
Porcine Endogenous Retroviruses and Xenotransplantation, 2021


Abstract
:1. Introduction
2. Xenotransplantation: The Need and the Achievements

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pig Transplant | Donor Pigs’ Genetic Background | Longest Survival Time (Days) | Reference |
|---|---|---|---|
| Islet cells | Wild-type minipigs | 950 | Shin et al. [17] |
| Heart, heterotopic | GTKO/CD46/TBM | 945 | Mohiuddin et al. [18] |
| Heart, orthotopic, life-supporting | GTKO/CD46/TBM | 195 | Längin et al. [19] |
| Kidney, life-supporting | GTKO/CD55 | 499 | Kim et al. [20] |
| Neurons | CTLA4-Ig | 549 | Aron Badin et al. [21] |
| Cornea | GTKO | 375 | Yoon et al. [22] |
| Liver | GTKO | 29 | Shah et al. [23] |
| Lung | GTKO/CD47/CD55 | 14 | Watanabe et al. [24] |
3. Exogenous and Endogenous Retroviruses
4. PERVs: Biology

5. Host Range In Vitro and In Vivo and Receptors
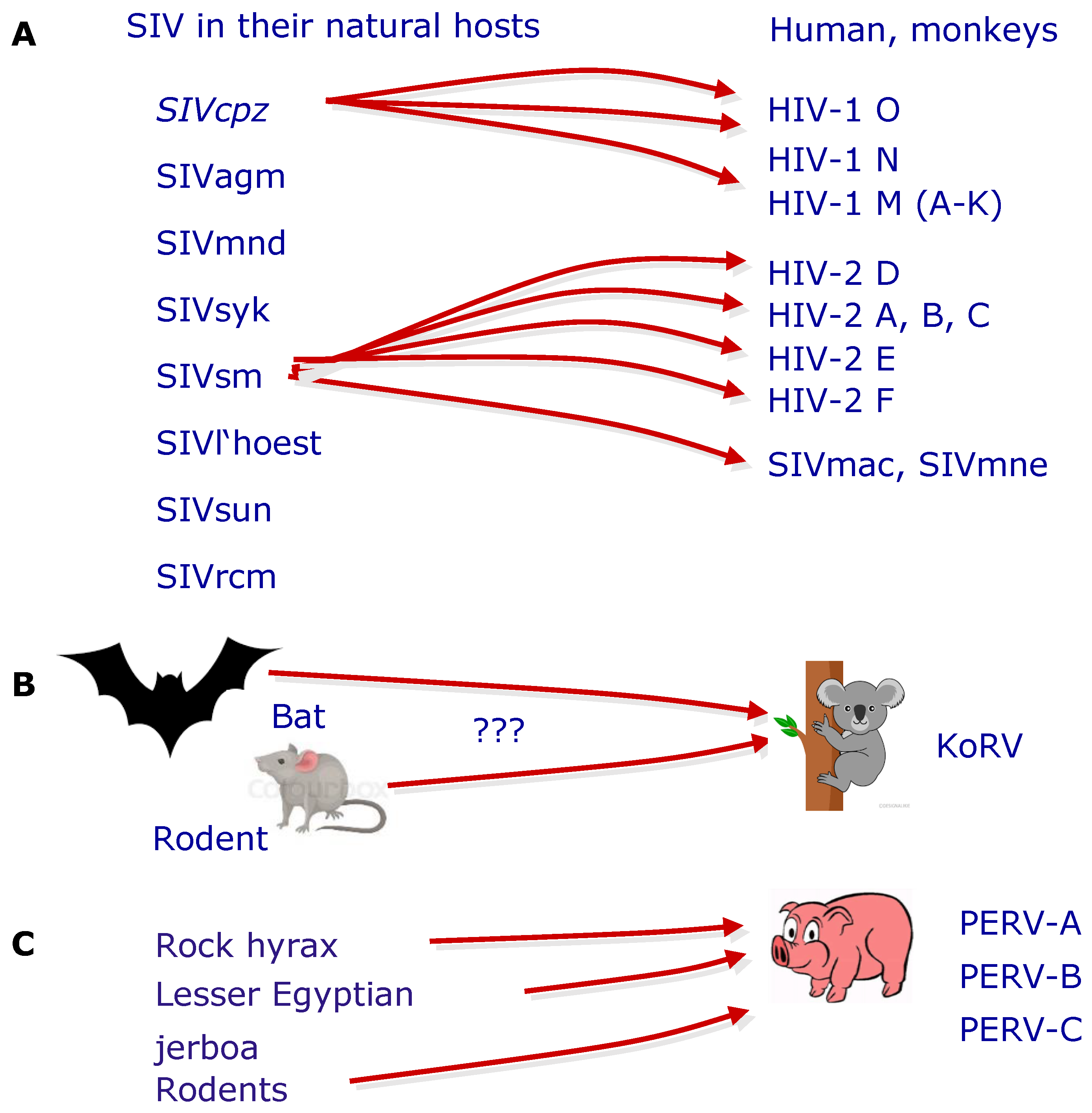
6. The Origin of PERV
7. Detection Systems
8. Copy Number
9. PERVs and Restriction Factors
10. Recombinant PERVs and Minipigs
11. PERVs and Stem Cells
12. PERVs and Pig Tumors
13. Absence of PERV Transmission in Preclinical and Clinical Trials
14. Strategies to Prevent PERV Infection
14.1. Selection of Suitable Pigs
14.2. Antiretroviral Drugs
14.3. Vaccination
14.4. RNA Interference
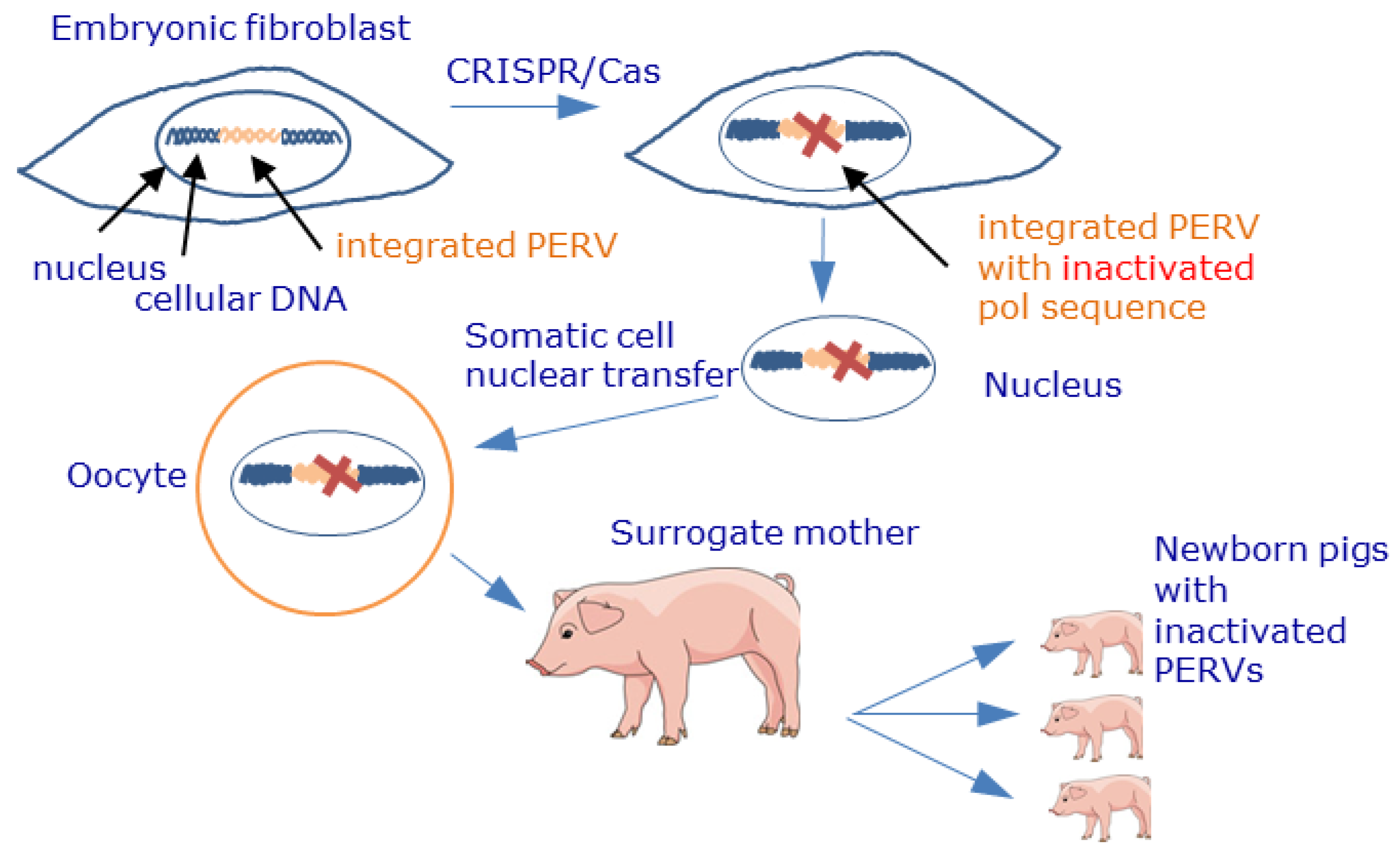
14.5. Genome Editing
- Furthermore, in all preclinical trials in nonhuman primates, no transmission of PERVs was observed [149,150,151,152]. However, nonhuman primates are not an ideal animal model to assess the risk of PERV transmission in xenotransplantation [153]. This is based on the fact that the main receptor for PERV-A and PERV-A/C entry, PERV-A receptor 1 (PAR-1), was found to be genetically deficient in baboons and cynomolgus monkeys (see above) [43]. In infection experiments in small animals and nonhuman primates with or without pharmaceutical immunosuppression, PERV transmission also was not observed: The mouse receptor was mutated and not effective, and the rat receptor was expressed only at low concentrations on the cell surface [44], showing that mouse and rat cells could not be infected [45,46,154].
