Shimura, H.; Pantaleo, V.; Ishihara, T.; Myojo, N.; Inaba, J.-i.; Sueda, K.; Burgyán, J.; Masuta, C. A viral satellite RNA induces yellow symptoms on tobacco by targeting a gene involved in chlorophyll biosynthesis using the RNA silencing machinery. PLoS Pathog. 2011, 7, e1002021. [Google Scholar] [CrossRef] [Green Version]
- Smith, N.A.; Eamens, A.L.; Wang, M.-B. Viral small interfering RNAs target host genes to mediate disease symptoms in plants. PLoS Pathog. 2011, 7, e1002022. [Google Scholar] [CrossRef] [Green Version]
- Bao, S.; Owens, R.A.; Sun, Q.; Song, H.; Liu, Y.; Eamens, A.L.; Feng, H.; Tian, H.; Wang, M.-B.; Zhang, R. Silencing of transcription factor encoding gene StTCP23 by small RNAs derived from the virulence modulating region of potato spindle tuber viroid is associated with symptom development in potato. PLoS Pathog. 2019, 15, e1008110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gellért, Á.; Pósa, T.; Fábián, A.; Szabó, L.; Bóka, K.; Forró, B.; Salánki, K.; Drahos, L.; Tóth, E.; Juhász, A.; et al. A single point mutation on the cucumber mosaic virus surface induces an unexpected and strong interaction with the F1 complex of the ATP synthase in Nicotiana clevelandii plants. Virus Res. 2018, 251, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zeng, R.; Chen, Z.; Liu, X.; Cao, Z.; Xie, Q.; Yang, C.; Lai, J. S-acylation of a geminivirus C4 protein is essential for regulating the CLAVATA pathway in symptom determination. J. Exp. Bot. 2018, 69, 4459–4468. [Google Scholar] [CrossRef] [Green Version]
- Salánki, K.; Kiss, L.; Gellért, Á.; Balázs, E. Identification a coat protein region of cucumber mosaic virus (CMV) essential for long-distance movement in cucumber. Arch. Virol. 2011, 156, 2279–2283. [Google Scholar] [CrossRef]
- Hasiów-Jaroszewska, B.; Borodynko, N.; Jackowiak, P.; Figlerowicz, M.; Pospieszny, H. Single mutation converts mild pathotype of the pepino mosaic virus into necrotic one. Virus Res. 2011, 159, 57–61. [Google Scholar] [CrossRef]
- Osterbaan, L.J.; Choi, J.; Kenney, J.; Flasco, M.; Vigne, E.; Schmitt-Keichinger, C.; Rebelo, A.R.; Cilia, M.; Fuchs, M. The identity of a single residue of the RNA-dependent RNA polymerase of grapevine fanleaf virus modulates vein clearing symptoms in Nicotiana benthamiana. Mol. Plant-Microbe Interact. 2019, 32, 790–801. [Google Scholar] [CrossRef]
- Zhan, B.; Zhao, W.; Li, S.; Yang, X.; Zhou, X. Functional scanning of apple geminivirus proteins as symptom determinants and suppressors of posttranscriptional gene silencing. Viruses 2018, 10, 488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandowski, D.J.; Dawson, W.O. A single amino acid change in tobacco mosaic virus replicase prevents symptom production. Mol. Plant-Microbe Interact. 1993, 6, 157–160. [Google Scholar] [CrossRef]
- Janzac, B.; Montarry, J.; Palloix, A.; Navaud, O.; Moury, B. A point mutation in the polymerase of potato virus Y confers virulence toward the Pvr4 resistance of pepper and a high competitiveness cost in susceptible cultivar. Mol. Plant-Microbe Interact. 2010, 23, 823–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, K.; Shiraishi, T.; Yoshida, T.; Fujita, N.; Netsu, O.; Yamaji, Y.; Namba, S. A replicase of potato virus X acts as the resistance-breaking determinant for JAX1-mediated resistance. Mol. Plant-Microbe Interact. 2013, 26, 1106–1112. [Google Scholar] [CrossRef]
