
Macrophages and Monocytes: “Trojan Horses” in COVID-19

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. SARS-CoV-2 Variants
2.2. Cell Culture
2.3. Flow Cytometry
2.4. Direct Infection of MDM and MN
2.5. MDM and MN Mediated Transmission of the Infectious Virus through Co-Culture with Infected VERO E6
2.6. Virus Transmission Assay
2.7. Kinetic of N and S Protein Expression and RNA Replication on MDM, MN and VERO E6
2.8. Mechanism of Virus Entry into MDM and MN
2.9. Light Microscopy and Light Microscopy Immunohistochemistry
2.10. Ultrastructural Study
2.11. Statistical Analysis
3. Results
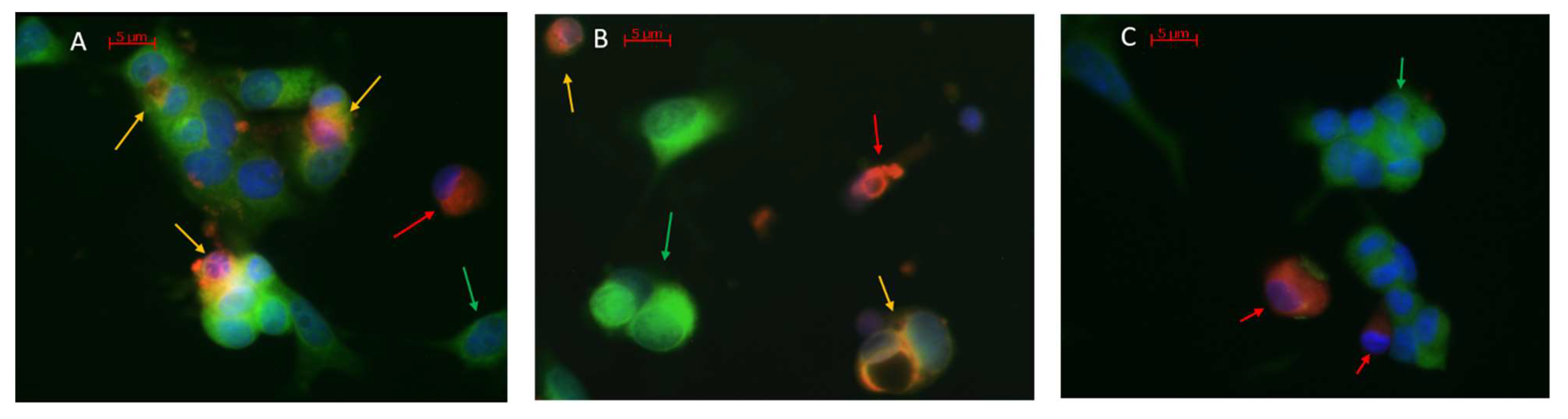
3.1. Analysis of Macrophages and Monocytes Infected with Different SARS-CoV-2 Variants
3.2. Analysis of MDM and MN Infection Co-Cultured with Infected VERO E6
3.3. Virus-Induced Fluorescent Probe Transmission Assay
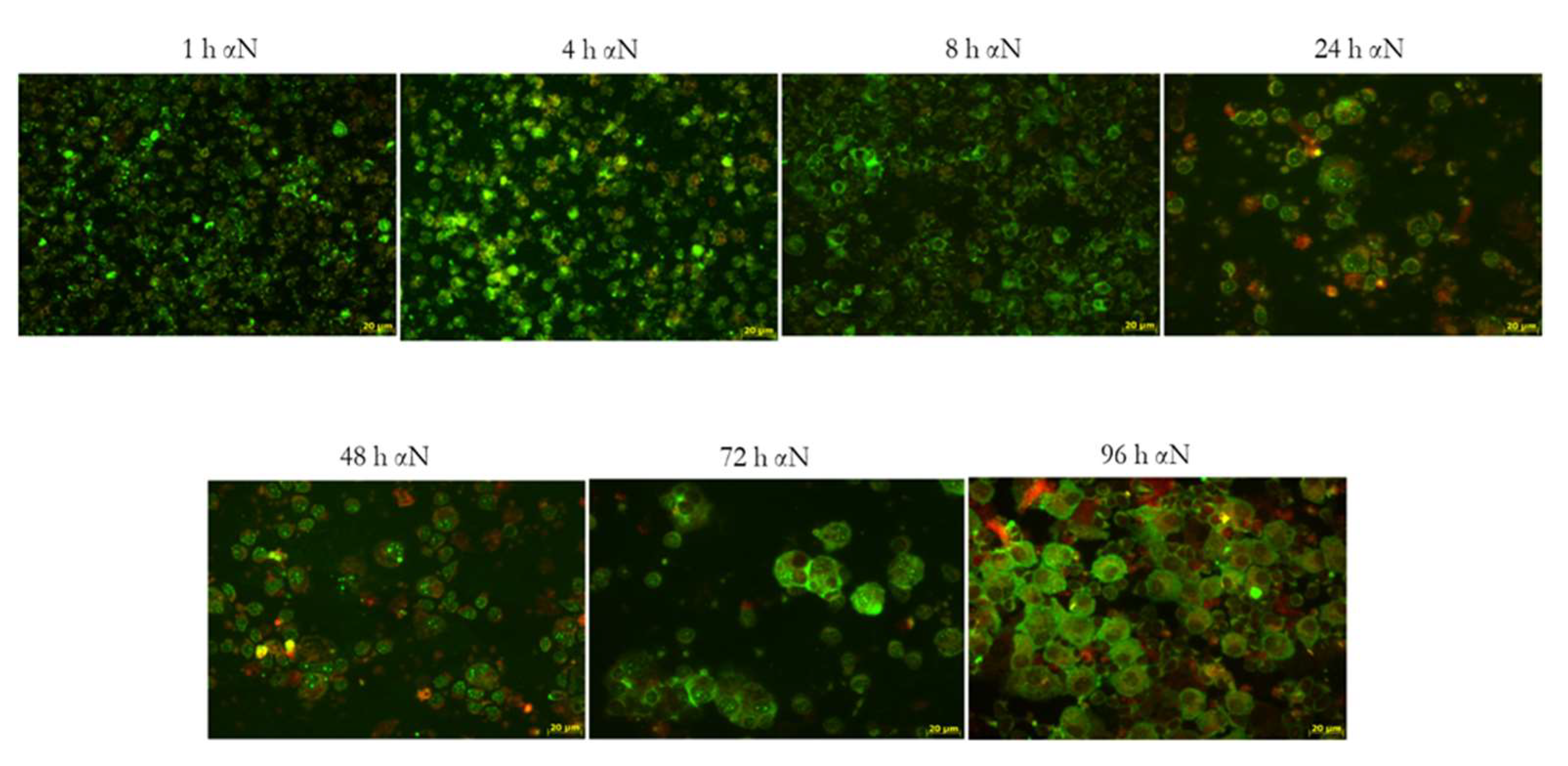
