
A 10-Year Retrospective Study of Inclusion Body Hepatitis in Meat-Type Chickens in Spain (2011–2021)

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Cases
2.2. Diagnosis of Suspected IBH Cases
2.3. Statistical Analysis
2.4. Phylogenetic Analysis
3. Results
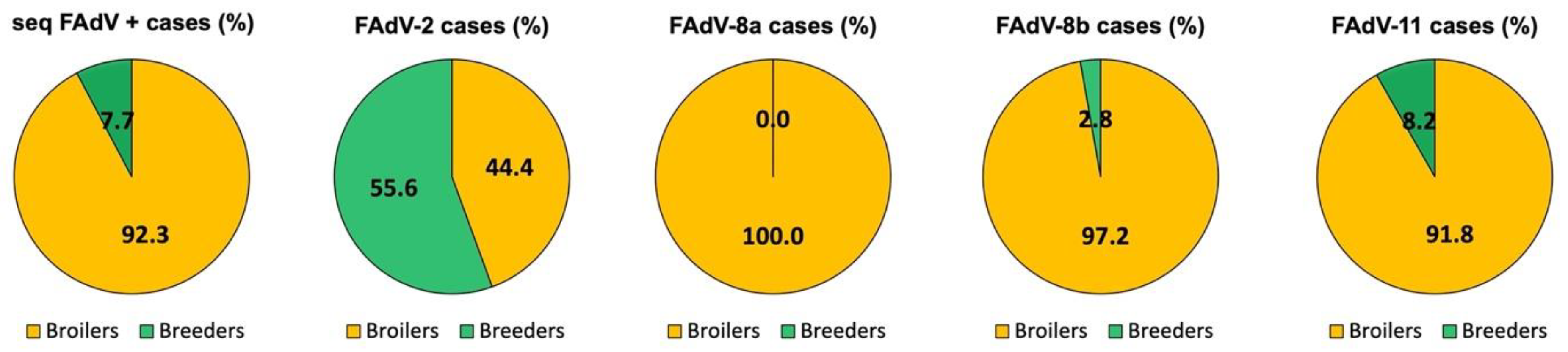
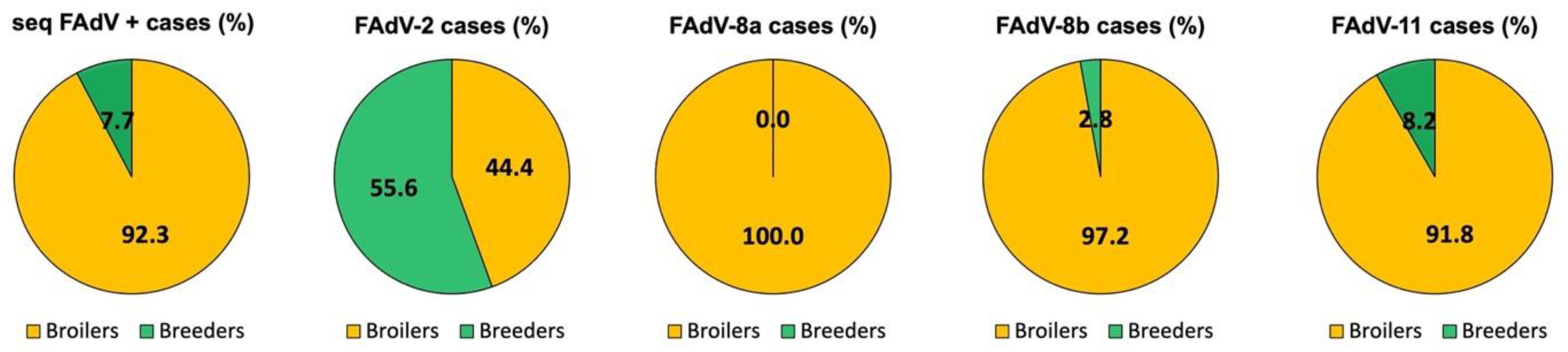
3.1. Distribution of IBH Cases Based on Poultry Type
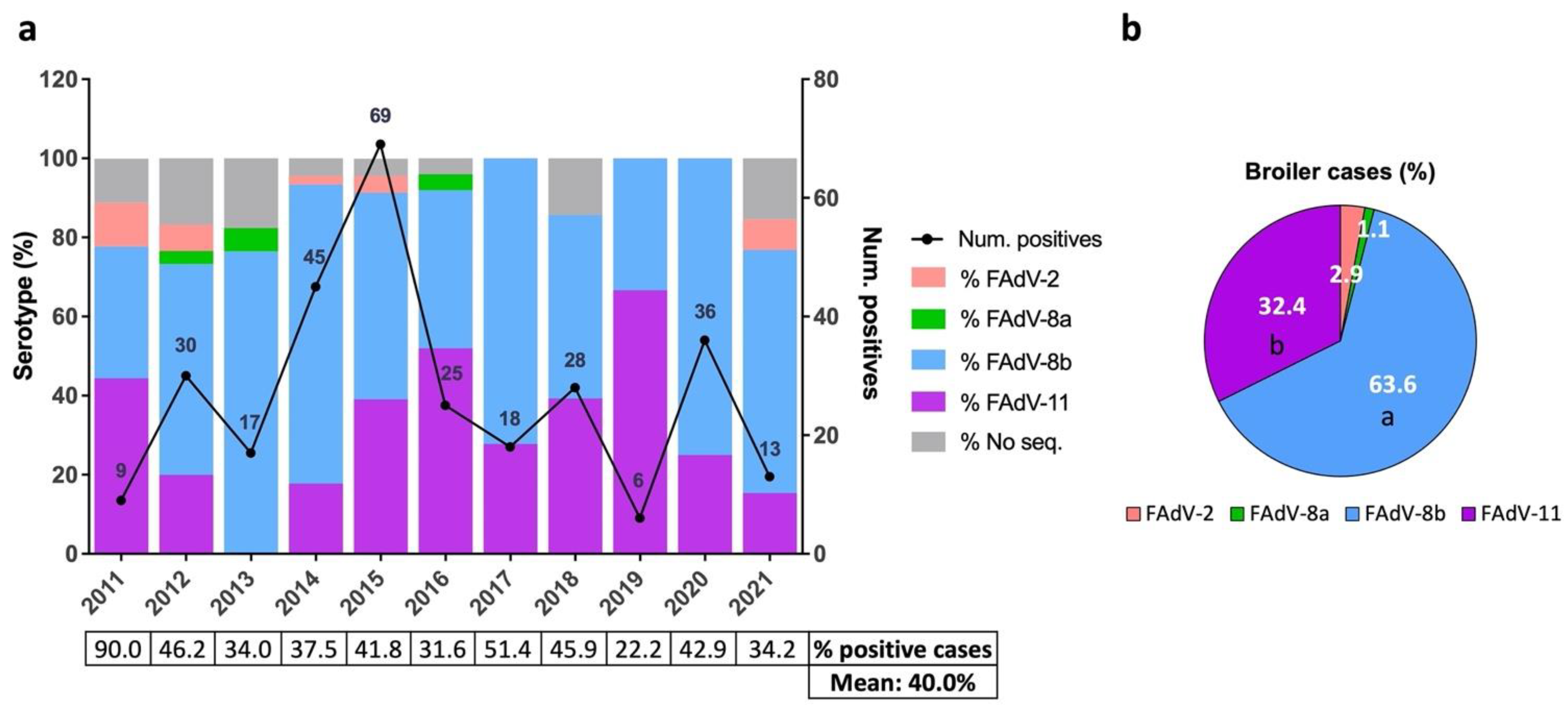
3.2. Distribution of IBH Cases in Broilers
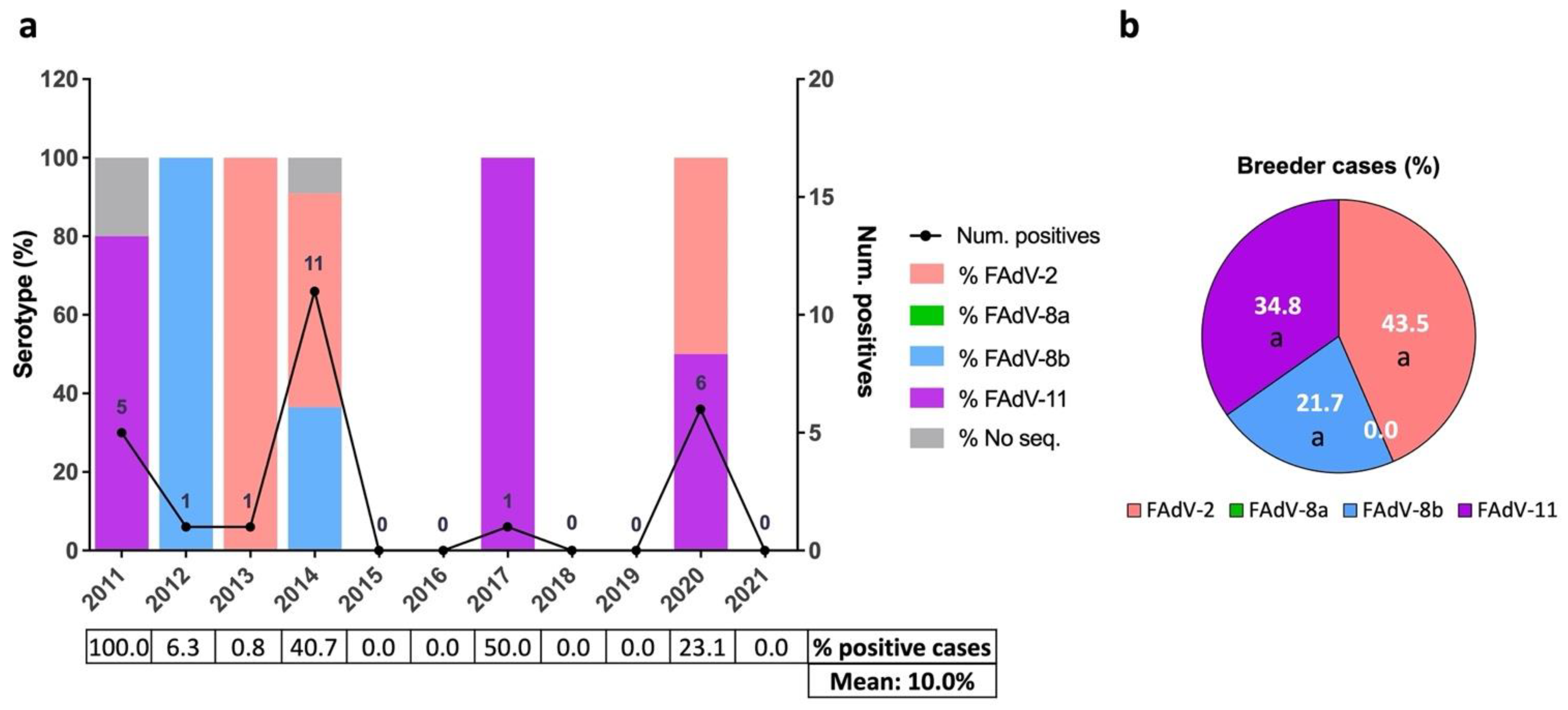
3.3. Distribution of IBH Cases in Broiler Breeder Pullets
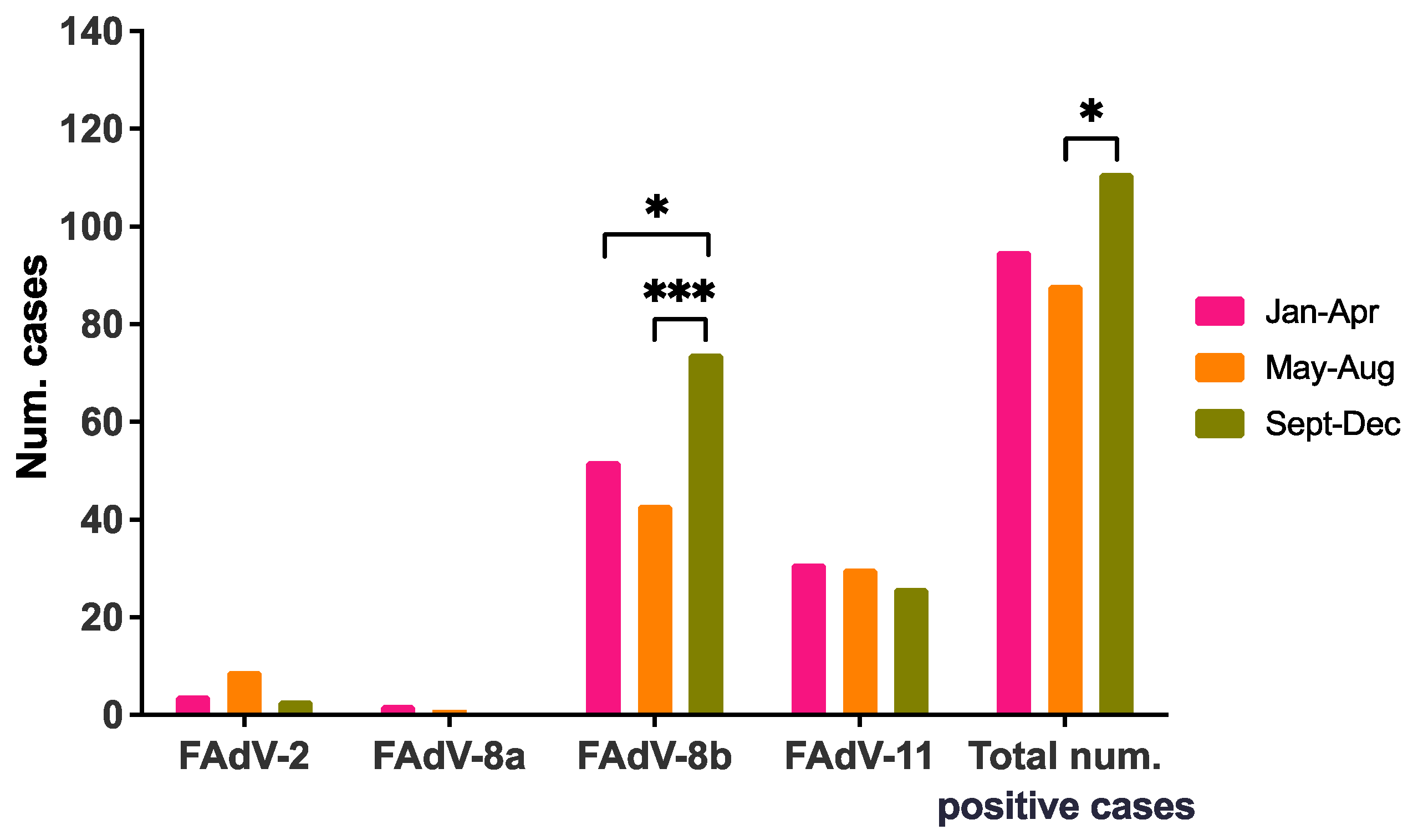
3.4. Intra-Annual Seasonality of IBH Cases
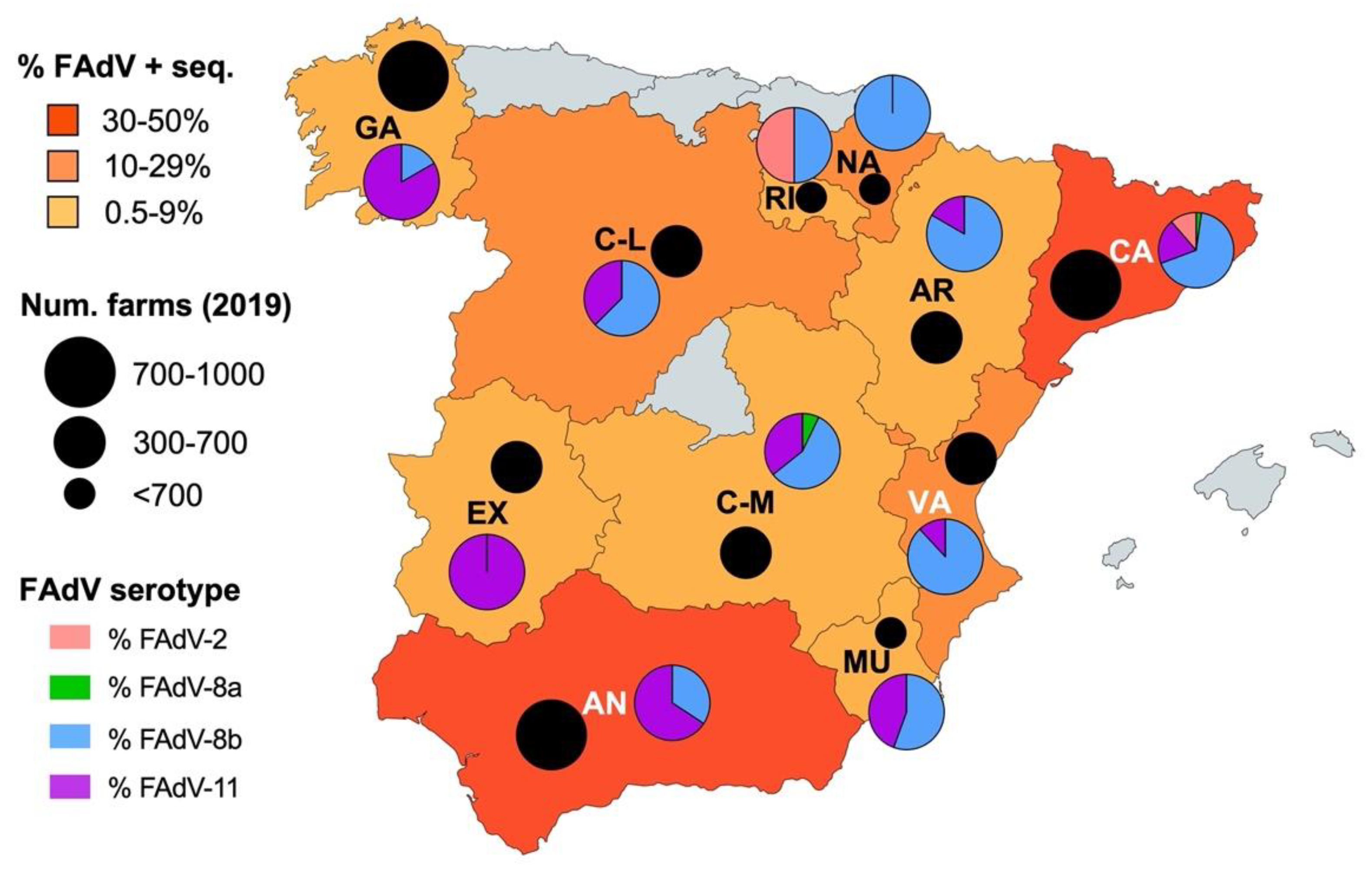
3.5. Geographic Distribution of IBH Cases
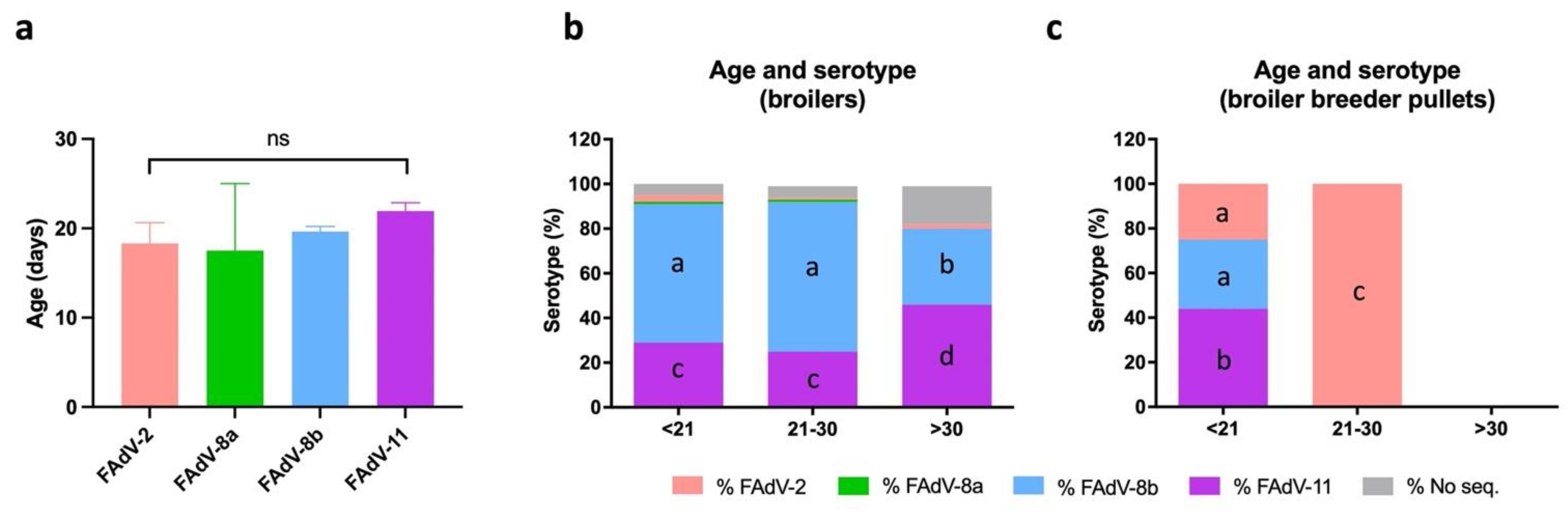
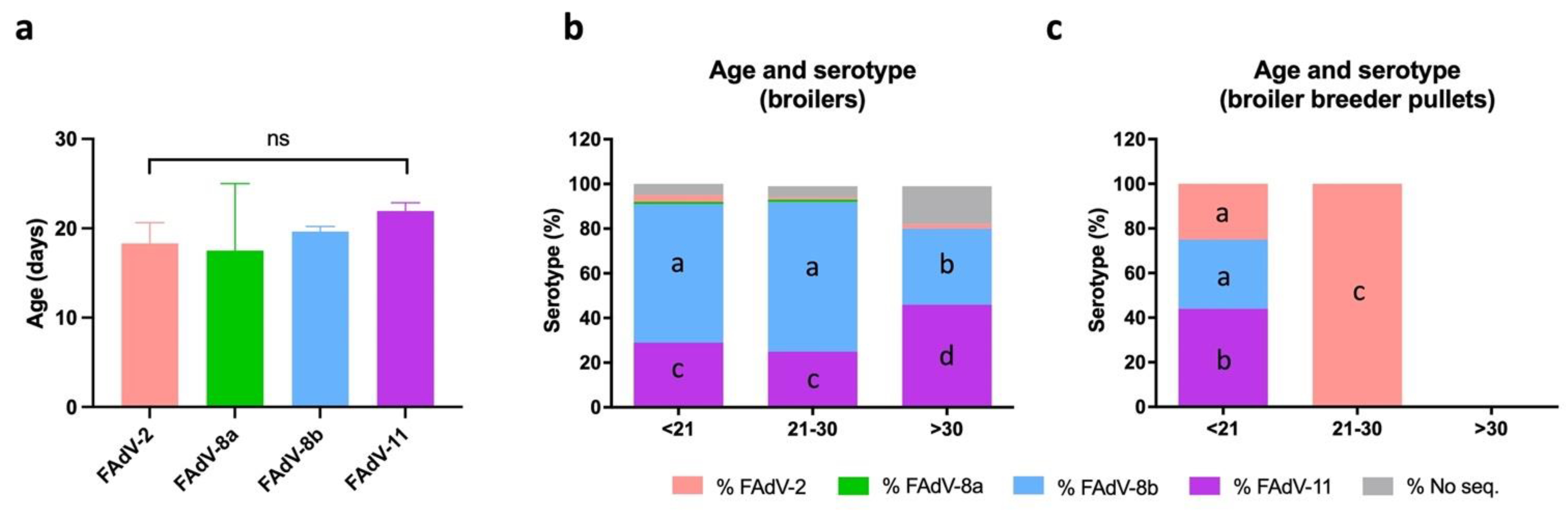
3.6. Distribution of IBH Cases Based on Age
3.7. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hess, M. Aviadenovirus infections. In Diseases of Poultry Swayne; Boulianne, M.D.E., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 322–331. [Google Scholar]
- Matos, M.; Grafl, B.; Liebhart, D.; Hess, M. The outcome of experimentally induced inclusion body hepatitis (IBH) by fowl aviadenoviruses (FAdVs) is crucially influenced by the genetic background of the host. Vet. Res. 2016, 47, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schachner, A.; Matos, M.; Grafl, B.; Hess, M. Fowl Adenovirus-Induced Diseases and Strategies for Their Control—A Review on the Current Global Situation. Avian Pathol. 