- The main obstacle is certainly the risk of inbreeding of CRISPR/Cas-inactivated pigs when generating high numbers of donor pigs for xenotransplantation.
15. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Cooper, D.K.C.; Hara, H.; Iwase, H.; Yamamoto, T.; Wang, Z.; Jagdale, A.; Bikhet, M.H.; Nguyen, H.Q.; Foote, J.B.; Paris, W.D.; et al. Pig kidney xenotransplantation: Progress toward clinical trials. Clin. Transplant. 2021, 35, e14139. [Google Scholar] [CrossRef] [PubMed]
- Coe, T.M.; Markmann, J.F.; Rickert, C.G. Current status of porcine islet xenotransplantation. Curr. Opin. Organ Transplant. 2020, 25, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Denner, J.; Tönjes, R.R. Infection barriers to successful xenotransplantation focusing on porcine endogenous retroviruses. Clin. Microbiol. Rev. 2012, 25, 318–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scobie, L.; Takeuchi, Y. Porcine endogenous retrovirus and other viruses in xenotransplantation. Curr. Opin. Organ Transplant. 2009, 14, 175–179. [Google Scholar] [CrossRef]
- Wilson, C.A. Porcine endogenous retroviruses and xenotransplantation. Cell. Mol. Life Sci. 2008, 65, 3399–3412. [Google Scholar] [CrossRef]
- Magre, S.; Takeuchi, Y.; Bartosch, B. Xenotransplantation and pig endogenous retroviruses. Rev. Med. Virol. 2003, 13, 311–329. [Google Scholar] [CrossRef]
- Blusch, J.H.; Patience, C.; Martin, U. Pig endogenous retroviruses and xenotransplantation. Xenotransplantation 2002, 9, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Kwong, A.J.; Kim, W.R.; Lake, J.R.; Smith, J.M.; Schladt, D.P.; Skeans, M.A.; Noreen, S.M.; Foutz, J.; Booker, S.E.; Cafarella, M.; et al. OPTN/SRTR 2019 Annual Data Report: Liver. Am. J. Transplant. 2021, 21 (Suppl. 2), 208–315. [Google Scholar] [CrossRef]
- Organ Donor Statistics. Available online: https://www.organdonor.gov/statistics-stories/statistics.html (accessed on 9 September 2021).
- Eurotransplant Statistics. Available online: https://www.eurotransplant.org/statistics/ (accessed on 9 September 2021).
- Mrzljak, A.; Novak, R.; Pandak, N.; Tabain, I.; Franusic, L.; Barbic, L.; Bogdanic, M.; Savic, V.; Mikulic, D.; Pavicic-Saric, J.; et al. Emerging and neglected zoonoses in transplant population. World J. Transplant. 2020, 10, 47–63. [Google Scholar] [CrossRef]
- Cooper, D.K.; Matsumoto, S.; Abalovich, A.; Itoh, T.; Mourad, N.I.; Gianello, P.; Wolf, E.; Cozzi, E. Progress in Clinical Encapsulated Islet Xenotransplantation. Transplantation 2016, 100, 2301–2308. [Google Scholar] [CrossRef]
- Matsumoto, S.; Tan, P.; Baker, J.; Durbin, K.; Tomiya, M.; Azuma, K.; Doi, M.; Elliott, R. Clinical Porcine Islet Xenotransplantation Under Comprehensive Regulation. Transplant. Proc. 2014, 46, 1992–1995. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.C.; Liu, X.; Jin, Y.; Jiang, D.L.; Wie, X.S.; Xie, H.T. Lamellar keratoplasty treatment of fungal corneal ulcers with acellular porcine corneal stroma. Am. J. Transplant. 2015, 15, 1068–1075. [Google Scholar] [CrossRef]
- Brink, J.G.; Hassoulas, J. The first human heart transplant and further advances in cardiac transplantation at Groote Schuur Hospital and the University of Cape Town. Cardiovasc. J. Afr. 2009, 20, 31–35. [Google Scholar]
- Ärzteblatt. Available online: https://www.aerzteblatt.de/nachrichten/100967/Erste-Herztransplantation-in-Deutschland-liegt-50-Jahrezurueck (accessed on 3 September 2021).
- Shin, J.-S.; Min, B.-H.; Kim, J.-M.; Kim, J.-S.; Yoon, I.H.; Kim, H.J.; Kim, Y.-H.; Jang, J.Y.; Kang, H.J.; Lim, D.-G.; et al. Failure of transplantation tolerance induction by autologous regulatory T cells in the pig-to-non-human primate islet xenotransplantation model. Xenotransplantation 2016, 23, 300–309. [Google Scholar] [CrossRef]
- Mohiuddin, M.M.; Singh, A.K.; Corcoran, P.C.; Iii, M.L.T.; Clark, T.; Lewis, B.G.; Hoyt, R.F.; Eckhaus, M.; Iii, R.N.P.; Belli, A.; et al. Chimeric 2C10R4 anti-CD40 antibody therapy is critical for long-term survival of GTKO.hCD46.hTBM pig-to-primate cardiac xenograft. Nat. Commun. 2016, 7, 11138. [Google Scholar] [CrossRef] [PubMed]
- Längin, M.; Mayr, T.; Reichart, B.; Michel, S.; Buchholz, S.; Guethoff, S.; Dashkevich, A.; Baehr, A.; Egerer, S.; Bauer, A.; et al. Consistent success in life-supporting porcine cardiac xenotransplantation. Nature 2018, 564, 430–433. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Mathews, D.V.; Breeden, C.P.; Higginbotham, L.B.; Ladowski, J.; Martens, G.; Stephenson, A.; Farris, A.B.; Strobert, E.A.; Jenkins, J.; et al. Long-term survival of pig-to-rhesus macaque renal xenografts is dependent on CD4 T cell depletion. Am. J. Transplant. 2019, 19, 2174–2185. [Google Scholar] [CrossRef] [PubMed]
- Aron Badin, R.; Vadori, M.; Vanhove, B.; Nerriere-Daguin, V.; Naveilhan, P.; Neveu, I.; Jan, C.; Lévèque, X.; Venturi, E.; Mermillod, P.; et al. Cell Therapy for Parkinson’s Disease: A Translational Approach to Assess the Role of Local and Systemic Immunosuppression. Am. J. Transplant. 2016, 16, 2016–2029. [Google Scholar] [CrossRef] [Green Version]
- Yoon, C.H.; Choi, S.H.; Choi, H.J.; Lee, H.J.; Kang, H.J.; Kim, J.M.; Park, C.G.; Choi, K.; Kim, H.; Ahn, C.; et al. Long-term survival of full-thickness corneal xenografts from α1,3-galactosyltransferase gene-knockout miniature pigs in non-human primates. Xenotransplantation 2020, 27, e12559. [Google Scholar] [CrossRef]