- Knorr, D.A.; Dawson, W.O. A point mutation in the tobacco mosaic virus capsid protein gene induces hypersensitivity in Nicotiana Sylvestris. Proc. Natl. Acad. Sci. USA 1988, 85, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, A.L.; Grantham, G.L. A spontaneous mutation in the movement protein gene of brome mosaic virus modulates symptom phenotype in Nicotiana benthamiana. J. Virol. 1995, 69, 2689. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.; Dreher, T. Increased viral yield and symptom severity result from a single amino acid substitution in the turnip yellow mosaic virus movement protein. Mol. Plant-Microbe Interact. 1993, 6, 268–273. [Google Scholar] [CrossRef]
- Fujita, N.; Komatsu, K.; Ayukawa, Y.; Matsuo, Y.; Hashimoto, M.; Netsu, O.; Teraoka, T.; Yamaji, Y.; Namba, S.; Arie, T. N-terminal region of cysteine-rich protein (CRP) in carlaviruses is involved in the determination of symptom types. Mol. Plant Pathol. 2018, 19, 180–190. [Google Scholar] [CrossRef] [Green Version]
- Nam, M.; Koh, S.; Kim, S.U.; Domier, L.L.; Jeon, J.H.; Kim, H.G.; Lee, S.-H.; Bent, A.F.; Moon, J.S. Arabidopsis TTR1 causes LRR-dependent lethal systemic necrosis, rather than systemic acquired resistance, to Tobacco ringspot virus. Mol. Cells 2011, 32, 421–429. [Google Scholar] [CrossRef]
- Gnanasekaran, P.; Ponnusamy, K.; Chakraborty, S. A geminivirus betasatellite encoded βC1 protein interacts with PsbP and subverts PsbP-mediated antiviral defence in plants. Mol. Plant Pathol. 2019, 20, 943–960. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Wang, Z.Q.; Xiao, R.; Cao, L.; Wang, Y.; Xie, Y.; Zhou, X. Mimic phosphorylation of a βC1 protein encoded by TYLCCNB impairs its functions as a viral suppressor of RNA silencing and a symptom determinant. J. Virol. 2017, 91, e00300–e00317. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, M. Grapevine viruses: A multitude of diverse species with simple but overall poorly adopted management solutions in the vineyard. J. Plant Pathol. 2020, 102, 643–653. [Google Scholar] [CrossRef]
- Martelli, G.P. An overview on grapevine viruses, viroids, and the diseases they cause. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 31–46. [Google Scholar] [CrossRef]
- Vuittenez, A. Variation des symptômes de la dégénérescence infectieuse de la vigne. Interprétation d’expériences de transmission de la maladie par greffage. Comptes Rendus Acad. Sci. 1956, 243, 515–517. [Google Scholar]
- Schmitt-Keichinger, C.; Hemmer, C.; Berthold, F.; Ritzenthaler, C. Molecular, cellular and structural biology of grapevine fanleaf virus. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 83–107. [Google Scholar] [CrossRef] [Green Version]
- Digiaro, M.; Elbeaino, T.; Martelli, G.P. Grapevine fanleaf virus and other old world nepoviruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 47–82. [Google Scholar] [CrossRef]
- Andret-Link, P.; Marmonier, A.; Belval, L.; Hleibieh, K.; Ritzenthaler, C.; Demangeat, G. Ectoparasitic nematode vectors of grapevine viruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 505–529. [Google Scholar] [CrossRef]
- Horváth, J.; Tóbiás, I.; Hunyadi, K. New natural herbaceous hosts of grapevine fanleaf nepovirus. Hortic. Sci. 1994, 26, 31–32. [Google Scholar]
- Izadpanah, K.; Zaki-Aghl, M.; Zhang, Y.P.; Daubert, S.D.; Rowhani, A. Bermuda grass as a potential reservoir host for Grapevine fanleaf virus. Plant Dis. 2003, 87, 1179–1182. [Google Scholar] [CrossRef] [PubMed]
- Vigne, E.; Gottula, J.; Schmitt-Keichinger, C.; Komar, V.; Ackerer, L.; Belval, L.; Rakotomalala, L.; Lemaire, O.; Ritzenthaler, C.; Fuchs, M. A strain-specific segment of the RNA-dependent RNA polymerase of grapevine fanleaf virus determines symptoms in Nicotiana species. J. Gen. Virol. 2013, 94, 2803–2813. [Google Scholar] [CrossRef]