3.4. Kinetics of N Protein Uptake by MDM and MN after Co-Cultivation
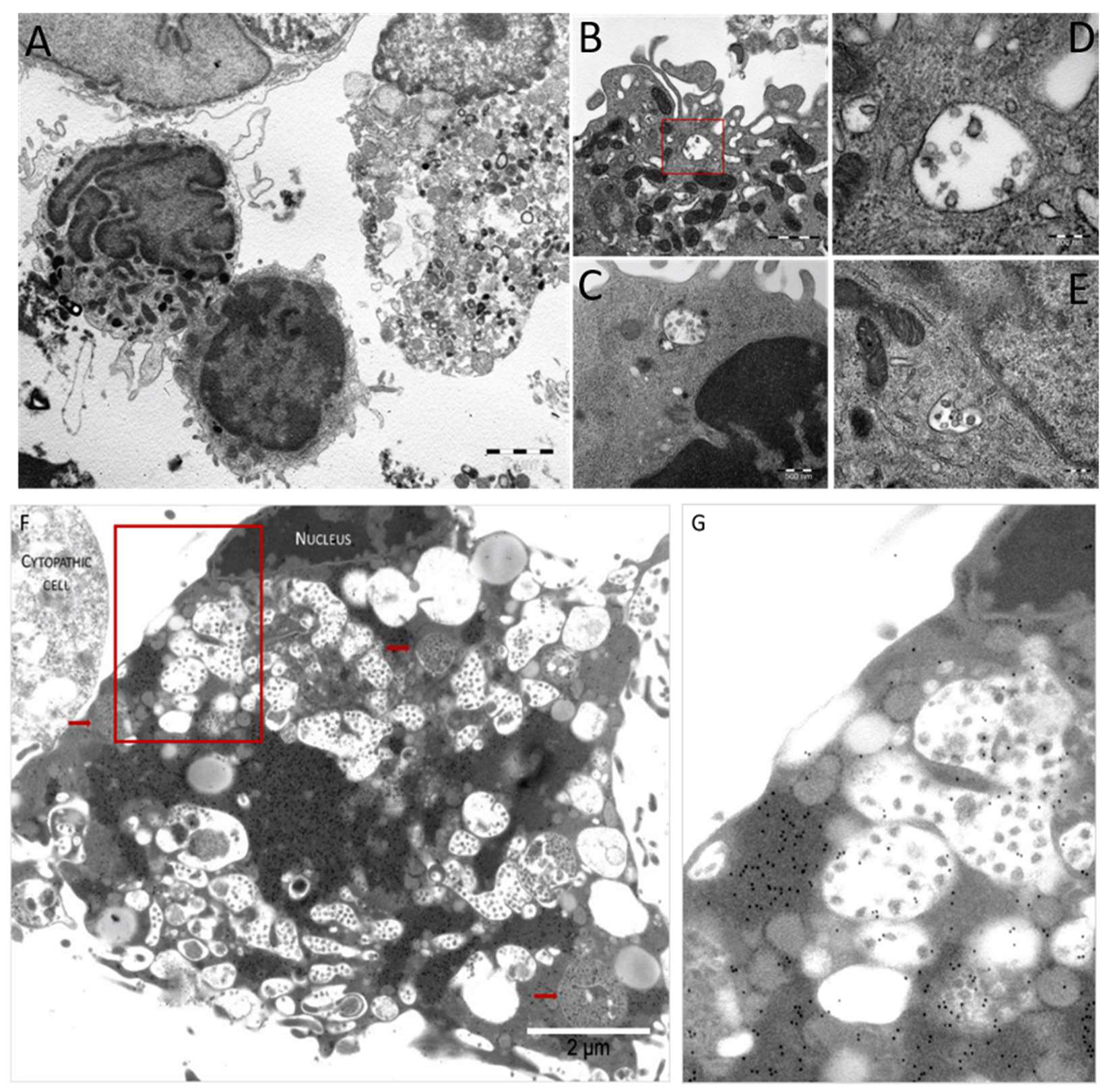
3.5. Pathologic Study of Infected MDM and VERO E6 Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cui, J.; Li, F.; Shi, Z.L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19—11 March 2020. 2020. Available online: https://www.who.int/dg/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---11-march-2020 (accessed on 17 March 2020).