- Shah, J.A.; Patel, M.S.; Elias, N.; Navarro-Alvarez, N.; Rosales, I.; Wilkinson, R.A.; Louras, N.J.; Hertl, M.; Fishman, J.A.; Colvin, R.B.; et al. Prolonged Survival Following Pig-to-Primate Liver Xenotransplantation Utilizing Exogenous Coagulation Factors and Costimulation Blockade. Am. J. Transplant. 2017, 17, 2178–2185. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, H.; Ariyoshi, Y.; Pomposelli, T.; Takeuchi, K.; Ekanayake-Alper, D.K.; Boyd, L.K.; Arn, S.J.; Sahara, H.; Shimizu, A.; Ayares, D.; et al. Intra-bone bone marrow transplantation from hCD47 transgenic pigs to baboons prolongs chimerism to >60 days and promotes increased porcine lung transplant survival. Xenotransplantation 2020, 27, e12552. [Google Scholar] [CrossRef] [PubMed]
- Coffin, L.M.; Hughes, S.H.; Varmus, H.E. (Eds.) Retroviruses; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Lower, R.; Lower, J.; Kurth, R. The viruses in all of us: Characteristics and biological significance of human endogenous retrovirus sequences. Proc. Natl. Acad. Sci. USA 1996, 93, 5177–5184. [Google Scholar] [CrossRef] [Green Version]
- Lavialle, C.; Cornelis, G.; Dupressoir, A.; Esnault, C.; Heidmann, O.; Vernochet, C.; Heidmann, T. Paleovirology of ‘syncytins’, retroviral env genes exapted for a role in placentation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120507. [Google Scholar] [CrossRef] [Green Version]
- Denner, J. Expression and function of endogenous retroviruses in the placenta. APMIS 2016, 124, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Denner, J.; Young, P.R. Koala retroviruses: Characterization and impact on the life of koalas. Retrovirology 2013, 10, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Tissier, P.; Stoye, J.P.; Takeuchi, Y.; Patience, C.; Weiss, R.A. Two sets of human-tropic pig retrovirus. Nature 1997, 389, 681–682. [Google Scholar] [CrossRef]
- Akiyoshi, D.E.; Denaro, M.; Zhu, H.; Greenstein, J.L.; Banerjee, P.; Fishman, J.A. Identification of a full-length cDNA for an endogenous retrovirus of miniature swine. J. Virol. 1998, 72, 4503–4507. [Google Scholar] [CrossRef] [Green Version]
- Denner, J. The transmembrane proteins contribute to immunodeficiencies induced by HIV-1 and other retroviruses. AIDS 2014, 28, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Haraguchi, S.; Good, R.A.; Day, N.K. Immunosuppressive retroviral peptides: cAMP and cytokine patterns. Immunol. Today 1995, 16, 595–603. [Google Scholar] [CrossRef]
- Tacke, S.J.; Kurth, R.; Denner, J. Porcine endogenous retroviruses inhibit human immune cell function: Risk for xenotransplanta-tion? Virology 2000, 268, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Denner, J.; Specke, V.; Thiesen, U.; Karlas, A.; Kurth, R. Genetic alterations of the long terminal repeat of an ecotropic porcine endogenous retrovirus during passage in human cells. Virology 2003, 314, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Denner, J. Why was PERV not transmitted during preclinical and clinical xenotransplantation trials and after inoculation of animals? Retrovirology 2018, 15, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Choi, J.Y.; Lee, H.-J.; Kim, K.-C.; Choi, B.-S.; Oh, Y.-K.; Kim, Y.B. Repression of porcine endogenous retrovirus infection by human APOBEC3 proteins. Biochem. Biophys. Res. Commun. 2011, 407, 266–270. [Google Scholar] [CrossRef]
- Martin, U.; Winkler, M.E.; Id, M.; Radeke, H.; Arseniev, L.; Takeuchi, Y.; Simon, A.R.; Patience, C.; Haverich, A.; Steinhoff, G. Productive infection of primary human endothelial cells by pig endogenous retrovirus (PERV). Xenotransplantation 2000, 7, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. Porcine endogenous retrovirus infection of human peripheral blood mononuclear cells. Xenotransplantation 2015, 22, 151–152. [Google Scholar] [CrossRef]
- Argaw, T.; Colon-Moran, W.; Wilson, C.A. Limited infection without evidence of replication by porcine endogenous retrovirus in guinea pigs. J. Gen. Virol. 2004, 85 Pt 1, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Ericsson, T.A.; Takeuchi, Y.; Templin, C.; Quinn, G.; Farhadian, S.F.; Wood, J.C.; Oldmixon, B.A.; Suling, K.M.; Ishii, J.K.; Kitagawa, Y.; et al. Identification of receptors for pig endogenous retrovirus. Proc. Natl. Acad. Sci. USA 2003, 100, 6759–6764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colon-Moran, W.; Argaw, T.; Wilson, C.A. Three cysteine residues of SLC52A1, a receptor for the porcine endogenous retrovirus-A (PERV-A), play a critical role in cell surface expression and infectivity. Virology 2017, 507, 140–150. [Google Scholar] [CrossRef]
- Mattiuzzo, G.; Takeuchi, Y. Suboptimal Porcine Endogenous Retrovirus Infection in Non-Human Primate Cells: Implication for Preclinical Xenotransplantation. PLoS ONE 2010, 5, e13203. [Google Scholar] [CrossRef]
- Mattiuzzo, G.; Matouskova, M.; Takeuchi, Y. Differential resistance to cell entry by porcine endogenous retrovirus subgroup A in rodent species. Retrovirology 2007, 4, 93. [Google Scholar] [CrossRef] [Green Version]
- Irgang, M.; Karlas, A.; Laue, C.; Specke, V.; Tacke, S.J.; Kurth, R.; Schrezenmeir, J.; Denner, J. Porcine endogenous retroviruses PERV-A and PERV-B infect neither mouse cells in vitro nor SCID mice In Vivo. Intervirology 2005, 48, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Martina, Y.; Marcucci, K.T.; Cherqui, S.; Szabo, A.; Drysdale, T.; Srinivisan, U.; Wilson, C.A.; Patience, C.; Salomon, D.R. Mice transgenic for a human porcine endogenous retrovirus receptor are susceptible to productive viral infection. J. Virol. 2006, 80, 3135–3146. [Google Scholar] [CrossRef] [Green Version]
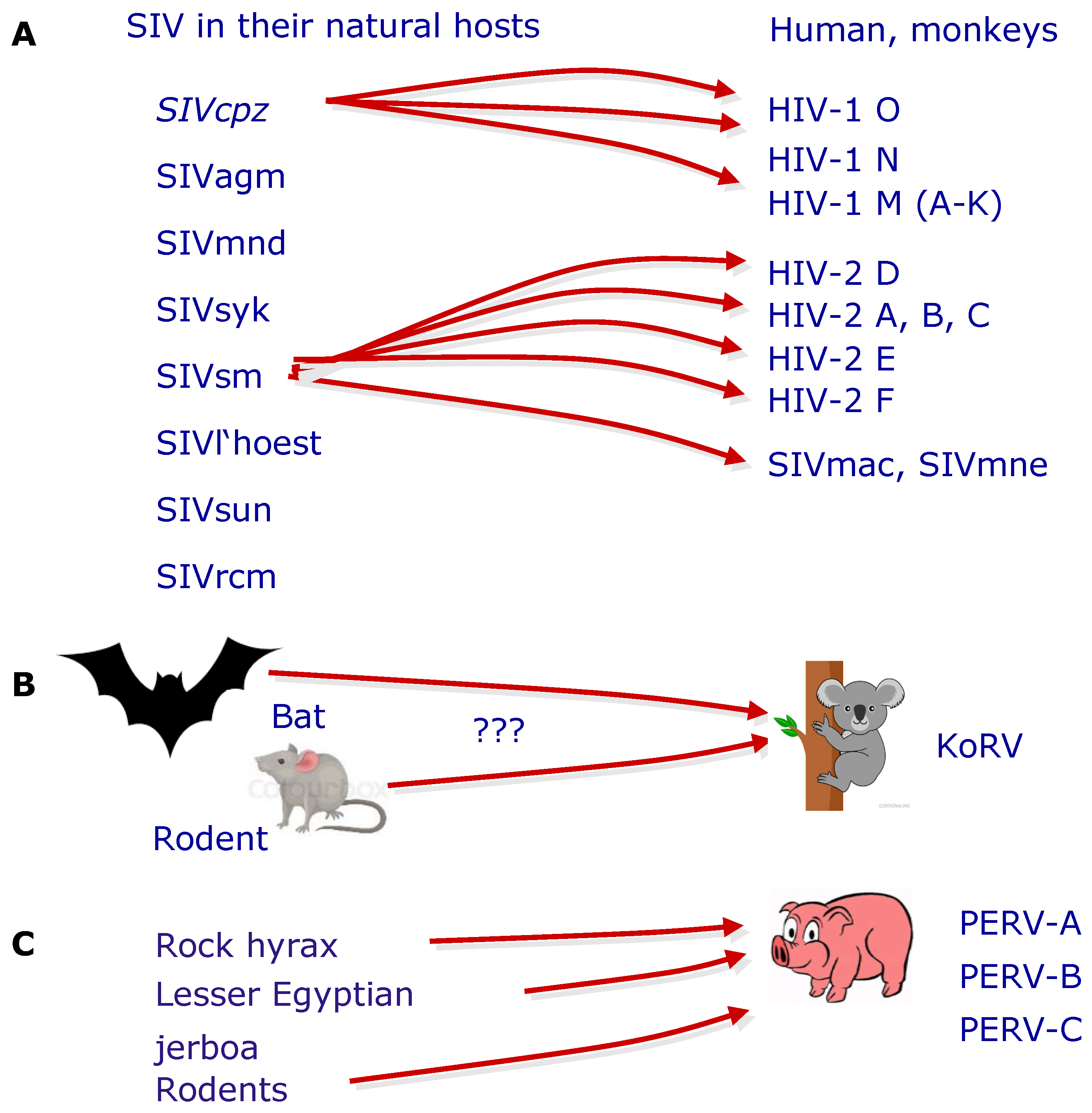
- Denner, J. The origin of porcine endogenous retroviruses (PERVs). Arch. Virol. 2021, 166, 1007–1013. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, X.; Duan, X.; Cui, J. Ancient origin and complex evolution of porcine endogenous retroviruses. Biosaf. Health 2020, 2, 142–151. [Google Scholar] [CrossRef]