- Martin, I.R.; Vigne, E.; Berthold, F.; Komar, V.; Lemaire, O.; Fuchs, M.; Schmitt-Keichinger, C. The 50 distal amino acids of the 2AHP homing protein of Grapevine fanleaf virus elicit a hypersensitive reaction on Nicotiana occidentalis. Mol. Plant Pathol. 2018, 19, 731–743. [Google Scholar] [CrossRef] [Green Version]
- Perrone, I.; Chitarra, W.; Boccacci, P.; Gambino, G. Grapevine-virus-environment interactions: An intriguing puzzle to solve. New Phytol. 2017, 213, 983–987. [Google Scholar] [CrossRef] [Green Version]
- Legin, R.; Bass, P.; Etienne, L.; Fuchs, M. Selection of mild virus strains of fanleaf degeneration by comparative field performance of infected grapevines. Vitis 1993, 32, 103–110. [Google Scholar]
- Vigne, E.; Marmonier, A.; Fuchs, M. Multiple interspecies recombination events within RNA2 of Grapevine fanleaf virus and Arabis mosaic virus. Arch. Virol. 2008, 153, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Vigne, E.; Demangeat, G.; Komar, V.; Fuchs, M. Characterization of a naturally occurring recombinant isolate of grapevine fanleaf virus. Arch. Virol. 2005, 150, 2241–2255. [Google Scholar] [CrossRef] [PubMed]
- Walter, B.; Bass, P.; Legin, R.; Martin, C.; Vernoy, R.; Collas, A.; Vesselle, G. The use of a green–grafting technique for the detection of virus–like diseases of the grapevine. J. Phytopathol. 1990, 128, 137–145. [Google Scholar] [CrossRef]
- Vigne, E.; Bergdoll, M.; Guyader, S.; Fuchs, M. Population structure and genetic variability within isolates of grapevine fanleaf virus from a naturally infected vineyard in France: Evidence for mixed infection and recombination. J. Gen. Virol. 2004, 85, 2435–2445. [Google Scholar] [CrossRef] [PubMed]
- Vigne, E.; Garcia, S.; Komar, V.; Lemaire, O.; Hily, J.-M. Comparison of serological and molecular methods with high-throughput sequencing for the detection and quantification of grapevine fanleaf virus in vineyard samples. Front. Microbiol. 2018, 9, 2726. [Google Scholar] [CrossRef] [Green Version]
- Hily, J.-M.; Demaneche, S.; Poulicard, N.; Tannieres, M.; Djennane, S.; Beuve, M.; Vigne, E.; Demangeat, G.; Komar, V.; Gertz, C.; et al. Metagenomic-based impact study of transgenic grapevine rootstock on its associated virome and soil bacteriome. Plant Biotechnol. J. 2018, 16, 208–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, M.; Schmitt-Keichinger, C.; Sanfaçon, H. A renaissance in nepovirus research provides new insights into their molecular interface with hosts and vectors. In Advances in Virus Research; Academic Press: Cambridge, MA, USA, 2017; Volume 97, pp. 61–105. [Google Scholar]
- Canaguier, A.; Grimplet, J.; Di Gaspero, G.; Scalabrin, S.; Duchêne, E.; Choisne, N.; Mohellibi, N.; Guichard, C.; Rombauts, S.; Le Clainche, I.; et al. A new version of the grapevine reference genome assembly (12X.v2) and of its annotation (VCost.v3). Genom. Data 2017, 14, 56–62. [Google Scholar] [CrossRef]
- Vitulo, N.; Forcato, C.; Carpinelli, E.C.; Telatin, A.; Campagna, D.; D’Angelo, M.; Zimbello, R.; Corso, M.; Vannozzi, A.; Bonghi, C.; et al. A deep survey of alternative splicing in grape reveals changes in the splicing machinery related to tissue, stress condition and genotype. BMC Plant Biol. 2014, 14, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Alexa, A.; Rahnenfuhrer, J. TopGO: Enrichment Analysis for Gene Ontology. R Package Version 2.32.0. 2016. Available online: https://bioconductor.org/packages/release/bioc/html/topGO.html (accessed on 14 October 2021).