- Mokhtari, T.; Hassani, F.; Ghaffari, N.; Ebrahimi, B.; Yarahmadi, A.; Hassanzadeh, G. COVID-19 and multiorgan failure: A narrative review on potential mechanisms. J. Mol. Histol. 2020, 51, 613–628. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Keidar, S.; Gamliel-Lazarovich, A.; Kaplan, M.; Pavlotzky, E.; Hamoud, S.; Hayek, T.; Karry, R.; Abassi, Z. Mineralocorticoid receptor blocker increases angiotensin-converting enzyme 2 activity in congestive heart failure patients. Circ. Res. 2005, 97, 946–953. [Google Scholar] [CrossRef] [Green Version]
- Meltzer, M.S.; Gendelman, H.E. Mononuclear phagocytes as targets, tissue reservoirs, and immunoregulatory cells in human immunodeficiency virus disease. Curr. Top. Microbiol. Immunol. 1992, 181, 239–263. [Google Scholar] [PubMed]
- Park, M.D. Macrophages: A Trojan horse in COVID-19? Nat. Rev. Immunol. 2020, 20, 351. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.; Feng, Z.; Diao, B.; Tu, C.; Qiao, Q.; Yang, H.; Zhang, Y.; Wang, G.; Wang, H.; Wang, C.; et al. SARS-CoV-2 Induces Lymphocytopenia by Promoting Inflammation and Decimates Secondary Lymphoid Organs. Front. Immunol. 2021, 12, 661052. [Google Scholar] [CrossRef] [PubMed]
- Yilla, M.; Harcourt, B.H.; Hickman, C.J.; McGrew, M.; Tamin, A.; Goldsmith, C.S.; Bellini, W.J.; Anderson, L.J. SARS-coronavirus replication in human peripheral monocytes/macrophages. Virus Res. 2005, 107, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chu, H.; Chan, J.F.; Yuen, K.Y. Middle East respiratory syndrome coronavirus infection: Virus-host cell interactions and implications on pathogenesis. Virol. J. 2015, 12, 218. [Google Scholar] [CrossRef] [Green Version]
- Boumaza, A.; Gay, L.; Mezouar, S.; Bestion, E.; Diallo, A.B.; Michel, M.; Desnues, B.; Raoult, D.; La Scola, B.; Halfon, P.; et al. Monocytes and Macrophages, Targets of Severe Acute Respiratory Syndrome Coronavirus 2: The Clue for Coronavirus Disease 2019 Immunoparalysis. J. Infect. Dis. 2021, 224, 395–406. [Google Scholar] [CrossRef]
- Revello, M.G.; Percivalle, E.; Arbustini, E.; Pardi, R.; Sozzani, S.; Gerna, G. In vitro generation of human cytomegalovirus pp65 antigenemia, viremia, and leukoDNAemia. J. Clin. Investig. 1998, 101, 2686–2692. [Google Scholar] [CrossRef] [Green Version]
- Percivalle, E.; Cambiè, G.; Cassaniti, I.; Nepita, E.V.; Maserati, R.; Ferrari, A.; Di Martino, R.; Isernia, P.; Mojoli, F.; Bruno, R.; et al. Prevalence of SARS-CoV-2 specific neutralising antibodies in blood donors from the Lodi Red Zone in Lombardy, Italy, as at 06 April 2020. Eurosurveillance 2020, 25, 2001031. [Google Scholar] [CrossRef] [PubMed]
- Alteri, C.; Cento, V.; Piralla, A.; Costabile, V.; Tallarita, M.; Colagrossi, L.; Renica, S.; Giardina, F.; Novazzi, F.; Gaiarsa, S.; et al. Genomic epidemiology of SARS-CoV-2 reveals multiple lineages and early spread of SARS-CoV-2 infections in Lombardy, Italy. Nat. Commun. 2021, 12, 434. [Google Scholar] [CrossRef] [PubMed]
- Loskutoff, N.M.; Huyser, C.; Singh, R.; Walker, D.L.; Thornhill, A.R.; Morris, L.; Webber, L. Use of a novel washing method combining multiple density gradients and trypsin for removing human immunodeficiency virus-1 and hepatitis C virus from semen. Fertil. Steril. 2005, 84, 1001–1010. [Google Scholar] [CrossRef]