- D’Arc, M.; Ayouba, A.; Esteban, A.; Learn, G.H.; Boué, V.; Liegeois, F.; Etienne, L.; Tagg, N.; Leendertz, F.H.; Boesch, C.; et al. Origin of the HIV-1 group O epidemic in western lowland gorillas. Proc. Natl. Acad. Sci. USA 2015, 112, E1343–E1352. [Google Scholar] [CrossRef] [Green Version]
- Etienne, L.; Hahn, B.; Sharp, P.M.; Matsen, F.A.; Emerman, M. Gene Loss and Adaptation to Hominids Underlie the Ancient Origin of HIV-1. Cell Host Microbe 2013, 14, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, P.; Hahn, B.H. Origins of HIV and the AIDS Pandemic. Cold Spring Harb. Perspect. Med. 2011, 1, a006841. [Google Scholar] [CrossRef] [Green Version]
- Denner, J. Transspecies Transmission of Gammaretroviruses and the Origin of the Gibbon Ape Leukaemia Virus (GaLV) and the Koala Retrovirus (KoRV). Viruses 2016, 8, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denner, J. Transspecies transmissions of retroviruses: New cases. Virology 2007, 369, 229–233. [Google Scholar] [CrossRef] [Green Version]
- Denner, J. Sensitive detection systems for infectious agents in xenotransplantation. Xenotransplantation 2020, e12594. [Google Scholar] [CrossRef] [Green Version]
- Godehardt, A.W.; Costa, M.R.; Tönjes, R.R. Review on porcine endogenous retrovirus detection assays—Impact on quality and safety of xenotransplants. Xenotransplantation 2015, 22, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Gola, J.; Mazurek, U. Detection of porcine endogenous retrovirus in xenotransplantation. Reprod. Biol. 2014, 14, 68–73. [Google Scholar] [CrossRef]
- Scobie, L.; Taylor, S.; Wood, J.C.; Suling, K.M.; Quinn, G.; Meikle, S.; Patience, C.; Schuurman, H.J.; Onions, D.E. Absence of replication-competent human-tropic porcine endogenous retroviruses in the germ line DNA of inbred miniature Swine. J. Virol. 2004, 78, 2502–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denner, J.; Scobie, L. Are there better assays to evaluate the risk of transmission of porcine endogenous retroviruses (PERVs) to human cells? Xenotransplantation 2019, 26, e12510. [Google Scholar] [CrossRef] [PubMed]
- Krüger, L.; Kristiansen, Y.; Reuber, E.; Möller, L.; Laue, M.; Reimer, C.; Denner, J. A Comprehensive Strategy for Screening for Xenotransplantation-Relevant Viruses in a Second Isolated Population of Gottingen Minipigs. Viruses 2019, 12, 38. [Google Scholar] [CrossRef] [Green Version]
- Fishman, J.A. Prevention of infection in xenotransplantation: Designated pathogen-free swine in the safety equation. Xenotransplantation 2020, 27, e12595. [Google Scholar] [CrossRef]
- Fishman, J.A. Infectious disease risks in xenotransplantation. Am. J. Transplant. 2018, 18, 1857–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fishman, J.A. Infection in xenotransplantation: Opportunities and challenges. Curr. Opin. Organ Transplant. 2019, 24, 527–534. [Google Scholar] [CrossRef]
- Gazda, L.S.; Collins, J.; Lovatt, A.; Holdcraft, R.W.; Morin, M.J.; Galbraith, D.; Graham, M.; Laramore, M.A.; MacLean, C.; Black, J.; et al. A comprehensive microbiological safety approach for agarose encapsulated porcine islets intended for clinical trials. Xenotransplantation 2016, 23, 444–463. [Google Scholar] [CrossRef] [PubMed]
- Hartline, C.B.; Conner, R.L.; James, S.H.; Potter, J.; Gray, E.; Estrada, J.; Tector, M.; Tector, A.J.; Prichard, M.N. Xenotransplantation panel for the detection of infectious agents in pigs. Xenotransplantation 2018, 25, e12427. [Google Scholar] [CrossRef] [Green Version]
- Kono, K.; Kataoka, K.; Yuan, Y.; Yusa, K.; Uchida, K.; Sato, Y. A highly sensitive method for the detection of recombinant PERV-A/C env RNA using next generation sequencing technologies. Sci. Rep. 2020, 10, 21935. [Google Scholar] [CrossRef]
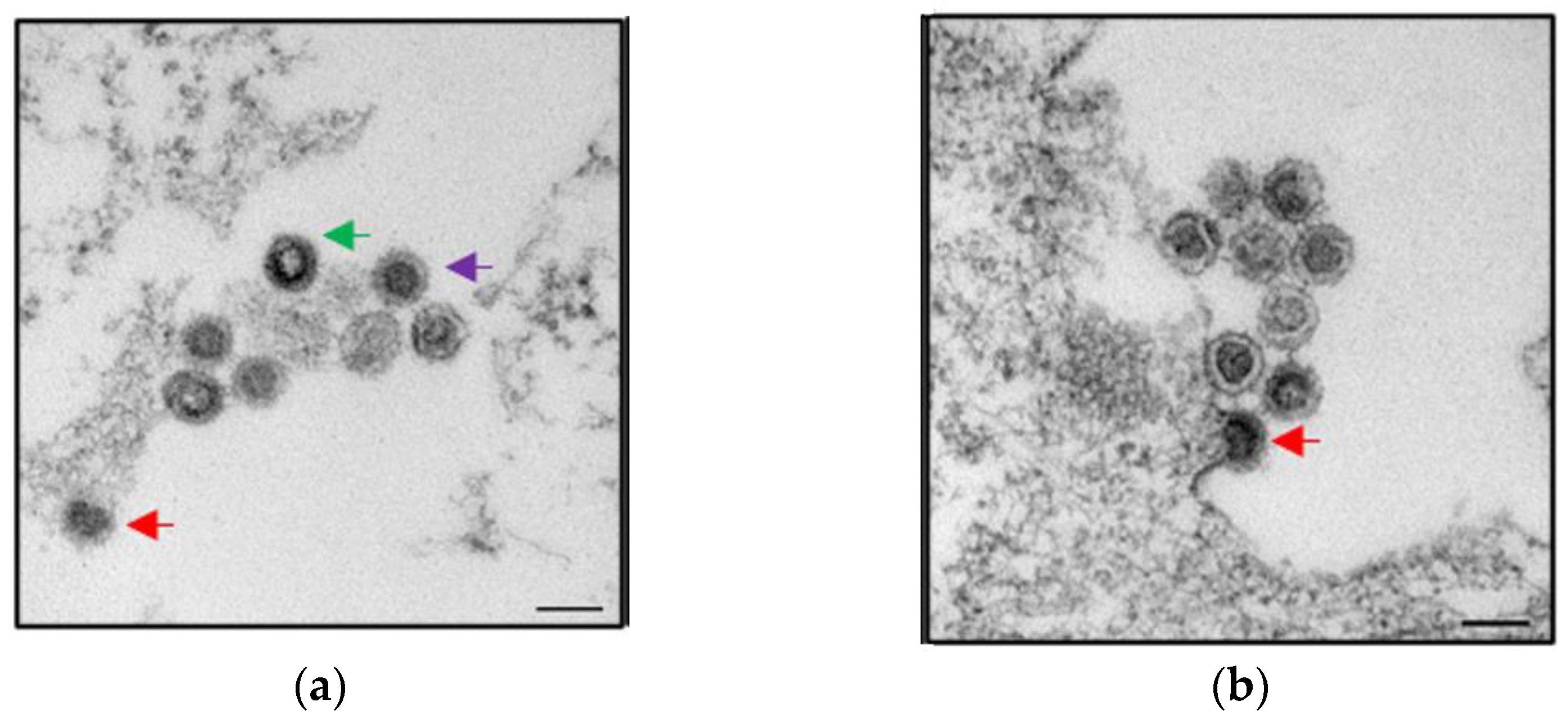
- Denner, J. How Active Are Porcine Endogenous Retroviruses (PERVs)? Viruses 2016, 8, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiebig, U.; Fischer, K.; Bähr, A.; Runge, C.; Schnieke, A.; Wolf, E.; Denner, J. Porcine endogenous retroviruses: Quantification of the copy number in cell lines, pig breeds, and organs. Xenotransplantation 2018, 25, e12445. [Google Scholar] [CrossRef]
- Krüger, L.; Stillfried, M.; Prinz, C.; Schröder, V.; Neubert, L.K.; Denner, J. Copy Number and Prevalence of Porcine Endogenous Retroviruses (PERVs) in German Wild Boars. Viruses 2020, 12, 419. [Google Scholar] [CrossRef] [Green Version]
- Krüger, L.; Nowak-Imialek, M.; Kristiansen, Y.; Herrmann, D.; Petersen, B.; Denner, J. Unexpected low expression of porcine endogenous retroviruses (PERVs) in porcine expanded potential stem cells (EPSCs). Virus Res. 2021, 294, 198295. [Google Scholar] [CrossRef] [PubMed]
- Meije, Y.; Tönjes, R.R.; Fishman, J.A. Retroviral Restriction Factors and Infectious Risk in Xenotransplantation. Am. J. Transplant. 2010, 10, 1511–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, E.H.; Jung, Y.T. Comparison of the effects of retroviral restriction factors involved in resistance to porcine endogenous ret-rovirus. J. Microbiol. Biotechnol. 2014, 24, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Bae, E.H.; Jung, Y.T. Tetherins of various species inhibit the release of porcine endogenous retrovirus from human cells. Acta Virol. 2014, 58, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Mattiuzzo, G.; Ivol, S.; Takeuchi, Y. Regulation of Porcine Endogenous Retrovirus Release by Porcine and Human Tetherins. J. Virol. 2010, 84, 2618–2622. [Google Scholar] [CrossRef] [Green Version]
- Al-Shehabi, H.; Fiebig, U.; Kutzner, J.; Denner, J.; Schaller, T.; Bannert, N.; Hofmann, H. Human SAMHD1 restricts the xenotransplantation relevant porcine endogenous retrovirus (PERV) in non-dividing cells. J. Gen. Virol. 2019, 100, 656–661. [Google Scholar] [CrossRef]