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.L.; Ledet-Jensen, J.; Ørntoft, T. Normalization of real-time quantitative RT-PCR data: A model-based variance estimation approach to identify genes suited for normalization—Applied to bladder- and colon-cancer data-sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.; Bakker, O.; van den Hoff, M.J.B.; Karlen, Y.; Moorman, A.F.M. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Koutouan, C.; Clerc, V.L.; Baltenweck, R.; Claudel, P.; Halter, D.; Hugueney, P.; Hamama, L.; Suel, A.; Huet, S.; Merlet, M.B.; et al. Link between carrot leaf secondary metabolites and resistance to Alternaria dauci. Sci. Rep. 2018, 8, 13746. [Google Scholar] [CrossRef]
- Komar, V.; Vigne, E.; Demangeat, G.; Fuchs, M. Beneficial effect of selective virus elimination on the performance of Vitis vinifera cv. Chardonnay. Am. J. Enol. Vitic. 2007, 58, 202–210. [Google Scholar]
- Komar, V.; Vigne, E.; Demangeat, G.; Lemaire, O.; Fuchs, M. Comparative performance of virus-infected Vitis vinifera cv. Savagnin rose grafted onto three rootstocks. Am. J. Enol. Vitic. 2010, 61, 68–73. [Google Scholar]
- Hily, J.-M.; Beuve, M.; Vigne, E.; Demangeat, G.; Candresse, T.; Lemaire, O. A genome-wide diversity study of grapevine rupestris stem pitting-associated virus. Arch. Virol. 2018, 163, 3105–3111. [Google Scholar] [CrossRef] [PubMed]
- The French–Italian public consortium for grapevine genome characterization. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar] [CrossRef]
- Perazzolli, M.; Moretto, M.; Fontana, P.; Ferrarini, A.; Velasco, R.; Moser, C.; Delledonne, M.; Pertot, I. Downy mildew resistance induced by Trichoderma harzianum T39 in susceptible grapevines partially mimics transcriptional changes of resistant genotypes. BMC Genom. 2012, 13, 660. [Google Scholar] [CrossRef]
- Lilly, S.T.; Drummond, R.S.M.; Pearson, M.N.; MacDiarmid, R.M. Identification and validation of reference genes for normalization of transcripts from virus-infected Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2010, 24, 294–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Shi, L.; Han, C.; Yu, J.; Li, D.; Zhang, Y. Validation of reference genes for gene expression studies in virus-infected Nicotiana benthamiana using quantitative real-time PCR. PLoS ONE 2012, 7, e46451. [Google Scholar] [CrossRef]
- Zhang, K.; Niu, S.; Di, D.; Shi, L.; Liu, D.; Cao, X.; Miao, H.; Wang, X.; Han, C.; Yu, J.; et al. Selection of reference genes for gene expression studies in virus-infected monocots using quantitative real-time PCR. J. Biotechnol. 2013, 168, 7–14. [Google Scholar] [CrossRef]
- Großkinsky, D.K.; Edelsbrunner, K.; Pfeifhofer, H.; van der Graaff, E.; Roitsch, T. Cis- and trans-zeatin differentially modulate plant immunity. Plant Signal. Behav. 2013, 8, e24798. [Google Scholar] [CrossRef]
- Flamini, R.; De Rosso, M. Chapter 5—High-resolution mass spectrometry and biological properties of grapevine and wine stilbenoids. In Studies in Natural Products Chemistry; Atta ur, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 61, pp. 175–210. [Google Scholar]