2018, 47, 111–126. [Google Scholar] [CrossRef]
- Ayala, A.J.; Yabsley, M.J.; Hernandez, S.M. A Review of Pathogen Transmission at the Backyard Chicken–Wild Bird Interface. Front. Vet. Sci. 2020, 7, 539925. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Kumar, V.; Asthana, M.; Shukla, S.K.; Chandra, R. Isolation and Identification of a Fowl Adenovirus from Wild Black Kites (Milvus migrans). J. Wildl. Dis. 2010, 46, 272–276. [Google Scholar] [CrossRef]
- Schachner, A.; Grafl, B.; Hess, M. Spotlight on Avian Pathology: Fowl Adenovirus (FAdV) in Chickens and beyond—An Unresolved Host-Pathogen Interplay. Avian Pathol. 2021, 50, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Franzo, G.; Prentza, Z.; Paparounis, T.; Tsiouris, V.; Centonze, G.; Legnardi, M.; Catelli, E.; Tucciarone, C.M.; Koutoulis, K.; Cecchinato, M. Molecular epidemiology of fowl adenoviruses in Greece. Poult. Sci. 2020, 99, 5983–5990. [Google Scholar] [CrossRef]
- Chen, L.; Yin, L.; Zhou, Q.; Peng, P.; Du, Y.; Liu, L.; Zhang, Y.; Xue, C.; Cao, Y. Epidemiological investigation of fowl adenovirus infections in poultry in China during 2015–2018. BMC Vet. Res. 2019, 15, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Radwan, M.M.; El-Deeb, A.H.; Mousa, M.R.; El-Sanousi, A.A.; Shalaby, M.A. First report of fowl adenovirus 8a from commercial broiler chickens in Egypt: Molecular characterization and pathogenicity. Poult. Sci. 2019, 98, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Abghour, S.; Zro, K.; Mouahid, M.; Tahiri, F.; Tarta, M.; Berrada, J.; Kichou, F. Isolation and characterization of fowl aviadenovirus serotype 11 from chickens with inclusion body hepatitis in Morocco. PLoS ONE 2019, 14, e0227004. [Google Scholar] [CrossRef]
- Mase, M.; Hiramatsu, K.; Nishijima, N.; Iguchi, H.; Honda, S.; Hanyu, S.; Iseki, H.; Watanabe, S. Fowl Adenoviruses Type 8b Isolated from Chickens with Inclusion Body Hepatitis in Japan. Avian Dis. 2020, 64, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Mete, A.; Armien, A.G.; Rejmanek, D.; Mott, M.; Crossley, B.M. Emergence of fowl aviadenovirus C-4 in a backyard chicken flock in California. J. Vet. Diagn. Investig. 2021, 33, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Niczyporuk, J.S.; Kozdrun, W.; Czekaj, H.; Piekarska, K.; Stys-Fijol, N. Characterisation of adenovirus strains represented species B and E isolated from broiler chicken flocks in eastern Poland. Heliyon 2021, 7, e06225. [Google Scholar] [CrossRef]