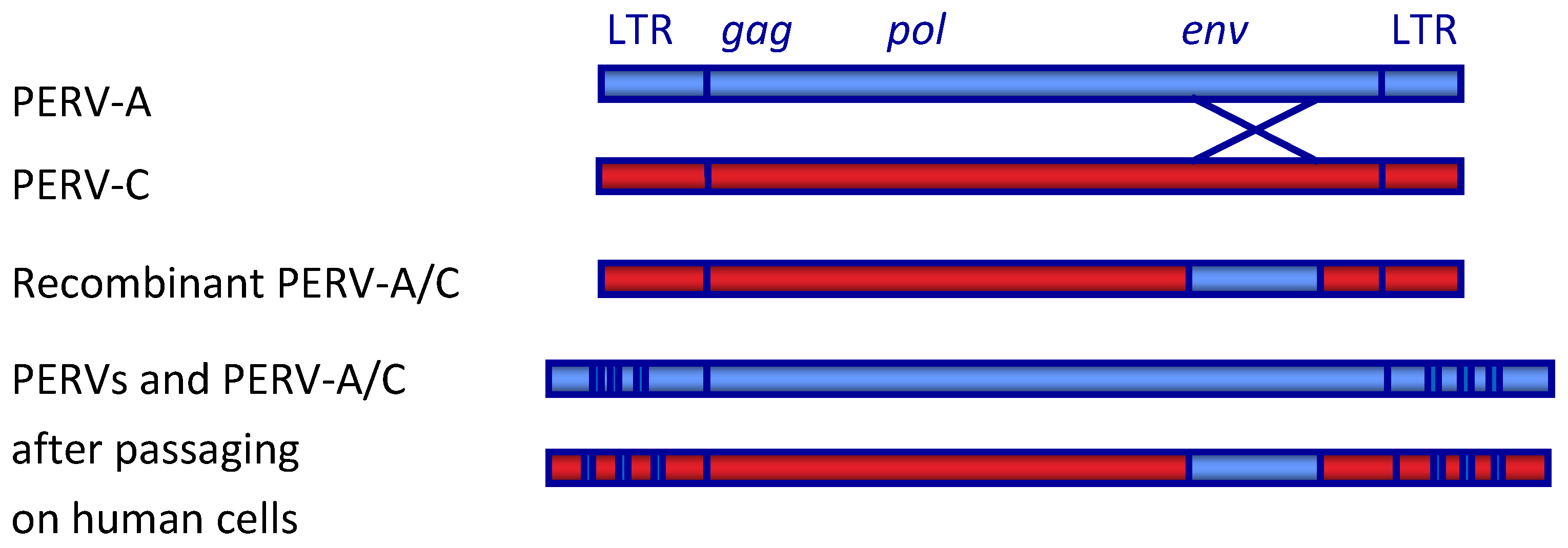
- Harrison, I.; Takeuchi, Y.; Bartosch, B.; Stoye, J.P. Determinants of High Titer in Recombinant Porcine Endogenous Retroviruses. J. Virol. 2004, 78, 13871–13879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartosch, B.; Stefanidis, D.; Myers, R.; Weiss, R.; Patience, C.; Takeuchi, Y. Evidence and Consequence of Porcine Endogenous Retrovirus Recombination. J. Virol. 2004, 78, 13880–13890. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.I.; Wilkinson, R.; Fishman, J.A. Genomic presence of recombinant porcine endogenous retrovirus in transmitting miniature swine. Virol. J. 2006, 3, 91. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.C.; Quinn, G.; Suling, K.M.; Oldmixon, B.A.; Van Tine, B.A.; Cina, R.; Arn, S.; Huang, C.A.; Scobie, L.; Onions, D.E.; et al. Identification of Exogenous Forms of Human-Tropic Porcine Endogenous Retrovirus in Miniature Swine. J. Virol. 2004, 78, 2494–2501. [Google Scholar] [CrossRef] [Green Version]
- Denner, J. Recombinant porcine endogenous retroviruses (PERV-A/C): A new risk for xenotransplantation? Arch. Virol. 2008, 153, 1421–1426. [Google Scholar] [CrossRef] [PubMed]
- Spizzo, T.; Denner, J.; Gazda, L.; Martin, M.; Nathu, D.; Scobie, L.; Takeuchi, Y. First update of the International Xenotransplantation Association consensus statement on conditions for undertaking clinical trials of porcine islet products in type 1 diabe-tes—Chapter 2a: Source pigs—Preventing xenozoonoses. Xenotransplantation 2016, 23, 25–31. [Google Scholar] [CrossRef]
- Denner, J.; Schuurmann, K.J. High prevalence of recombinant porcine endogenous retroviruses (PERV-A/Cs) in minipigs: A review on origin and presence. Viruses 2021, 13, 1869. [Google Scholar] [CrossRef] [PubMed]
- Pal, N.; Baker, R.; Schalk, S.; Scobie, L.; Tucker, A.W.; Opriessnig, T. Detection of Porcine Endogenous Retrovirus (PERV) Viremia in Diseased Versus Healthy US Pigs by Qualitative and Quantitative Real-Time RT-PCR. Transbound. Emerg. Dis. 2011, 58, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Dieckhoff, B.; Kessler, B.; Jobst, D.; Kues, W.; Petersen, B.; Pfeifer, A.; Kurth, R.; Niemann, H.; Wolf, E.; Denner, J. Distribution and expression of porcine endogenous retroviruses in multi-transgenic pigs generated for xenotransplantation. Xenotransplantation 2009, 16, 64–73. [Google Scholar] [CrossRef]
- Tacke, S.J.; Specke, V.; Denner, J. Differences in Release and Determination of Subtype of Porcine Endogenous Retroviruses Produced by Stimulated Normal Pig Blood Cells. Intervirology 2003, 46, 17–24. [Google Scholar] [CrossRef]
- Semaan, M.; Rotem, A.; Barkai, U.; Bornstein, S.; Denner, J. Screening pigs for xenotransplantation: Prevalence and expression of porcine endogenous retroviruses in Göttingen minipigs. Xenotransplantation 2013, 20, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Ma, Y.; Yan, Q.; Lv, M.; Zhao, X.; Yin, H.; Zhang, N.; Jia, J.; Yu, R.; Zhang, J. Construction and characterization of an infectious replication competent clone of porcine endogenous retrovirus from Chinese miniature pigs. Virol. J. 2013, 10, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büscher, K.; Terstegge, S.; Koch, P.; Itskovitz-Eldor, J.; Brüstle, O.; Denner, J. High level of expression of the human endogenous retrovirus HERV-K in human embryonic stem cells is decreased during differentiation. In Proceedings of the 24th Annual Meeting of the Society for Virology, Alps of Tyrol, Austria, 26–29 March 2014. [Google Scholar]
- Fuchs, N.V.; Loewer, S.; Daley, G.Q.; Izsvák, Z.; Löwer, J.; Löwer, R. Human endogenous retrovirus K (HML-2) RNA and protein expression is a marker for human embryonic and induced pluripotent stem cells. Retrovirology 2013, 10, 115. [Google Scholar] [CrossRef] [Green Version]
- Santoni, F.A.; Guerra, J.; Luban, J. HERV-H RNA is abundant in human embryonic stem cells and a precise marker for pluripo-tency. Retrovirology 2012, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Macfarlan, T.S.; Gifford, W.D.; Driscoll, S.; Lettieri, K.; Rowe, H.; Bonanomi, D.; Firth, A.; Singer, O.; Trono, D.; Pfaff, S.L. Embryonic stem cell potency fluctuates with endogenous retrovirus activity. Nature 2012, 487, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Godehardt, A.W.; Petkov, S.; Gulich, B.; Fischer, N.; Niemann, H.; Tonjes, R.R. Comparative gene expression profiling of pig-derived iPSC-like cells: Effects of induced pluripotency on expression of porcine endogenous retrovirus (PERV). Xenotransplantation 2018, 25, e12429. [Google Scholar] [CrossRef]
- Gao, X.; Nowak-Imialek, M.; Chen, X.; Chen, D.; Herrmann, D.; Ruan, D.; Chen, A.C.H.; Eckersley-Maslin, M.A.; Ahmad, S.; Lee, Y.L.; et al. Establishment of porcine and human expanded potential stem cells. Nat. Cell Biol. 2019, 21, 687–699. [Google Scholar] [CrossRef] [Green Version]
- Büscher, K.; Trefzer, U.; Hofmann, M.; Sterry, W.; Kurth, R.; Denner, J. Expression of Human Endogenous Retrovirus K in Melanomas and Melanoma Cell Lines. Cancer Res. 2005, 65, 4172–4180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezaei, S.; Hayward, J.; Norden, S.; Pedersen, J.; Mills, J.; Hearps, A.; Tachedjian, G. HERV-K Gag RNA and Protein Levels Are Elevated in Malignant Regions of the Prostate in Males with Prostate Cancer. Viruses 2021, 13, 449. [Google Scholar] [CrossRef]
- Salavatiha, Z.; Soleimani-Jelodar, R.; Jalilvand, S. The role of endogenous retroviruses-K in human cancer. Rev. Med. Virol. 2020, 30, 1–13. [Google Scholar] [CrossRef]
- Kassiotis, G. Endogenous Retroviruses and the Development of Cancer. J. Immunol. 2014, 192, 1343–1349. [Google Scholar] [CrossRef] [Green Version]