- Calderón, A.A.; Zapata, J.M.; Muñoz, R.; Pedreño, M.A.; Barceló, A.R. Resveratrol production as a part of the hypersensitive like response of grapevine cells to an elicitor from Trichoderma viride. New Phytol. 1993, 124, 455–463. [Google Scholar] [CrossRef]
- Chang, X.; Heene, E.; Qiao, F.; Nick, P. The phytoalexin resveratrol regulates the initiation of hypersensitive cell death in vitis cell. PLoS ONE 2011, 6, e26405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stempien, E.; Goddard, M.-L.; Leva, Y.; Bénard-Gellon, M.; Laloue, H.; Farine, S.; Kieffer-Mazet, F.; Tarnus, C.; Bertsch, C.; Chong, J. Secreted proteins produced by fungi associated with Botryosphaeria dieback trigger distinct defense responses in Vitis vinifera and Vitis rupestris cells. Protoplasma 2018, 255, 613–628. [Google Scholar] [CrossRef]
- Künstler, A.; Bacsó, R.; Gullner, G.; Hafez, Y.M.; Király, L. Staying alive—is cell death dispensable for plant disease resistance during the hypersensitive response? Physiol. Mol. Plant Pathol. 2016, 93, 75–84. [Google Scholar] [CrossRef]
- Geng, C.; Wang, H.-Y.; Liu, J.; Yan, Z.-Y.; Tian, Y.-P.; Yuan, X.-F.; Gao, R.; Li, X.-D. Transcriptomic changes in Nicotiana benthamiana plants inoculated with the wild-type or an attenuated mutant of Tobacco vein banding mosaic virus. Mol. Plant Pathol. 2016, 18, 1175–1188. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Aix, C.; Pascual, L.; Cañizares, J.; Sánchez-Pina, M.A.; Aranda, M.A. Transcriptomic profiling of melon necrotic spot virus-infected melon plants revealed virus strain and plant cultivar-specific alterations. BMC Genom. 2016, 17, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Wang, Z.; Gu, Q.; Li, H.; Han, W.; Shi, Y. Transcriptome analysis of Cucumis sativus infected by Cucurbit chlorotic yellows virus. Virol. J. 2017, 14, 18. [Google Scholar] [CrossRef] [Green Version]
- Paudel, D.B.; Sanfaçon, H. Exploring the diversity of mechanisms associated with plant tolerance to virus infection. Front. Plant Sci. 2018, 9, 1575. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, X.; Hong, Y.; Liu, Y. Chloroplast in plant-virus interaction. Front. Microbiol. 2016, 7, 1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sgherri, C.; Ranieri, A.; Quartacci, M.F. Antioxidative responses in Vitis vinifera infected by grapevine fanleaf virus. J. Plant Physiol. 2013, 170, 121–128. [Google Scholar] [CrossRef]
- Abracheva, P. The chlorophyll content of vine leaves as influenced by the Court-noué. Hortic. Vitic. Sci. 1977, 14, 102–106. [Google Scholar]
- Li, Y.-Q.; Li, Z.-L.; Zhao, W.-J.; Wen, R.-X.; Meng, Q.-W.; Zeng, Y. Synthesis of stilbene derivatives with inhibition of SARS coronavirus replication. Eur. J. Med. Chem. 2006, 41, 1084–1089. [Google Scholar] [CrossRef]
- Song, P.; Yu, X.; Yang, W.; Wang, Q. Natural phytoalexin stilbene compound resveratrol and its derivatives as anti-tobacco mosaic virus and anti-phytopathogenic fungus agents. Sci. Rep. 2021, 11, 16509. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moffett, P.; Gad, L.; John, P.C. Mechanisms of recognition in dominant R gene mediated resistance. Adv. Virus Res. 2009, 75, 1–33. [Google Scholar]