- Chitradevi, S.; Sukumar, K.; Suresh, P.; Balasubramaniam, G.A.; Kannan, D. Molecular typing and pathogenicity assessment of fowl adenovirus associated with inclusion body hepatitis in chicken from India. Trop. Anim. Health Prod. 2021, 53, 1–12. [Google Scholar] [CrossRef] [PubMed]
- De la Torre, D.; Nuñez, L.F.N.; Parra, S.H.S.; Astolfi-Ferreira, C.S.; Ferreira, A.J.P. Molecular characterization of fowl adenovirus group I in commercial broiler chickens in Brazil. VirusDisease 2018, 29, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Oliver-Ferrando, S.; Dolz, R.; Calderon, C.; Valle, R.; Rivas, R.; Pérez, M.; Biarnés, M.; Blanco, A.; Bertran, K.; Ramis, A.; et al. Epidemiological and pathological investigation of fowl aviadenovirus serotypes 8b and 11 isolated from chickens with inclusion body hepatitis in Spain (2011–2013). Avian Pathol. 2016, 46, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raue, R.; Gerlach, H.; Müller, H. Phylogenetic analysis of the hexon loop 1 region of an adenovirus from psittacine birds supports the existence of a new psittacine adenovirus (PsAdV). Arch. Virol. 2005, 150, 1933–1943. [Google Scholar] [CrossRef]
- Subdirección General de Producciones Ganaderas y Cinegéticas; Dirección General de Producciones y Mercados Agrarios. El Sector de La Avicultura de Carne En Cifras: Principales Indicadores Económicos 2019; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2020.