- Dieckhoff, B.; Puhlmann, J.; Büscher, K.; Hafner-Marx, A.; Herbach, N.; Bannert, N.; Büttner, M.; Wanke, R.; Kurth, R.; Denner, J. Expression of porcine endogenous retroviruses (PERVs) in melanomas of Munich miniature swine (MMS) Troll. Vet. Microbiol. 2007, 23, 53–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krüger, L.; Böttger, J.; Huang, C.A.; Denner, J. Absence of porcine endogenous retrovirus (PERV) production from pig lymphoma cell lines. Virus Res. 2021, 295, 198286. [Google Scholar] [CrossRef] [PubMed]
- Denner, J.; Schuurman, H.J.; Patience, C. The International Xenotransplantation Association consensus statement on conditions for undertaking clinical trials of porcine islet products in type 1 diabetes—Chapter 5: Strategies to prevent transmission of porcine endogenous retroviruses. Xenotransplantation 2009, 16, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Wynyard, S.; Nathu, D.; Garkavenko, O.; Denner, J.; Elliott, R. Microbiological safety of the first clinical pig islet xenotransplantation trial in New Zealand. Xenotransplantation 2014, 21, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.A.; Wynyard, S.; Matsumoto, S.; Abalovich, A.; Denner, J.; Elliott, R. No PERV transmission during a clinical trial of pig islet cell transplantation. Virus Res. 2017, 227, 34–40. [Google Scholar] [CrossRef]
- Valdes-Gonzalez, R.; Dorantes, L.M.; Bracho-Blanchet, E.; Rodrãguez-Ventura, A.; Djg, W. No evidence of porcine endogenous retrovirus in patients with type 1 diabetes after long-term porcine islet xenotransplantation. J. Med. Virol. 2010, 82, 331–334. [Google Scholar] [CrossRef]
- Plotzki, E.; Buerck, L.W.-V.; Knauf, Y.; Becker, T.; Maetz-Rensing, K.; Schuster, M.; Baehr, A.; Klymiuk, N.; Wolf, E.; Seissler, J.; et al. Virus safety of islet cell transplantation from transgenic pigs to marmosets. Virus Res. 2015, 204, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.A.; Ludwig, S.; Ludwig, B.; Rotem, A.; Barkai, U.; Bornstein, S.R.; Denner, J. Islet cell transplantation from Göttingen minipigs to cynomolgus monkeys: Analysis of virus safety. Xenotransplantation 2016, 23, 320–327. [Google Scholar] [CrossRef]
- Denner, J.; Längin, M.; Reichart, B.; Krüger, L.; Fiebig, U.; Mokelke, M.; Radan, J.; Mayr, T.; Milusev, A.; Luther, F.; et al. Impact of porcine cytomegalovirus on long-term orthotopic cardiac xenotransplant survival. Sci. Rep. 2020, 10, 17531. [Google Scholar] [CrossRef]
- Choi, H.J.; Kim, J.; Kim, J.Y.; Lee, H.J.; Wee, W.R.; Kim, M.K.; Hwang, E.S. Long-term safety from transmission of porcine endogenous retrovirus after pig-to-non-human primate corneal transplantation. Xenotransplantation 2017, 24, e12314. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.H.; Choi, H.J.; Kim, M.K. Corneal xenotransplantation: Where are we standing? Prog. Retin. Eye Res. 2021, 80, 100876. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Hirose, T.; Lassiter, G.; Sasaki, H.; Rosales, I.; Coe, T.M.; Rickert, C.G.; Matheson, R.; Colvin, R.B.; Qin, W.; et al. Kidney transplantation from triple-knockout pigs expressing multiple human proteins in cynomolgus macaques. Am. J. Transplant. 2021. [Google Scholar] [CrossRef]
- Ma, D.; Hirose, T.; Rosales, I.; Sasaki, H.; Colvin, R.; Markmann, J.; Qin, W.; Kan, Y.; Sasaki, H.; Kawai, T.; et al. Successful long-termTMA and rejection free survival of a kidney xenograft with triple xenoantigen knockout plus insertion of multiple human transgenes. Xenotransplantation 2019, 104, S82. [Google Scholar] [CrossRef]
- Adams, A.B.; Lovasik, B.P.; Faber, D.A.; Burlak, C.; Breeden, C.; Estrada, J.L.; Reyes, L.M.; Vianna, R.M.; Tector, M.F.; Tector, A.J. Anti-C5 Antibody Tesidolumab Reduces Early Antibody-mediated Rejection and Prolongs Survival in Renal Xenotransplantation. Ann. Surg. 2021, 274, 473–480. [Google Scholar] [CrossRef]
- Takamura, T.; Sasaki, H.; Hirayama, H.; Kiyoshi, A.; Inoue, M.; Matsui, K.; Matsumoto, N.; Saito, Y.; Fujimoto, T.; Tajiri, S.; et al. Techniques of orthotopic renal transplantation. II. Size-matched porcine grafts in monkey recipients. Acta Cirúrgica Bras. 2021, 36, e360503. [Google Scholar] [CrossRef]
- Hawthorne, W.J.; Salvaris, E.J.; Phillips, P.; Hawkes, J.; Liuwantara, D.; Burns, H.; Barlow, H.; Stewart, A.B.; Peirce, S.B.; Hu, M.; et al. Control of IBMIR in Neonatal Porcine Islet Xenotransplantation in Baboons. Am. J. Transplant. 2014, 14, 1300–1309. [Google Scholar] [CrossRef] [Green Version]
- Kaulitz, D.; Mihica, D.; Adlhoch, C.; Semaan, M.; Denner, J. Improved pig donor screening including newly identified variants of porcine endogenous retrovirus-C (PERV-C). Arch. Virol. 2013, 158, 341–348. [Google Scholar] [CrossRef]
- Kaulitz, D.; Mihica, D.; Dorna, J.; Costa, M.R.; Petersen, B.; Niemann, H.; Tönjes, R.R.; Denner, J. Development of sensitive methods for detection of porcine endogenous retrovirus-C (PERV-C) in the genome of pigs. J. Virol. Methods 2011, 175, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Ramsoondar, J.; Vaught, T.; Ball, S.; Mendicino, M.; Monahan, J.; Jobst, P.; Vance, A.; Duncan, J.; Wells, K.; Ayares, D. Production of transgenic pigs that express porcine endogenous retrovirus small interfering RNAs. Xenotransplantation 2009, 16, 164–180. [Google Scholar] [CrossRef] [PubMed]
- Karlas, A.; Kurth, R.; Denner, J. Inhibition of porcine endogenous retroviruses by RNA interference: Increasing the safety of xenotransplantation. Virology 2004, 325, 18–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieckhoff, B.; Karlas, A.; Hofmann, A.; Kues, W.A.; Petersen, B.; Pfeifer, A.; Niemann, H.; Kurth, R.; Denner, J. Inhibition of porcine endogenous retroviruses (PERVs) in primary porcine cells by RNA interference using lentiviral vectors. Arch. Virol. 2006, 152, 629–634. [Google Scholar] [CrossRef]