- Soosaar, J.L.M.; Burch-Smith, T.M.; Dinesh-Kumar, S.P. Mechanisms of plant resistance to viruses. Nat. Rev. Microbiol. 2005, 3, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Morel, J.-B.; Dangl, J.L. The hypersensitive response and the induction of cell death in plants. Cell Death Differ. 1997, 4, 671–683. [Google Scholar] [CrossRef] [Green Version]
- Bendahmane, A.; Kanyuka, K.; Baulcombe, D.C. The Rx gene from potato controls separate virus resistance and cell death responses. Plant Cell 1999, 11, 781–791. [Google Scholar] [CrossRef] [Green Version]
- Lukan, T.; Baebler, Š.; Pompe-Novak, M.; Guček, K.; Zagorščak, M.; Coll, A.; Gruden, K. Cell death is not sufficient for the restriction of potato virus y spread in hypersensitive response-conferred resistance in potato. Front. Plant Sci. 2018, 9, 168. [Google Scholar] [CrossRef] [Green Version]
- Cole, A.B.; Király, L.; Ross, K.; Schoelz, J.E. Uncoupling resistance from cell death in the hypersensitive response of Nicotiana species to cauliflower mosaic virus Infection. Mol. Plant-Microbe Interact. 2001, 14, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, M.; Desvoyes, B.; Turina, M.; Noad, R.; Scholthof, H.B. Genetic dissection of tomato bushy stunt virus P19-protein-mediated host-dependent symptom induction and systemic invasion. Virology 2000, 266, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Sekine, K.-T.; Ishihara, T.; Hase, S.; Kusano, T.; Shah, J.; Takahashi, H. Single amino acid alterations in Arabidopsis thaliana RCY1 compromise resistance to cucumber mosaic virus, but differentially suppress hypersensitive response-like cell death. Plant Mol. Biol. 2006, 62, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, F.; Zhang, G.; Jia, S.; Yan, Z. FaWRKY11 transcription factor positively regulates resistance to Botrytis cinerea in strawberry fruit. Sci. Hortic. 2021, 279, 109893. [Google Scholar] [CrossRef]
- Arraño-Salinas, P.; Dominguez-Figueroa, J.; Herrera-Vasquez, A.; Zavala, D.; Medina, J.; Vicente-Carbajosa, J.; Meneses, C.; Canessa, P.; Moreno, A.A.; Blanco-Herrera, F. WRKY7, -11 and -17 transcription factors are modulators of the bZIP28 branch of the unfolded protein response during PAMP-triggered immunity in Arabidopsis thaliana. Plant Sci. 2018, 277, 242–250. [Google Scholar] [CrossRef]
- Kim, K.C.; Fan, B.; Chen, Z. Pathogen-induced Arabidopsis WRKY7 is a transcriptional repressor and enhances plant susceptibility to Pseudomonas syringae. Plant Physiol. 2006, 142, 1180–1192. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Le Henanff, G.; Bertsch, C.; Walter, B. Identification, expression analysis and characterization of defense and signaling genes in Vitis vinifera. Plant Physiol. Biochem. 2008, 46, 469–481. [Google Scholar] [CrossRef]
- Reim, S.; Rohr, A.-D.; Winkelmann, T.; Weiß, S.; Liu, B.; Beerhues, L.; Schmitz, M.; Hanke, M.-V.; Flachowsky, H. Genes involved in stress response and especially in phytoalexin biosynthesis are upregulated in four Malus genotypes in response to apple replant disease. Front. Plant Sci. 2020, 10, 1724. [Google Scholar] [CrossRef]