- Jaroszeski, M.J.; Gilbert, R.; Heller, R. Detection and quantitation of cell-cell electrofusion products by flow cytometry. Anal. Biochem. 1994, 216, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro Surveill. 2020, 25, 2000045. [Google Scholar]
- De Gasparo, R.; Pedotti, M.; Simonelli, L.; Nickl, P.; Muecksch, F.; Cassaniti, I.; Percivalle, E.; Lorenzi, J.; Mazzola, F.; Magrì, D.; et al. Bispecific IgG neutralizes SARS-CoV-2 variants and prevents escape in mice. Nature 2021, 593, 424–428. [Google Scholar]
- Ismail, N.; Wang, Y.; Dakhlallah, D.; Moldovan, L.; Agarwal, K.; Batte, K.; Shah, P.; Wisler, J.; Eubank, T.D.; Tridandapani, S.; et al. Macrophage microvesicles induce macrophage differentiation and miR-223 transfer. Blood 2013, 121, 984–995. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhong, L.; Deng, J.; Peng, J.; Dan, H.; Zeng, X.; Li, T.; Chen, Q. High expression of ACE2 receptor of 2019-nCoV on the epithelial cells of oral mucosa. Int. J. Oral. Sci. 2020, 12, 8. [Google Scholar] [CrossRef]
- Yeager, C.L.; Ashmun, R.A.; Williams, R.K.; Cardellichio, C.B.; Shapiro, L.H.; Look, A.T.; Holmes, K.V. Human aminopeptidase N is a receptor for human coronavirus 229E. Nature 1992, 357, 420–422. [Google Scholar] [CrossRef] [Green Version]
- Shirato, K.; Kanou, K.; Kawase, M.; Matsuyama, S. Clinical Isolates of Human Coronavirus 229E Bypass the Endosome for Cell Entry. J. Virol. 2016, 91, e01387-16. [Google Scholar] [CrossRef] [Green Version]
- Shirato, K.; Kawase, M.; Matsuyama, S. Wild-type human coronaviruses prefer cell-surface TMPRSS2 to endosomal cathepsins for cell entry. Virology 2018, 517, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R. In vitro detection of apoptosis in monocytes/macrophages infected with human coronavirus. Clin. Diagn. Lab. Immunol. 2002, 9, 1392–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, J.; Uckeley, Z.M.; Doldan, P.; Stanifer, M.; Boulant, S.; Lozach, P.Y. TMPRSS2 expression dictates the entry route used by SARS-CoV-2 to infect host cells. EMBO J. 2021, 40, e107821. [Google Scholar] [CrossRef] [PubMed]
- Lempp, F.A.; Soriaga, L.; Montiel-Ruiz, M.; Benigni, F.; Noack, J.; Park, Y.J.; Bianchi, S.; Walls, A.C.; Bowen, J.E.; Zhou, J.; et al. Lectins enhance SARS-CoV-2 infection and influence neutralizing antibodies. Nature 2021. Epub ahead of print. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Dilution | Pretreatment | Secondary | Code | Specificity |
|---|---|---|---|---|---|
| SARS-CoV-2 Spike S1 | 1:200 | 2 cycles MW (900 W) + 3 cycles MW (720 W), pH 9.9 | DR | Sino Biological 40150-R007 | 2019-nCoV S |
| SARS-CoV-2 Nucleocapsid | 1:2500 | 2 cycles MW (900 W) +3 cycles MW (720 W), pH 9.9 | DR | Sino Biological 40143-R019 | 2019-nCoV N |
| CD68 (PGM1) | 1:100 | 15’ Trypsin | DM | Dako M0876 | Subcellular fraction of human MØ |
| CD68 (KP1) | 1:100 | 15’ Trypsin | DM | Santa Cruz SC-20060 | Subcellular fraction of human MØ |
| CD163 | 1:200 | no | DM | ThermoFisher Scientific MA5-11458 | Human MN and MØ. Circulating MN and most tissue MØ |
| MDM | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Characteristics | Virus Variant/ Lineage | 1 h | 4 h | 8 h | 24 h | 48 h | 72 h | 96 h | |||||||
| Mean | 95% CI | Mean | 95% CI | Mean | 95% CI | Mean | 95% CI | Mean | 95% CI | Mean | 95% CI | Mean | 95% CI | ||
| Infected Cells | B.1.1/Alpha | 30% | 19.1–41.5 | 72% | 55.1–88.2 | TMTC | na | TMTC | na | TMTC | na | TMTC | na | TMTC | na |