- Dieckhoff, B.; Petersen, B.; Kues, W.; Kurth, R.; Niemann, H.; Denner, J. Knockdown of porcine endogenous retrovirus (PERV) expression by PERV-specific shRNA in transgenic pigs. Xenotransplantation 2008, 15, 36–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semaan, M.; Kaulitz, D.; Petersen, B.; Niemann, H.; Denner, J. Long-term effects of PERV-specific RNA interference in transgenic pigs. Xenotransplantation 2012, 19, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Fiebig, U.; Stephan, O.; Kurth, R.; Denner, J. Neutralizing antibodies against conserved domains of p15E of porcine endogenous retroviruses: Basis for a vaccine for xenotransplantation? Virology 2003, 307, 406–413. [Google Scholar] [CrossRef] [Green Version]
- Kaulitz, D.; Fiebig, U.; Eschricht, M.; Wurzbacher, C.; Kurth, R.; Denner, J. Generation of neutralising antibodies against porcine endogenous retroviruses (PERVs). Virology 2011, 411, 78–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denner, J.; Mihica, D.; Kaulitz, D.; Schmidt, C.-M. Increased titers of neutralizing antibodies after immunization with both envelope proteins of the porcine endogenous retroviruses (PERVs). Virol. J. 2012, 9, 260. [Google Scholar] [CrossRef] [Green Version]
- Waechter, A.; Eschricht, M.; Denner, J. Neutralization of porcine endogenous retrovirus by antibodies against the membrane-proximal external region of the transmembrane envelope protein. J. Gen. Virol. 2013, 94 Pt 3, 643–651. [Google Scholar] [CrossRef]
- Waechter, A.; Denner, J. Novel neutralising antibodies targeting the N-terminal helical region of the transmembrane envelope protein p15E of the porcine endogenous retrovirus (PERV). Immunol. Res. 2014, 58, 9–19. [Google Scholar] [CrossRef]
- Semaan, M.; Ivanusic, D.; Denner, J. Cytotoxic Effects during Knock Out of Multiple Porcine Endogenous Retrovirus (PERV) Sequences in the Pig Genome by Zinc Finger Nucleases (ZFN). PLoS ONE 2015, 10, e0122059. [Google Scholar] [CrossRef] [Green Version]
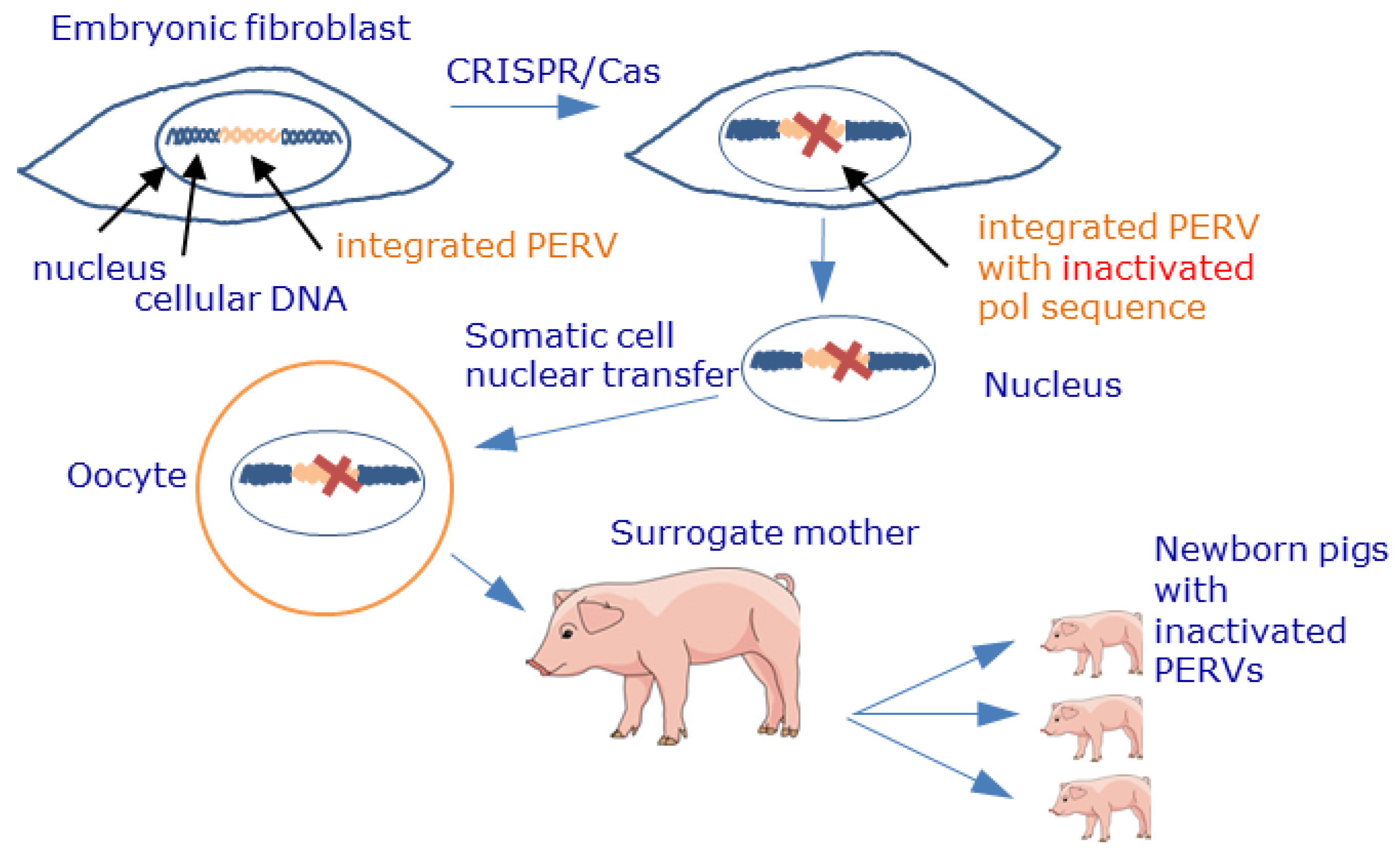
- Yang, L.; Güell, M.; Niu, D.; George, H.; Lesha, E.; Grishin, D.; Aach, J.; Shrock, E.; Xu, W.; Poci, J.; et al. Genome-wide inactivation of porcine endogenous retroviruses (PERVs). Science 2015, 350, 1101–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, D.; Wei, H.J.; Lin, L.; George, H.; Wang, T.; Lee, I.H.; Zhao, H.Y.; Wang, Y.; Kan, Y.; Shrock, E.; et al. Inactivation of porcine endogenous retrovirus in pigs using CRISPR-Cas9. Science 2017, 357, 1303–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denner, J. Can Antiretroviral Drugs Be Used to Treat Porcine Endogenous Retrovirus (PERV) Infection after Xenotransplantation? Viruses 2017, 9, 213. [Google Scholar] [CrossRef] [Green Version]
- Argaw, T.; Colon-Moran, W.; Wilson, C. Susceptibility of porcine endogenous retrovirus to anti-retroviral inhibitors. Xenotransplantation 2016, 23, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Demange, A.; Yajjou-Hamalian, H.; Gallay, K.; Luengo, C.; Beven, V.; Leroux, A.; Confort, M.P.; Al Andary, E.; Gouet, P.; Moreau, K.; et al. Porcine endogenous retrovirus-A/C: Biochemical properties of its integrase and susceptibility to raltegravir. J. Gen. Virol. 2015, 96, 3124–3130. [Google Scholar] [CrossRef]
- Qari, S.H.; Magre, S.; García-Lerma, J.G.; Hussain, A.I.; Takeuchi, Y.; Patience, C.; Weiss, R.A.; Heneine, W. Susceptibility of the Porcine Endogenous Retrovirus to Reverse Transcriptase and Protease Inhibitors. J. Virol. 2001, 75, 1048–1053. [Google Scholar] [CrossRef] [Green Version]
- Powell, S.K.; Gates, M.E.; Langford, G.; Gu, M.-L.; Lockey, C.; Long, Z.; Otto, E. Antiretroviral Agents Inhibit Infection of Human Cells by Porcine Endogenous Retroviruses. Antimicrob. Agents Chemother. 2000, 44, 3432–3433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephan, O.; Schwendemann, J.; Specke, V.; Tacke, S.J.; Boller, K.; Denner, J. Porcine endogenous retroviruses (PERVs): Generation of specific antibodies, development of an immunoperoxidase assay (IPA) and inhibition by AZT. Xenotransplantation 2001, 8, 310–316. [Google Scholar] [CrossRef]
- Richman, D.D. HIV drug resistance. Annu. Rev. Pharmacol. Toxicol. 1993, 33, 149–164. [Google Scholar] [CrossRef]
- Sparkes, A.H. Feline leukaemia virus and vaccination. J. Feline Med. Surg. 2003, 5, 97–100. [Google Scholar] [CrossRef]
- Bayer, W.; Schimmer, S.; Hoffmann, D.; Dittmer, U.; Wildner, O. Evaluation of the Friend Virus model for the development of improved adenovirus-vectored anti-retroviral vaccination strategies. Vaccine 2008, 26, 716–726. [Google Scholar] [CrossRef] [PubMed]
- Weinhold, K.J.; Huper, G.; Matthews, T.J.; Fischinger, P.J.; Ihle, J.N.; Schwarz, H.; Thiel, H.-J.; Schäfer, W.; Bolognesi, D.P. Properties of mouse leukemia viruses XIX. Effective antibody therapy of AKR leukemia occurs independently of virus neutralization and produces long-term changes in the virus status of the thymus. Virology 1984, 135, 105–117. [Google Scholar] [CrossRef]
- Denner, J. Immunising with the transmembrane envelope proteins of different retroviruses including HIV-1: A comparative study. Hum. Vaccines Immunother. 2013, 9, 462–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langhammer, S.; Hübner, J.; Jarrett, O.; Kurth, R.; Denner, J. Immunization with the transmembrane protein of a retrovirus, feline leukemia virus: Absence of antigenemia following challenge. Antivir. Res. 2011, 89, 119–123. [Google Scholar] [CrossRef]