- Committee of Professional Agricultural Organisations and General Confederation of Agricultural Co-operatives in the European Union (COPA-COGECA); Association of Poultry Processors and Poultry Trade in the EU (AVEC). Guide to Good Hygiene Practice for the Prevention and Control of Pathogenic Microorganisms with Particular Reference to Salmonella in Gallus Gallus (Broilers) Reared for Meat—on Farms, and during Catching, Loading and Transport. Official Journal of the European Union, 2010. [Google Scholar]
- Official Journal of the European Union. European Commission Commission Regulation (EU) No 200/2010 of 10 March 2010 Implementing Regulation (EC) No 2160/2003 of the European Parliament and of the Council as Regards a Union Target for the Reduction of the Prevalence of Salmonella Serotypes in Adult Breeding Flocks of Gallus Gallus. Official Journal of the European Union, 2010. [Google Scholar]
- Kiss, I.; Homonnay, Z.; Mató, T.; Bányai, K.; Palya, V. Research Note: An overview on distribution of fowl adenoviruses. Poult. Sci. 2021, 100, 101052. [Google Scholar] [CrossRef] [PubMed]
- Morshed, R.; Hosseini, H.; Langeroudi, A.G.; Fard, M.H.B.; Charkhkar, S. Fowl Adenoviruses D and E Cause Inclusion Body Hepatitis Outbreaks in Broiler and Broiler Breeder Pullet Flocks. Avian Dis. 2017, 61, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Mirzazadeh, A.; Grafl, B.; Berger, E.; Schachner, A.; Hess, M. Longitudinal Serological Monitoring of Commercial Broiler Breeders for Fowl Adenoviruses (FAdVs)—Presence of Antibodies Is Linked with Virus Excretion. Avian Dis. 2020, 65, 177–187. [Google Scholar] [CrossRef]
- Alvarado, I.R.; Villegas, P.; El-Attrache, J.; Jensen, E.; Rosales, G.; Perozo, F.; Purvis, L.B. Genetic Characterization, Pathogenicity, and Protection Studies with an Avian Adenovirus Isolate Associated with Inclusion Body Hepatitis. Avian Dis. 2007, 51, 27–32. [Google Scholar] [CrossRef]
- Gupta, A.; Popowich, S.; Ojkic, D.; Kurukulasuriya, S.; Chow-Lockerbie, B.; Gunawardana, T.; Goonewardene, K.; Karunarathna, R.; Ayalew, L.E.; Ahmed, K.A.; et al. Inactivated and live bivalent fowl adenovirus (FAdV8b + FAdV11) breeder vaccines provide broad-spectrum protection in chicks against inclusion body hepatitis (IBH). Vaccine 2017, 36, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-S.; Lim, T.-H.; Lee, D.-H.; Youn, H.-N.; Yuk, S.-S.; Kim, B.-Y.; Choi, S.-W.; Jung, C.-H.; Han, J.-H.; Song, C.-S. An inactivated oil-emulsion fowl Adenovirus serotype 4 vaccine provides broad cross-protection against various serotypes of fowl Adenovirus. Vaccine 2014, 32, 3564–3568. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, S.; Zou, K.; Zhang, Y.; Xu, S.; Yin, Y. Variant Serotypes of Fowl Adenovirus Isolated from Commercial Poultry Between 2007 and 2017 in Some Regions of China. Avian Dis. 2018, 62, 171–176. [Google Scholar] [CrossRef]
- Lai, V.D.; Min, K.; Lai, H.T.L.; Mo, J. Epidemiology of fowl adenovirus (FAdV) infections in South Korean chickens during 2013–2019 following introduction of FAdV-4 vaccines. Avian Pathol. 2021, 50, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L. Average Monthly Rainfall in Spain 2020. 2021. Available online: https://www.statista.com/statistics/802729/average-monthly-rainfall-spain/ (accessed on 27 October 2021).
- Fernández, L. Average Monthly Temperature in Spain 2020. 2021. Available online: https://www.statista.com/statistics/802681/average-monthly-temperature-spain/ (accessed on 27 October 2021).
- Singh, A.; Oberoi, M.; Jand, S.; Singh, B. Epidemiology of inclusion body hepatitis in poultry in northern India from 1990 to 1994. Rev. Sci. Tech. l’OIE 1996, 15, 1053–1060. [Google Scholar] [CrossRef] [Green Version]
- Mittal, D.; Jindal, N.; Khokhar, R.S. Epidemiological Studies on Inclusion Body Hepatitis-Hydropericardium Syndrome in Broiler Chicken in Haryana State. Haryana Vet. 2014, 53, 34–38. [Google Scholar]