| Chinese | 33% | 26.2–40.5 | 81% | 71.6–90.7 | TMTC | na | TMTC | na | TMTC | na | TMTC | na | TMTC | na | |
| CPE | B.1.1/Alpha | - | na | - | na | - | na | ± | na | + | na | + | na | + | na |
| Chinese | - | na | - | na | ± | na | ± | na | + | na | + | na | + | na | |
| Average Cell dimension (µm2) | B.1.1/Alpha | 562.6 | 441.0–684.2 | 837.5 | 620–1056 | 1262 | 1029–1495 | 2246 | 1317–3175 | 4521 | 2640–6403 | 6009 | 3774–8544 | 11522 | 3197–26240 |
| Chinese | 189.3 | 131.8–246.7 | 628.2 | 550.3–706.2 | 906.9 | 653–1161 | 2416 | 1742–3090 | 3750 | 2279–5222 | 5004 | 2900–7107 | 8509 | 3095–13923 | |
| «blackberry-shaped» forms * | B.1.1/Alpha | 0 | na | 0 | na | 0 | na | 1 | 0.76–2.1 | 2 | 0.9–3.8 | 2 | 0.3–3 | 2 | 0.9–3.8 |
| Chinese | 0 | na | 0 | na | 2 | 0.9–3.8 | 5 | 0.03–9 | 2 | 0.9–3.8 | 2 | 0.4–4.5 | 2 | 0.9–3.8 | |
| Multinucleated Cells | B.1.1/Alpha | <10% | na | <10% | na | <10% | na | 32% | 26.5–36.8 | 48% | 34.2–62.5 | 51% | 33.2–68.1 | 51% | 33.2–68.1 |
| Chinese | <10% | na | 12% | 8.8–14.5 | 33% | 22.2–43.8 | 29% | 19.7–38.1 | 54% | 14.4–92.0 | 50% | 36.0–63.4 | 69% | 50.0–87.3 | |
| Spike protein | B.1.1/Alpha | - | na | - | na | - | na | - | na | - | na | - | na | + | na |
| Chinese | - | na | - | na | - | na | - | na | - | na | - | na | + | na | |
| MN | |||||||||||||||
| Infected Cells | B.1.1/Alpha/Chinese | 11% | 4.299–17.06 | 75% | 64.39–86.55 | TMTC | na | TMTC | na | TMTC | na | TMTC | na | TMTC | na |
| CPE | B.1.1/Alpha/Chinese | - | na | - | na | - | na | ± | na | + | na | + | na | + | na |
| Average Cell dimension (µm2) | B.1.1/Alpha/Chinese | 31.92 | 25.01–38.83 | 50.98 | 45.33–56.63 | 57.9 | 48.74–67.06 | 117.7 | 72.28–163.2 | 133.4 | 110.0–156.8 | 259 | 186.9–331.1 | 215.4 | 148.5–282.3 |
| «blackberry-shaped» forms * | B.1.1/Alpha/Chinese | 0 | na | 0 | na | 0 | na | 0 | na | 0 | na | 0 | na | 0 | na |
| Multinucleated Cells | B.1.1/Alpha/Chinese | <10% | na | <10% | na | <10% | na | <10% | na | 0 | na | 4.00% | 1.221–6.001 | 11% | 4.263–16.88 |
| Spike protein | B.1.1/Alpha/Chinese | - | na | - | na | - | na | - | na | - | na | - | na | + | na |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Percivalle, E.; Sammartino, J.C.; Cassaniti, I.; Arbustini, E.; Urtis, M.; Smirnova, A.; Concardi, M.; Belgiovine, C.; Ferrari, A.; Lilleri, D.; et al. Macrophages and Monocytes: “Trojan Horses” in COVID-19. Viruses 2021, 13, 2178. https://doi.org/10.3390/v13112178
Percivalle E, Sammartino JC, Cassaniti I, Arbustini E, Urtis M, Smirnova A, Concardi M, Belgiovine C, Ferrari A, Lilleri D, et al. Macrophages and Monocytes: “Trojan Horses” in COVID-19. Viruses. 2021; 13(11):2178. https://doi.org/10.3390/v13112178
Chicago/Turabian StylePercivalle, Elena, Josè Camilla Sammartino, Irene Cassaniti, Eloisa Arbustini, Mario Urtis, Alexandra Smirnova, Monica Concardi, Cristina Belgiovine, Alessandro Ferrari, Daniele Lilleri, and et al. 2021. "Macrophages and Monocytes: “Trojan Horses” in COVID-19" Viruses 13, no. 11: 2178. https://doi.org/10.3390/v13112178
APA StylePercivalle, E., Sammartino, J. C., Cassaniti, I., Arbustini, E., Urtis, M., Smirnova, A., Concardi, M., Belgiovine, C., Ferrari, A., Lilleri, D., Piralla, A., & Baldanti, F. (2021). Macrophages and Monocytes: “Trojan Horses” in COVID-19. Viruses, 13(11), 2178. https://doi.org/10.3390/v13112178