- Keller, M.; Petersen, B.; Niemann, H.; Denner, J. Lack of antibody response in pigs immunized with the transmembrane envelope protein of porcine endogenous retroviruses. J. Gen. Virol. 2014, 95 Pt 8, 1827–1831. [Google Scholar] [CrossRef] [Green Version]
- Denner, J.; Petersen, B.; Niemann, H. Tolerance and immune response to the porcine endogenous retrovirus in German landrace pigs immunised with viral proteins. Virus Res. 2015, 208, 39–43. [Google Scholar] [CrossRef]
- Tanihara, F.; Hirata, M.; Otoi, T. Current status of the application of gene editing in pigs. J. Reprod. Dev. 2021, 67, 177–187. [Google Scholar] [CrossRef]
- Godehardt, A.W.; Fischer, N.; Rauch, P.; Gulich, B.; Boller, K.; Church, G.M.; Tönjes, R.R. Characterization of porcine endogenous retrovirus particles released by the CRISPR/Cas9 inactivated cell line PK15 clone. Xenotransplantation 2020, 27, e12563. [Google Scholar] [CrossRef]
- Denner, J.; Scobie, L.; Schuurman, H.-J. Is it currently possible to evaluate the risk posed by PERVs for clinical xenotransplantation? Xenotransplantation 2018, 25, e12403. [Google Scholar] [CrossRef] [Green Version]
- Güell, M.; Niu, D.; Kan, Y.; George, H.; Wang, T.; Wang, G.; Church, G.; Yang, L.; Lee, I.-H. PERV inactivation is necessary to guarantee absence of pig-to-patient PERVs transmission in xenotransplantation. Xenotransplantation 2017, 24, e12366. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. Paving the Path toward Porcine Organs for Transplantation. N. Engl. J. Med. 2017, 377, 1891–1893. [Google Scholar] [CrossRef] [PubMed]
- Scobie, L.; Denner, J.; Schuurman, H.-J. Inactivation of porcine endogenous retrovirus in pigs using CRISPR-Cas9, editorial commentary. Xenotransplantation 2017, 24, e12363. [Google Scholar] [CrossRef]
- Denner, J. Advances in organ transplant from pigs. Science 2017, 357, 1238–1239. [Google Scholar] [CrossRef] [PubMed]
- Garkavenko, O.; Dieckhoff, B.; Wynyard, S.; Denner, J.; Elliott, R.B.; Tan, P.L.; Croxson, M.C. Absence of transmission of potentially xenotic viruses in a prospective pig to primate islet xenotransplantation study. J. Med. Virol. 2008, 80, 2046–2052. [Google Scholar] [CrossRef]
- Moscoso, I.; Hermida-Prieto, M.; Mañez, R.; Lopez-Pelaez, E.; Centeno, A.; Diaz, T.M.; Domenech, N. Lack of Cross-Species Transmission of Porcine Endogenous Retrovirus in Pig-to-Baboon Xenotransplantation with Sustained Depletion of anti-alphagal Antibodies. Transplantation 2005, 79, 777–782. [Google Scholar] [CrossRef]
- Nishitai, R.; Ikai, I.; Shiotani, T.; Katsura, N.; Matsushita, T.; Yamanokuchi, S.; Matsuo, K.; Sugimoto, S.; Yamaoka, Y. Absence of PERV infection in baboons after transgenic porcine liver perfusion. J. Surg. Res. 2005, 124, 45–51. [Google Scholar] [CrossRef]
- Martin, U.; Tacke, S.J.; Simon, A.R.; Schroder, C.; Wiebe, K.; Lapin, B.; Haverich, A.; Denner, J.; Steinhoff, G. Absence of PERV specific humoral immune response in baboons after transplantation of porcine cells or organs. Transpl. Int. 2002, 15, 361–368. [Google Scholar] [CrossRef]
- Denner, J.; Graham, M. Xenotransplantation of islet cells: What can the non-human primate model bring for the evaluation of efficacy and safety? Xenotransplantation 2015, 22, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Denner, J.; Specke, V.; Karlas, A.; Chodnevskaja, I.; Meyer, T.; Moskalenko, V.; Kurth, R.; Ulrichs, K. No transmission of porcine endogenous retroviruses (PERVs) in a long-term pig to rat xenotransplantation model and no infection of immunosuppressed rats. Ann. Transplant. 2008, 13, 20–31. [Google Scholar]
- Rodríguez, T.C.; Dadafarin, S.; Pratt, H.E.; Liu, P.; Amrani, N.; Zhu, L.J. Genome-wide detection and analysis of CRISPR-Cas off-targets. Prog. Mol. Biol. Transl. Sci. 2021, 181, 31–43. [Google Scholar] [CrossRef]
- Aryal, N.; Wasylishen, A.; Lozano, G. CRISPR/Cas9 can mediate high-efficiency off-target mutations in mice In Vivo. Cell Death Dis. 2018, 9, 1099. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.-H.; Deng, W.-Y.; Jiang, H.-T.; Li, T.; Wang, Y. Kidney xenotransplantation: Recent progress in preclinical research. Clin. Chim. Acta 2021, 514, 15–23. [Google Scholar] [CrossRef] [PubMed]

| Recipient | Pig Transplant | Immunosuppression | PERV Screening | Reference | |
|---|---|---|---|---|---|
| In the Donor Pig | In the Recipient | ||||
| Rhesus monkeys | Fresh or decellularized cornea from SNU minipigs | Methylprednisolone, IVIG, Anti-CD40, or anti-CD154 | PERV | No (PCR *) | [106] |
| Baboon | Orthotopic heart from GTKO, CD46, hTM pigs | anti-CD20, anti-CD40, MMF, methylpredni-solone | PERV-A, PERV-B, PERV-C | No (PCR) | [105] |
| Streptozoto-cin-induced diabetic cynomolgus macaques | Encapsulated islets from Large White-Yorkshire × Landrace F1 hybrids | None | PERV-A, PERV-B, PERV-C | No (PCR) | [63] |
| Nondiabetic cynomolgus macaques | Göttingen minipigs | None | PERV-A, PERV-B, PERV-C | No (PCR, Western blot) | [104] |
| Normo-glycaemic marmosets | Islet cells from German landrace hybrid pigs expressing LEA29Y | None | PERV-A, PERV-B, PERV-C | No (PCR, Western blot) | [103] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denner, J. Porcine Endogenous Retroviruses and Xenotransplantation, 2021. Viruses 2021, 13, 2156. https://doi.org/10.3390/v13112156
Denner J. Porcine Endogenous Retroviruses and Xenotransplantation, 2021. Viruses. 2021; 13(11):2156. https://doi.org/10.3390/v13112156
Chicago/Turabian StyleDenner, Joachim. 2021. "Porcine Endogenous Retroviruses and Xenotransplantation, 2021" Viruses 13, no. 11: 2156. https://doi.org/10.3390/v13112156
APA StyleDenner, J. (2021). Porcine Endogenous Retroviruses and Xenotransplantation, 2021. Viruses, 13(11), 2156. https://doi.org/10.3390/v13112156