- Niu, Y.; Sun, Q.; Zhang, G.; Sun, W.; Liu, X.; Xiao, Y.; Shang, Y.; Liu, S. Epidemiological investigation of outbreaks of fowl adenovirus infections in commercial chickens in China. Transbound. Emerg. Dis. 2017, 65, e121–e126. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertran, K.; Blanco, A.; Antilles, N.; Nofrarías, M.; Valle, R.M.; Cobos, À.; Ramis, A.; Biarnés, M.; Majó, N. A 10-Year Retrospective Study of Inclusion Body Hepatitis in Meat-Type Chickens in Spain (2011–2021). Viruses 2021, 13, 2170. https://doi.org/10.3390/v13112170
Bertran K, Blanco A, Antilles N, Nofrarías M, Valle RM, Cobos À, Ramis A, Biarnés M, Majó N. A 10-Year Retrospective Study of Inclusion Body Hepatitis in Meat-Type Chickens in Spain (2011–2021). Viruses. 2021; 13(11):2170. https://doi.org/10.3390/v13112170
Chicago/Turabian StyleBertran, Kateri, Angela Blanco, Noelia Antilles, Miquel Nofrarías, Rosa M. Valle, Àlex Cobos, Antonio Ramis, Mar Biarnés, and Natàlia Majó. 2021. "A 10-Year Retrospective Study of Inclusion Body Hepatitis in Meat-Type Chickens in Spain (2011–2021)" Viruses 13, no. 11: 2170. https://doi.org/10.3390/v13112170
APA StyleBertran, K., Blanco, A., Antilles, N., Nofrarías, M., Valle, R. M., Cobos, À., Ramis, A., Biarnés, M., & Majó, N. (2021). A 10-Year Retrospective Study of Inclusion Body Hepatitis in Meat-Type Chickens in Spain (2011–2021). Viruses, 13(11), 2170. https://doi.org/10.3390/v13112170