- Mishra, A.K.; Kumar, A.; Mishra, D.; Nath, V.S.; Jakše, J.; Kocábek, T.; Killi, U.K.; Morina, F.; Matoušek, J. Genome-wide transcriptomic analysis reveals insights into the response to citrus bark cracking viroid (CBCVd) in hop (Humulus lupulus L.). Viruses 2018, 10, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, P.; Ding, Y. Stories of salicylic acid: A plant defense hormone. Trends Plant. Sci. 2020, 25, 549–565. [Google Scholar] [CrossRef]
- Van Butselaar, T.; Van den Ackerveken, G. Salicylic acid steers the growth–immunity tradeoff. Trends Plant Sci. 2020, 25, 566–576. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment a | Plant b | Total Reads c | Mapped Reads d | % e | Unique Reads f | % e | Reads Assigned to a Gene g | % e |
|---|---|---|---|---|---|---|---|---|
| Control | B31 | 51,964,098 | 42,741,560 | 82 | 42,153,543 | 81 | 40,248,531 | 77 |
| B34 | 59,910,092 | 52,451,408 | 88 | 51,647,516 | 86 | 49,130,979 | 82 | |
| C51 | 65,724,896 | 56,105,085 | 85 | 55,354,647 | 84 | 52,784,724 | 80 | |
| F13 | B52 | 69,714,014 | 59,901,468 | 86 | 58,981,711 | 85 | 56,246,251 | 81 |
| B53 | 75,771,722 | 66,064,642 | 87 | 65,065,605 | 86 | 62,069,793 | 82 | |
| C38 | 85,633,030 | 74,306,333 | 87 | 73,166,998 | 85 | 70,037,177 | 82 | |
| B844 | B47 | 165,443,500 | 139,722,683 | 84 | 137,021,434 | 83 | 129,811,616 | 78 |
| C39 | 54,436,852 | 46,539,167 | 85 | 45,791,912 | 84 | 43,348,284 | 80 | |
| C40 | 58,344,878 | 49,214,165 | 84 | 48,446,050 | 83 | 46,189,830 | 79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin, I.R.; Vigne, E.; Velt, A.; Hily, J.-M.; Garcia, S.; Baltenweck, R.; Komar, V.; Rustenholz, C.; Hugueney, P.; Lemaire, O.; et al. Severe Stunting Symptoms upon Nepovirus Infection Are Reminiscent of a Chronic Hypersensitive-like Response in a Perennial Woody Fruit Crop. Viruses 2021, 13, 2138. https://doi.org/10.3390/v13112138
Martin IR, Vigne E, Velt A, Hily J-M, Garcia S, Baltenweck R, Komar V, Rustenholz C, Hugueney P, Lemaire O, et al. Severe Stunting Symptoms upon Nepovirus Infection Are Reminiscent of a Chronic Hypersensitive-like Response in a Perennial Woody Fruit Crop. Viruses. 2021; 13(11):2138. https://doi.org/10.3390/v13112138
Chicago/Turabian StyleMartin, Isabelle R., Emmanuelle Vigne, Amandine Velt, Jean-Michel Hily, Shahinez Garcia, Raymonde Baltenweck, Véronique Komar, Camille Rustenholz, Philippe Hugueney, Olivier Lemaire, and et al. 2021. "Severe Stunting Symptoms upon Nepovirus Infection Are Reminiscent of a Chronic Hypersensitive-like Response in a Perennial Woody Fruit Crop" Viruses 13, no. 11: 2138. https://doi.org/10.3390/v13112138
APA StyleMartin, I. R., Vigne, E., Velt, A., Hily, J. -M., Garcia, S., Baltenweck, R., Komar, V., Rustenholz, C., Hugueney, P., Lemaire, O., & Schmitt-Keichinger, C. (2021). Severe Stunting Symptoms upon Nepovirus Infection Are Reminiscent of a Chronic Hypersensitive-like Response in a Perennial Woody Fruit Crop. Viruses, 13(11), 2138. https://doi.org/10.3390/v13112138